
Anti-Biofilm Activity of Phenyllactic Acid against Clinical Isolates of Fluconazole-Resistant Candida albicans

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection
2.3. Identification of Candida spp.
2.4. Antifungal Susceptibility Testing
2.5. Time-Kill Kinetics Assay of the PLA
2.6. Biofilm Formation Studies
2.7. Biofilm Treatment
2.8. Confocal Laser Scanner Microscope Observations of a Selected Biofilm
2.9. Quantitative Real-Time PCR (qRT-PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Acronym | Primer Name | Sequence (5′→3′) | Tm (°C) | Amplicon Length (bp) | Reference |
|---|---|---|---|---|---|---|
| Actin | actin | C.albicans_actin_F | AGCCCAATCCAAAAGAGGTATT | 62 | 153 | [30] |
| C.albicans_actin_R | GCTTCGGTCAACAAAACTGG | 63 | ||||
| Ergosterol biosynthesis enzyme | ERG11 | C.albicans_ERG11_F | ATTGTTGAAACTGTCATTG | 62 | 185 | [31] |
| C.albicans_ERG11_R | CCCCTAATAATATACTGATCTG | 63 | ||||
| Agglutinin like-sequence 3 | ALS3 | C.albicans_ALS3_F | CTAATGCTGCTACGTATAATT | 56 | 201 | [30] |
| C.albicans_ALS3_R | CCTGAAATTGACATGTAGCA | 58 | ||||
| Hyphal wall protein 1 | HWP1 | C.albicans_HWP1_F | CAGCCACTGAAACACCAACT | 63 | 135 | [30] |
| C.albicans_HWP1_R | CAGAAGTAACAACAACAACACCAG | 63 |
2.10. Galleria Mellonella Survival Assay
2.11. Statistical Analysis
3. Results
3.1. Microorganisms: Selection and Identification
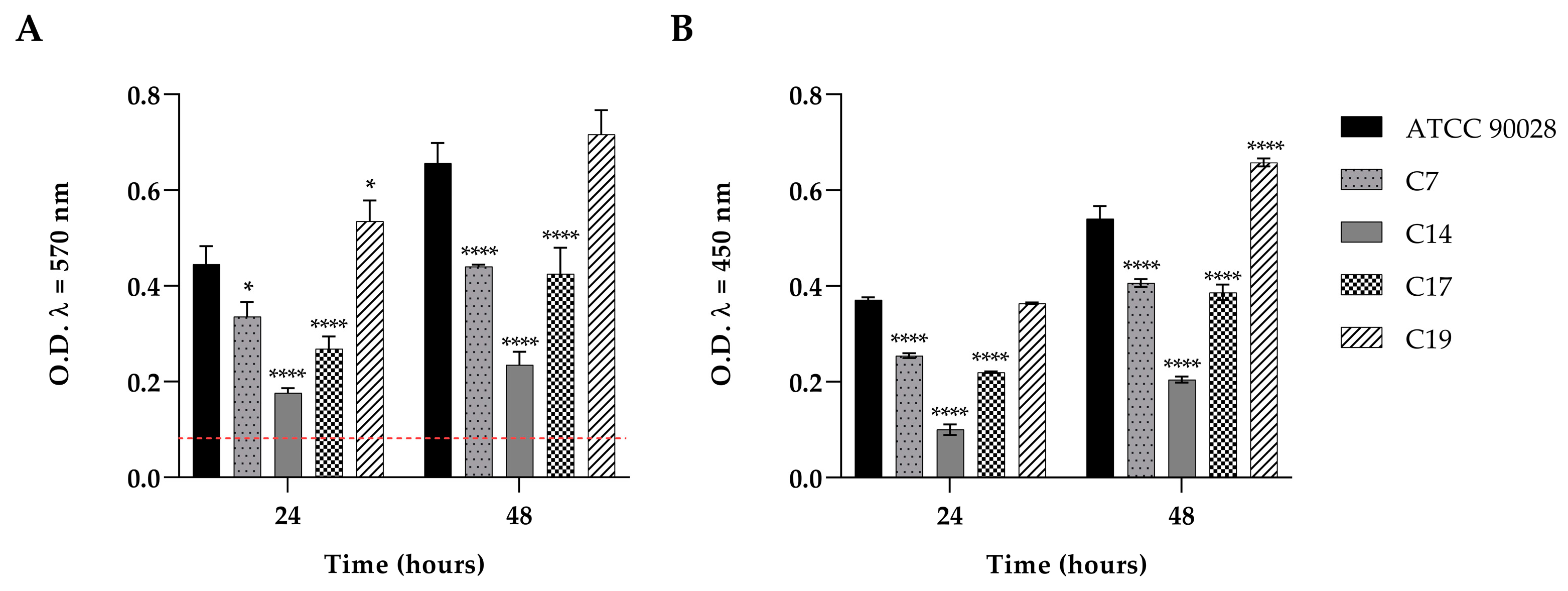
3.2. Biofilm Formation
3.3. Susceptibility of C. albicans Planktonic Cells to PLA
3.4. Effect of PLA in Biofilm Inhibition and Eradication of C. albicans
3.5. Biofilm Visualization by CLSM
3.6. Effect of PLA on ERG11, ALS3, and HWP1 Gene Expression
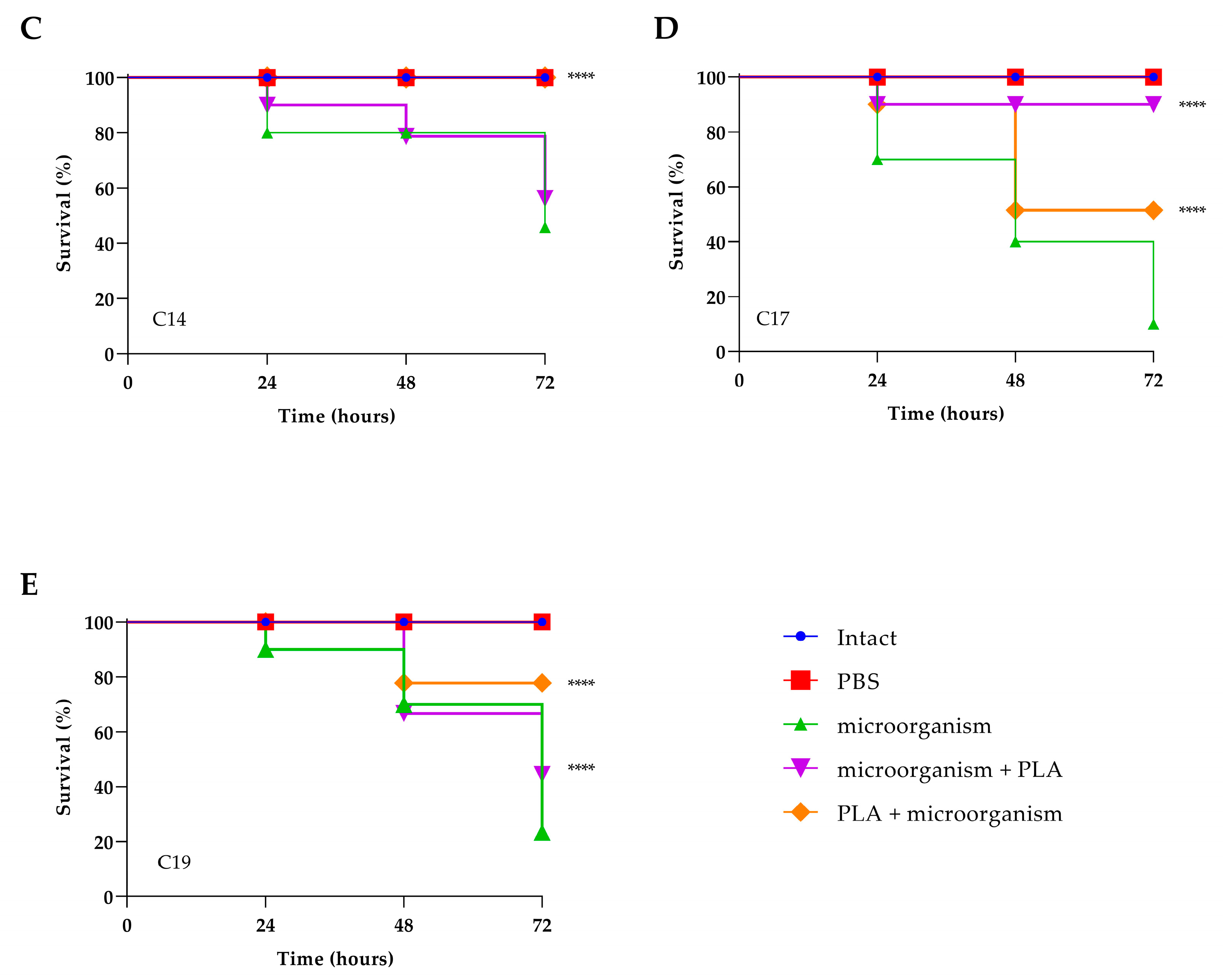
3.7. Toxicity Assessment and Evaluation of PLA Treatment of Experimental Infection by C. albicans in Galleria mellonella
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Willems, H.M.E.; Ahmed, S.S.; Liu, J.; Xu, Z.; Peters, B.M. Vulvovaginal Candidiasis: A Current Understanding and Burning Questions. J. Fungi 2020, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.; Sampaio, S.A.; De Filippis, I.; Fabri, A.; Araujo, E.S.; de Sequeira, P.C.; de Mendonça, M.C.; de Oliveira, L. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: A case study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Zhang, Y.; Zhang, T.; Xue, Y.; Xu, H.; An, R. Risk factors of vulvovaginal candidiasis among women of reproductive age in Xi’an: A cross-sectional study. BioMed Res. Int. 2018, 2018, 9703754. [Google Scholar] [CrossRef] [Green Version]
- Erfaninejad, M.; Salahshouri, A.; Amirrajab, N. Barriers and facilitators of adherence to treatment among women with vulvovaginal candidiasis: A qualitative study. Eur. J. Med. Res. 2022, 27, 303. [Google Scholar] [CrossRef]
- Anh, D.N.; Hung, D.N.; Tien, T.V.; Dinh, V.N.; Son, V.T.; Luong, N.V.; Van, N.T.; Quynh, N.T.N.; Van Tuan, N.; Tuan, L.Q.; et al. Prevalence, species distribution and antifungal susceptibility of Candida albicans causing vaginal discharge among symptomatic non-pregnant women of reproductive age at a tertiary care hospital, Vietnam. BMC Infect. Dis. 2021, 21, 523. [Google Scholar] [CrossRef]
- Salazar, S.B.; Simões, R.S.; Pedro, N.A.; Pinheiro, M.J.; Carvalho, M.F.N.N.; Mira, N.P. An Overview on Conventional and Non-Conventional Therapeutic Approaches for the Treatment of Candidiasis and Underlying Resistance Mechanisms in Clinical Strains. J. Fungi 2020, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Gabaldón, T.; Garcia-Rubio, R.; Jenks, J.D.; Hoenigl, M.; Salzer, H.J.F.; Ilkit, M.; Lass-Flörl, C.; Perlin, D.S. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Anantha, R.V.; Shaler, C.R.; Meilleur, C.E.; Parfitt, J.; Haeryfar, S.M.M.; Hernandez-Alejandro, R. The Future Liver Remnant in Patients Undergoing the Associating Liver Partition with Portal Vein Ligation for Staged Hepatectomy (ALPPS) Maintains the Immunological Components of a Healthy Organ. Front. Med. 2016, 3, 32. [Google Scholar] [CrossRef] [Green Version]
- Kadyan, S.; Rashmi, H.M.; Pradhan, D.; Kumari, A.; Chaudhari, A.; Deshwal, G.K. Effect of lactic acid bacteria and yeast fermentation on antimicrobial, antioxidative and metabolomic profile of naturally carbonated probiotic whey drink. LWT 2021, 142, 111059. [Google Scholar] [CrossRef]
- Vinicius De Melo Pereira, G.; De Carvalho Neto, D.P.; Junqueira, A.C.D.O.; Karp, S.G.; Letti, L.A.J.; Magalhães Júnior, A.I.; Soccol, C.R. A Review of Selection Criteria for Starter Culture Development in the Food Fermentation Industry. Food Rev. Int. 2020, 36, 135–167. [Google Scholar] [CrossRef]
- Sun, J.; Li, L.; Li, J. Effects of furan-phosphamide derivative on flame retardancy and crystallization behaviors of poly(lactic acid). Chem. Eng. J. 2019, 369, 150–160. [Google Scholar] [CrossRef]
- Sorrentino, E.; Tremonte, P.; Succi, M.; Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Sturchio, M.; Coppola, R. Detection of Antilisterial Activity of 3-Phenyllactic Acid Using Listeria innocua as a Model. Front. Microbiol. 2018, 9, 1373. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.; Truong, V.-L.; Lee, M.; Kim, D.; Kim, M.S.; Cho, H.; Jung, Y.H.; Yang, J.; Jeong, W.S.; Kim, Y. Investigation of phenyllactic acid as a potent tyrosinase inhibitor produced by probiotics. Curr. Res. Food Sci. 2023, 6, 100413. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Ying, J.-P.; Xin, W.-G.; Yang, L.-Y.; Li, X.-Z.; Zhang, Q.-L. Antibacterial activity and action target of phenyllactic acid against Staphylococcus aureus and its application in skim milk and cheese. J. Dairy Sci. 2022, 105, 9463–9475. [Google Scholar] [CrossRef]
- Liu, F.; Wang, F.; Du, L.; Zhao, T.; Doyle, M.P.; Wang, D.; Zhang, X.; Sun, Z.; Xu, W. Antibacterial and antibiofilm activity of phenyllactic acid against Enterobacter cloacae. Food Control 2018, 84, 442–448. [Google Scholar] [CrossRef]
- Chatterjee, M.; D’Morris, S.; Paul, V.; Warrier, S.; Vasudevan, A.K.; Vanuopadath, M.; Nair, S.S.; Paul-Prasanth, B.; Mohan, C.G.; Biswas, R. Mechanistic understanding of Phenyllactic acid mediated inhibition of quorum sensing and biofilm development in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 8223–8236. [Google Scholar] [CrossRef]
- Liu, F.; Du, L.; Zhao, T.; Zhao, P.; Doyle, M.P. Effects of phenyllactic acid as sanitizing agent for inactivation of Listeria monocytogenes biofilms. Food Control 2017, 78, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Sun, Z.; Wang, F.; Liu, Y.; Zhu, Y.; Du, L.; Wang, D.; Xu, W. Inhibition of biofilm formation and exopolysaccharide synthesis of Enterococcus faecalis by phenyllactic acid. Food Microbiol. 2020, 86, 103344. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38-Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Zare-Bidaki, M.; Maleki, A.; Ghanbarzadeh, N.; Nikoomanesh, F. Expression pattern of drug-resistance genes ERG11 and TAC1 in Candida albicans Clinical isolates. Mol. Biol. Rep. 2022, 49, 11625–11633. [Google Scholar] [CrossRef]
- CLSI, C.; Wayne, P. Method for antifungal disk diffusion susceptibility testing of yeasts, approved guideline. CLSI Doc. M44-A 2004, 24, 1–36. [Google Scholar]
- Santos, E.R.; Dal Forno, C.F.; Hernandez, M.G.; Kubiça, T.F.; Venturini, T.P.; Chassot, F.; Santurio, J.M.; Alves, S.H. Susceptibility of Candida spp. isolated from blood cultures as evaluated using the M27-A3 and new M27-S4 approved breakpoints. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, L.R.; Rosalen, P.L.; Ferreira, G.L.; Freires, I.A.; de Carvalho, F.G.; Castellano, L.R.; de Castro, R.D. Antifungal activity, mode of action and anti-biofilm effects of Laurus nobilis Linnaeus essential oil against Candida spp. Arch. Oral Biol. 2017, 73, 179–185. [Google Scholar] [CrossRef]
- Maione, A.; La Pietra, A.; de Alteriis, E.; Mileo, A.; De Falco, M.; Guida, M.; Galdiero, E. Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida auris and Klebsiella pneumoniae. Microorganisms 2022, 10, 1773. [Google Scholar] [CrossRef]
- Flamm, R.K.; Farrell, D.J.; Rhomberg, P.R.; Scangarella-Oman, N.E.; Sader, H.S. Gepotidacin (GSK2140944) In Vitro Activity against Gram-Positive and Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2017, 61, e00468-17. [Google Scholar] [CrossRef] [Green Version]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Maione, A.; Bellavita, R.; de Alteriis, E.; Galdiero, S.; Albarano, L.; La Pietra, A.; Guida, M.; Parrilli, E.; D’Angelo, C.; Galdiero, E.; et al. WMR Peptide as Antifungal and Antibiofilm against Albicans and Non-Albicans Candida Species: Shreds of Evidence on the Mechanism of Action. Int. J. Mol. Sci. 2022, 23, 2151. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Maione, A.; de Alteriis, E.; Carraturo, F.; Galdiero, S.; Falanga, A.; Guida, M.; Di Cosmo, A.; Maselli, V.; Galdiero, E. The Membranotropic Peptide gH625 to Combat Mixed Candida albicans/Klebsiella pneumoniae Biofilm: Correlation between In Vitro Anti-Biofilm Activity and In Vivo Antimicrobial Protection. J. Fungi 2021, 7, 26. [Google Scholar] [CrossRef]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Flavones, Flavonols, and Glycosylated Derivatives—Impact on Candida albicans Growth and Virulence, Expression of CDR1 and ERG11, Cytotoxicity. Pharmaceuticals 2021, 14, 27. [Google Scholar] [CrossRef]
- Bellavita, R.; Maione, A.; Merlino, F.; Siciliano, A.; Dardano, P.; De Stefano, L.; Galdiero, S.; Galdiero, E.; Grieco, P.; Falanga, A. Antifungal and Antibiofilm Activity of Cyclic Temporin L Peptide Analogues against Albicans and Non-Albicans Candida Species. Pharmaceutics 2022, 14, 454. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef] [Green Version]
- Baron, R.; Taye, M.; der Vaart, I.B.; Ujčič-Voortman, J.; Szajewska, H.; Seidell, J.C.; Verhoeff, A. The relationship of prenatal antibiotic exposure and infant antibiotic administration with childhood allergies: A systematic review. BMC Pediatr. 2020, 20, 312. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Tu, J.; Ji, C.; Li, Z.; Han, G.; Liu, N.; Li, J.; Sheng, C. Discovery of Piperidol Derivatives for Combinational Treatment of Azole-Resistant Candidiasis. ACS Infect. Dis. 2021, 7, 650–660. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prema, P.; Smila, D.; Palavesam, A.; Immanuel, G. Production and Characterization of an Antifungal Compound (3-Phenyllactic Acid) Produced by Lactobacillus plantarum Strain. Food Bioprocess Technol. 2010, 3, 379–386. [Google Scholar] [CrossRef]
- Jiang, D.; Ning, F.; Wang, Y. Additive manufacturing of biodegradable iron-based particle reinforced polylactic acid composite scaffolds for tissue engineering. J. Mater. Process. Technol. 2021, 289, 116952. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Q.; Zhao, J.; Zhang, H.; Chen, W. Streptococcus mutans and Candida albicans Biofilm Inhibitors Produced by Lactiplantibacillus plantarum CCFM8724. Curr. Microbiol. 2022, 79, 143. [Google Scholar] [CrossRef]
- Shi, C.; Liu, J.; Li, W.; Zhao, Y.; Meng, L.; Xiang, M. Expression of fluconazole resistance-associated genes in biofilm from 23 clinical isolates of Candida albicans. Braz. J. Microbiol. 2019, 50, 157–163. [Google Scholar] [CrossRef]
- Min, K.; Ichikawa, Y.; Woolford, C.A.; Mitchell, A.P. Candida albicans Gene Deletion with a Transient CRISPR-Cas9 System. mSphere 2016, 1, e00130-16. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm formation is a risk factor for mortality in patients with Candida albicans bloodstream infection-Scotland, 2012-2013. Clin. Microbiol. Infect. 2016, 22, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Prasath, K.G.; Tharani, H.; Kumar, M.S.; Pandian, S.K. Palmitic Acid Inhibits the Virulence Factors of Candida tropicalis: Biofilms, Cell Surface Hydrophobicity, Ergosterol Biosynthesis, and Enzymatic Activity. Front. Microbiol. 2020, 11, 864. [Google Scholar] [CrossRef]
- Piatek, M.; Sheehan, G.; Kavanagh, K. Galleria mellonella: The Versatile Host for Drug Discovery, In Vivo Toxicity Testing and Characterising Host-Pathogen Interactions. Antibiotics 2021, 10, 1545. [Google Scholar] [CrossRef]









| Simple | Microorganism | Accession n. |
|---|---|---|
| C1 | C. tropicalis 16 | EF216862.1 |
| C2 | C. albicans SKH402 | MT640023.1 |
| C3 | C. albicans SKH402 | MT640023.1 |
| C4 | C. glabrata AO34 | KX273821.1 |
| C5 | C. albicans SKH402 | MT640023.1 |
| C6 | C. albicans SKH402 | MT640023.1 |
| C7 | C. albicans AUMC13495 | MH534919.1 |
| C8 | C. haemulonii LIP Ch9 | KJ476201 |
| C9 | C. albicans SKH402 | MT640023.1 |
| C10 | C. dubliniensis 94234 | KF537273 |
| C11 | C. albicans AUMC13532 | MH534924.1 |
| C12 | C. albicans JRP61 | MF276778.1 |
| C13 | C. albicans N96C | KP675681.1 |
| C14 | C. albicans n43b | KP675647.1 |
| C15 | C. glabrata RCPF 1398 | MK998697.1 |
| C16 | C. dubliniensis 95677 | KF537274 |
| C17 | C. albicans n99b | KP675684.1 |
| C18 | C. albicans P51L1 | KP878253.1 |
| C19 | C. albicans 123D | KP765027.1 |
| C20 | C. albicans AUMC13532 | MH534924.1 |
| C21 | C. albicans JRP61 | MF276778.1 |
| C22 | C. dubliniensis 98277 | KF537275 |
| C23 | C. albicans SKH402 | MT640023.1 |
| C24 | C. albicans m62a | KP675249.1 |
| C25 | C. orthopsilosis 1719 | JQ023169 |
| C26 | C. albicans FC6841 | MH628212.1 |
| C27 | C. albicans SKH402 | MT640023.1 |
| C28 | C. albicans N96C | KP675681.1 |
| C29 | C. haemulonii LIP Ch4 | KJ476196 |
| C30 | C. albicans P51L1 | KP878253.1 |
| C31 | C. albicans FC6841 | MH628212.1 |
| Semple | AMB | FLC | CSF |
|---|---|---|---|
| C2 C. albicans | S | S | S |
| C3 C. albicans | S | S | S |
| C5 C. albicans | S | S | S |
| C6 C. albicans | S | S | S |
| C7 C. albicans | S | R | S |
| C9 C. albicans | S | S | S |
| C11 C. albicans | S | S | S |
| C12 C. albicans | S | S | S |
| C13 C. albicans | S | S | S |
| C14 C. albicans | S | R | I |
| C17 C. albicans | S | R | S |
| C18 C. albicans | S | S | S |
| C19 C. albicans | S | R | R |
| C20 C. albicans | S | S | S |
| C21 C. albicans | S | S | S |
| C23 C. albicans | S | S | S |
| C24 C. albicans | S | I | S |
| C26 C. albicans | S | S | S |
| C27 C. albicans | S | S | S |
| C28 C. albicans | S | S | S |
| C30 C. albicans | S | S | S |
| C31 C. albicans | S | S | S |
| PLA (mg mL−1) | ||||
|---|---|---|---|---|
| MIC | MFC | MFC/MIC Ratio | ||
| ATCC90028 | 7.5 | 7.5 | 1.0 | fungicidal |
| C7 | 7.5 | 7.5 | 1.0 | fungicidal |
| C14 | 7.5 | 7.5 | 1.0 | fungicidal |
| C17 | 7.5 | 10.0 | 1.3 | fungicidal |
| C19 | 7.5 | 10.0 | 1.3 | fungicidal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maione, A.; Imparato, M.; Buonanno, A.; Carraturo, F.; Schettino, A.; Schettino, M.T.; Galdiero, M.; de Alteriis, E.; Guida, M.; Galdiero, E. Anti-Biofilm Activity of Phenyllactic Acid against Clinical Isolates of Fluconazole-Resistant Candida albicans. J. Fungi 2023, 9, 355. https://doi.org/10.3390/jof9030355
Maione A, Imparato M, Buonanno A, Carraturo F, Schettino A, Schettino MT, Galdiero M, de Alteriis E, Guida M, Galdiero E. Anti-Biofilm Activity of Phenyllactic Acid against Clinical Isolates of Fluconazole-Resistant Candida albicans. Journal of Fungi. 2023; 9(3):355. https://doi.org/10.3390/jof9030355
Chicago/Turabian StyleMaione, Angela, Marianna Imparato, Annalisa Buonanno, Federica Carraturo, Antonetta Schettino, Maria Teresa Schettino, Marilena Galdiero, Elisabetta de Alteriis, Marco Guida, and Emilia Galdiero. 2023. "Anti-Biofilm Activity of Phenyllactic Acid against Clinical Isolates of Fluconazole-Resistant Candida albicans" Journal of Fungi 9, no. 3: 355. https://doi.org/10.3390/jof9030355
APA StyleMaione, A., Imparato, M., Buonanno, A., Carraturo, F., Schettino, A., Schettino, M. T., Galdiero, M., de Alteriis, E., Guida, M., & Galdiero, E. (2023). Anti-Biofilm Activity of Phenyllactic Acid against Clinical Isolates of Fluconazole-Resistant Candida albicans. Journal of Fungi, 9(3), 355. https://doi.org/10.3390/jof9030355