
Influence of Mycorrhiza on C:N:P Stoichiometry in Senesced Leaves

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
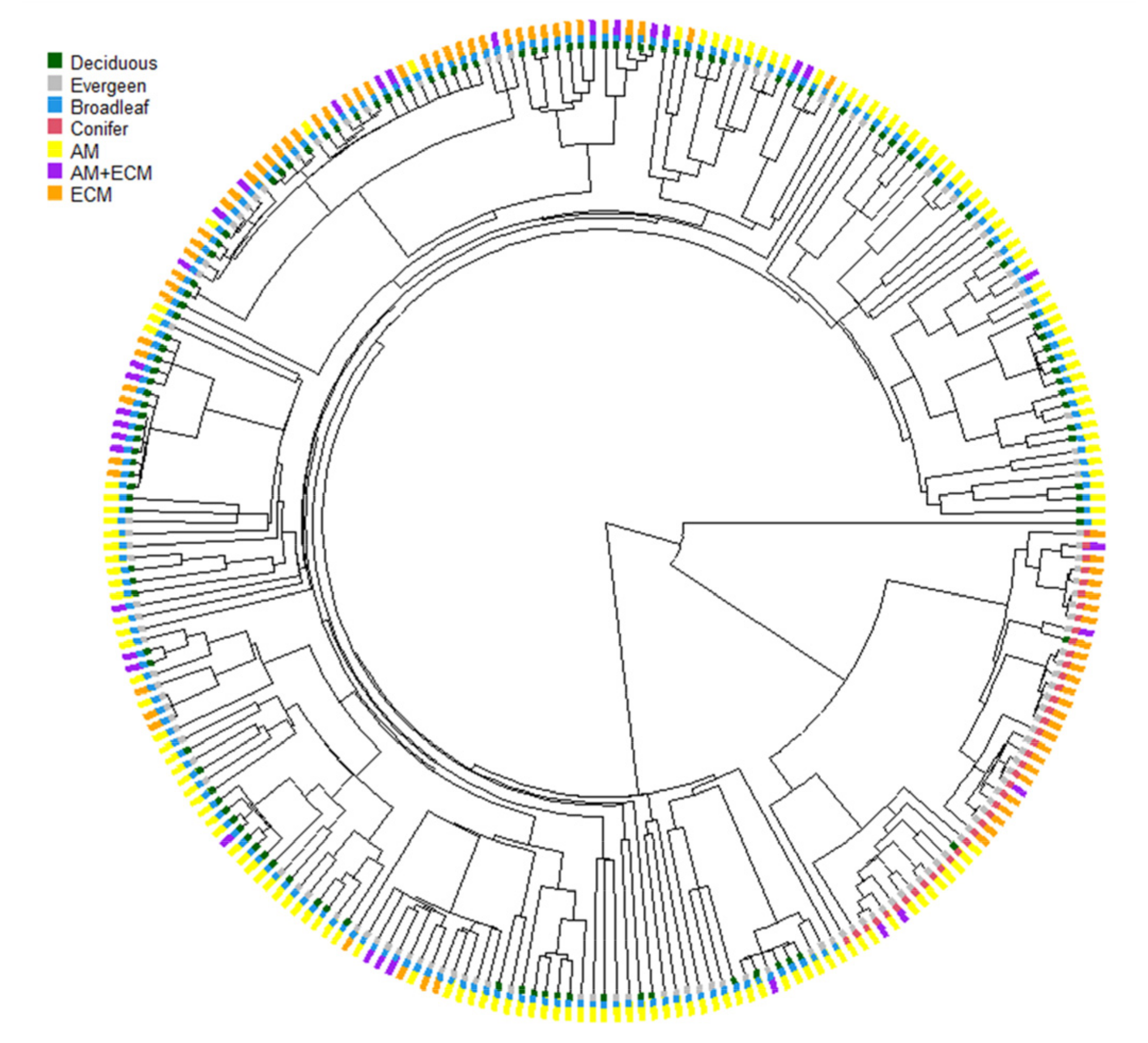
2.2. Mycorrhizal Classification
2.3. Data Analysis
3. Results
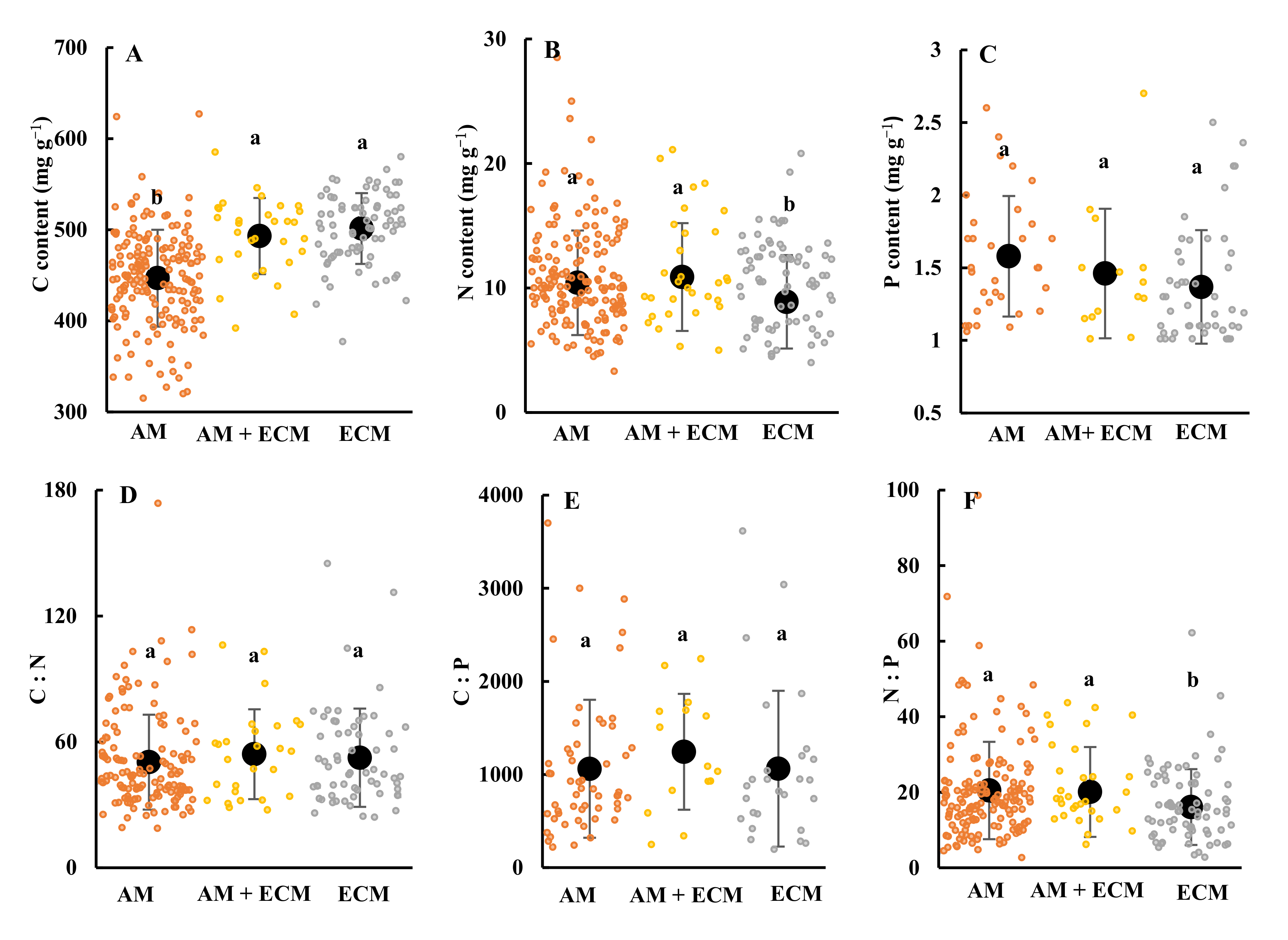
3.1. The Contents of Senesced Leaves’ Carbon, Nitrogen and Phosphorus and Their Stoichiometric Ratios among Plant Associations with Different Mycorrhizal Types
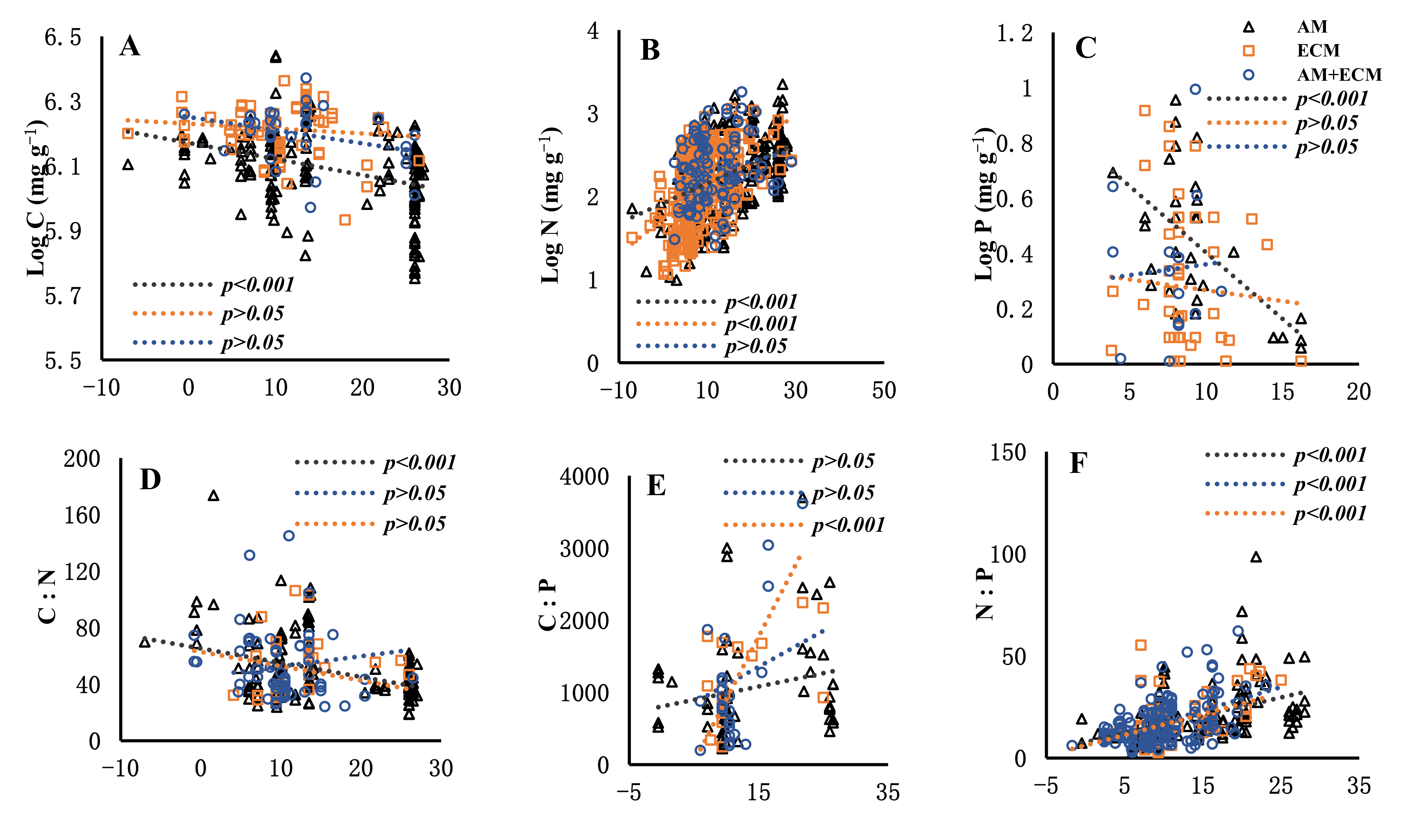
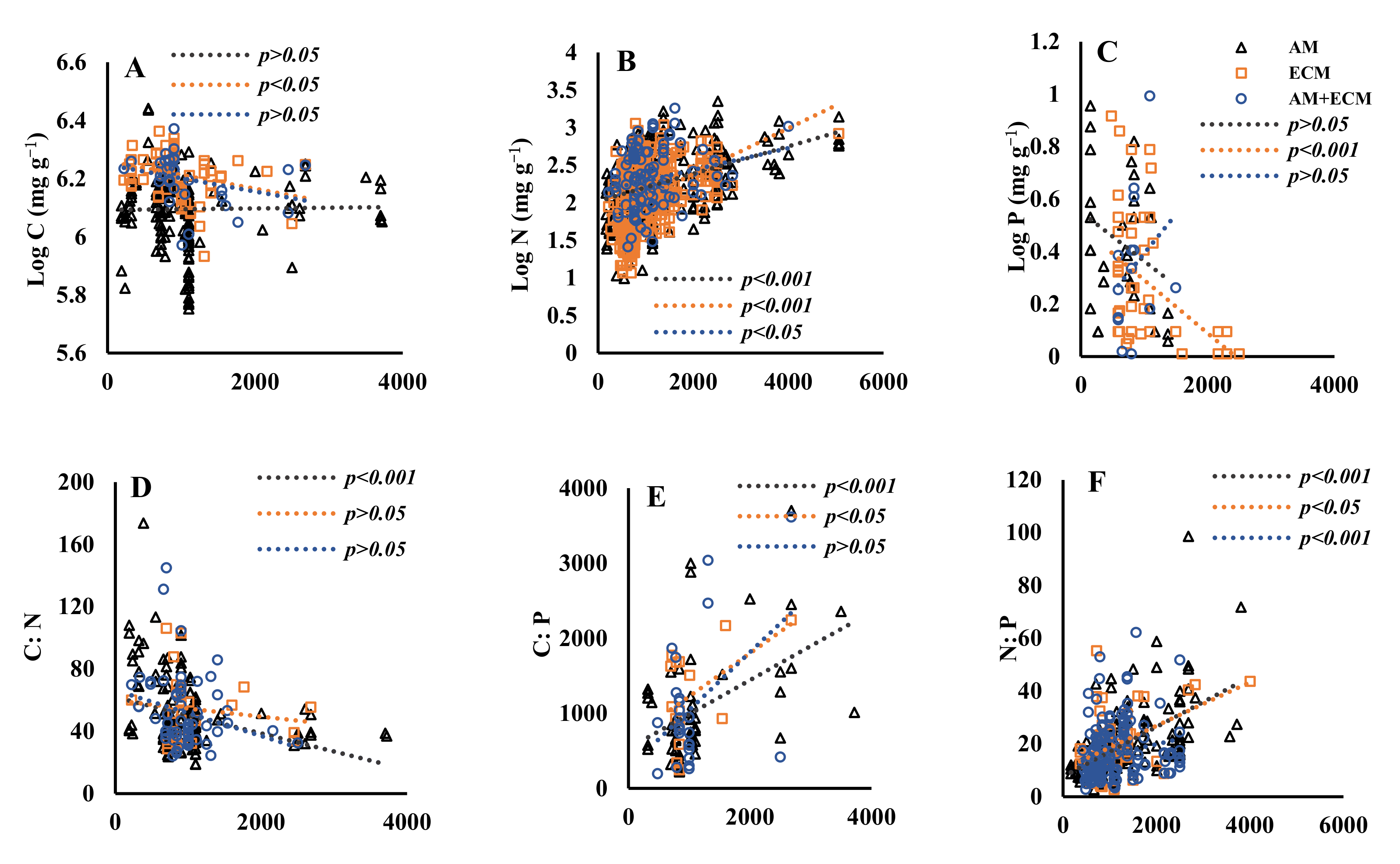
3.2. The Regulation of Senesced Leaves’ Carbon, Nitrogen, Phosphorus and Their Stoichiometric Ratios by MAT and MAP among Plant Associations with Different Mycorrhizal Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef]
- Jiang, C.; Yu, G.; Li, Y.; Cao, G.; Yang, Z.P.; Sheng, W.; Yu, W. Nutrient resorption of coexistence species in alpine meadow of the Qinghai-Tibetan Plateau explains plant adaptation to nutrient-poor environment. Ecol. Eng. 2012, 44, 1–9. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Cui, J.; Zhao, X.; Zhang, T. Nutrient resorption in Caragana microphylla along a chronosequence of plantations: Implications for desertified land restoration in North China. Ecol. Eng. 2013, 53, 299–305. [Google Scholar] [CrossRef]
- Qiu, X.; Wang, H.; Peng, D.; Liu, X.; Yang, F.; Li, Z.; Cheng, S. Thinning drives C:N:P stoichiometry and nutrient resorption in Larix principis-rupprechtii plantations in North China. For. Ecol. Manag. 2020, 462, 117984. [Google Scholar] [CrossRef]
- Givnish, T.J.; Dawson, T.E.; Killingbeck, K.T. Nutrients in Senesced Leaves: Keys to the Search for Potential Resorption and Resorption Proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Teklay, T. Seasonal dynamics in the concentrations of macronutrients and organic constituents in green and senesced leaves of three agroforestry species in southern Ethiopia. Plant Soil 2004, 267, 297–307. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S. The Mineral Nutrition of Wild Plants Revisited: A Re-evaluation of Processes and Patterns. Adv. Ecol. Res. 1999, 30, 1–67. [Google Scholar] [CrossRef]
- Eckstein, R.L.; Karlsson, P.S.; Weih, M. Leaf life span and nutrient resorption as determinants of plant nutrient conservation in temperate-arctic regions. New Phytol. 1999, 143, 177–189. [Google Scholar] [CrossRef]
- Hawkesford, M.J. Reducing the reliance on nitrogen fertilizer for wheat production. J. Cereal Sci. 2014, 59, 276–283. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on Wheat Yield and Quality with Reduced Nitrogen Supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Anderegg, L.D.L.; Berner, L.T.; Badgley, G.; Sethi, M.L.; Law, B.E.; HilleRisLambers, J. Within-species patterns challenge our understanding of the leaf economics spectrum. Ecol. Lett. 2018, 21, 734–744. [Google Scholar] [CrossRef] [PubMed]
- de la Riva, E.G.; Tosto, A.; Pérez-Ramos, I.M.; Navarro-Fernández, C.M.; Olmo, M.; Anten, N.P.R.; Marañón, T.; Villar, R. A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? J. Veg. Sci. 2016, 27, 187–199. [Google Scholar] [CrossRef]
- Hungate, B.A.; Dukes, J.S.; Shaw, M.R.; Luo, Y.; Field, C.B. Nitrogen and Climate Change. Science 2003, 302, 1512–1513. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Van Bodegom, P.M.; Aerts, R.; Callaghan, T.V.; Van Logtestijn, R.S.; Alatalo, J.; Chapin, F.S.; Gerdol, R.; Gudmundsson, J.; Gwynn-Jones, D.; et al. Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol. Lett. 2007, 10, 619–627. [Google Scholar] [CrossRef]
- Shi, Z.; Li, K.; Zhu, X.; Wang, F. The worldwide leaf economic spectrum traits are closely linked with mycorrhizal traits. Fungal Ecol. 2020, 43, 100877. [Google Scholar] [CrossRef]
- Yang, S.; Shi, Z.; Zhang, M.; Li, Y.; Gao, J.; Wang, X.; Liu, D. Stoichiometry of Carbon, Nitrogen and Phosphorus in Shrub Organs Linked Closely With Mycorrhizal Strategy in Northern China. Front. Plant Sci. 2021, 12, 687347. [Google Scholar] [CrossRef]
- Butterly, C.R.; Armstrong, R.; Chen, D.; Tang, C. Carbon and nitrogen partitioning of wheat and field pea grown with two nitrogen levels under elevated CO2. Plant Soil 2015, 391, 367–382. [Google Scholar] [CrossRef]
- Couture, J.J.; Meehan, T.D.; Lindroth, R.L. Atmospheric change alters foliar quality of host trees and performance of two outbreak insect species. Oecologia 2012, 168, 863–876. [Google Scholar] [CrossRef]
- Pleijel, H.; Högy, P. CO2 dose-response functions for wheat grain, protein and mineral yield based on FACE and open-top chamber experiments. Environ. Pollut. 2015, 198, 70–77. [Google Scholar] [CrossRef]
- Sardans, J.; Grau, O.; Chen, H.Y.H.; Janssens, I.A.; Ciais, P.; Piao, S.; Peñuelas, J. Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Chang. Biol. 2017, 23, 3849–3856. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Güsewell, S.; Koerselman, W.; Verhoeven, J.T.A. Biomass N:P Ratios as Indicators of Nutrient Limitation for Plant Population in Wetlands. Ecol. Appl. 2003, 13, 372–384. [Google Scholar] [CrossRef]
- Xia, C.; Yu, D.; Wang, Z.; Xie, D. Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China. Ecol. Eng. 2014, 70, 406–413. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nat. Clim. Chang. 2015, 5, 465–469. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Negative effects of fertilization on plant nutrient resorption. Ecology 2015, 96, 373–380. [Google Scholar] [CrossRef]
- Güsewell, S. High nitrogen: Phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytol. 2005, 166, 537–550. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, H.; Yu, Q.; Wang, Z.; Wei, C.; Long, M.; Kattge, J.; Smith, M.; Han, X. Sampling Date, Leaf Age and Root Size: Implications for the Study of Plant C:N:P Stoichiometry. PLoS ONE 2013, 8, e60360. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Deng, L.; Guo, X.; Han, F.; Ju, W.; Wang, X.; Chen, H.; Tan, W.; Zhang, X. Patterns of soil microbial nutrient limitations and their roles in the variation of soil organic carbon across a precipitation gradient in an arid and semi-arid region. Sci. Total Environ. 2019, 658, 1440–1451. [Google Scholar] [CrossRef]
- Yan, W.; Zhong, Y.; Shangguan, Z. The relationships and sensibility of wheat C:N:P stoichiometry and water use efficiency under nitrogen fertilization. Plant Soil Environ. 2015, 61, 201–207. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Q.; Yin, H.; Zhao, C.; Zhang, L.; Zhou, G.; Yin, C.; Lu, Z.; Xiong, G.; Li, Y.; et al. C:N:P stoichiometry of Ericaceae species in shrubland biomes across Southern China: Influences of climate, soil and species identity. J. Plant Ecol. 2019, 12, 346–357. [Google Scholar] [CrossRef]
- Du, C.; Wang, X.; Zhang, M.; Jing, J.; Gao, Y. Effects of elevated CO2 on plant C-N-P stoichiometry in terrestrial ecosystems: A meta-analysis. Sci. Total Environ. 2019, 650, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef]
- Mao, R.; Chen, H.M.; Zhang, X.H.; Shi, F.X.; Song, C.C. Effects of P addition on plant C:N:P stoichiometry in an N-limited temperate wetland of Northeast China. Sci. Total Environ. 2016, 559, 1–6. [Google Scholar] [CrossRef]
- Bo, F.; Zhang, Y.; Chen, H.Y.H.; Wang, P.; Ren, X.; Guo, J. The C:N:P stoichiometry of planted and natural larix principis-rupprechtii stands along altitudinal gradients on the Loess Plateau, China. Forests 2020, 11, 363. [Google Scholar] [CrossRef]
- Sun, X.; Guo, J.; Guo, S.; Guo, H.; Hu, S. Divergent responses of leaf N:P:K stoichiometry to nitrogen fertilization in rice and weeds. Weed Sci. 2019, 67, 339–345. [Google Scholar] [CrossRef]
- Averill, C.; Bhatnagar, J.M.; Dietze, M.C.; Pearse, W.D.; Kivlin, S.N. Global imprint of mycorrhizal fungi on whole-plant nutrient economics. Proc. Natl. Acad. Sci. USA 2019, 116, 23163–23168. [Google Scholar] [CrossRef] [PubMed]
- Sizonenko, T.A.; Dubrovskiy, Y.A.; Novakovskiy, A.B. Changes in mycorrhizal status and type in plant communities along altitudinal and ecological gradients—A case study from the Northern Urals (Russia). Mycorrhiza 2020, 30, 445–454. [Google Scholar] [CrossRef]
- Kyaschenko, J.; Clemmensen, K.E.; Karltun, E.; Lindahl, B.D. Below-ground organic matter accumulation along a boreal forest fertility gradient relates to guild interaction within fungal communities. Ecol. Lett. 2017, 20, 1546–1555. [Google Scholar] [CrossRef]
- Jiang, J.; Moore, J.A.M.; Priyadarshi, A.; Classen, A.T. Plant-mycorrhizal interactions mediate plant community coexistence by altering resource demand. Ecology 2017, 98, 187–197. [Google Scholar] [CrossRef]
- Rosling, A.; Midgley, M.G.; Cheeke, T.; Urbina, H.; Fransson, P.; Phillips, R.P. Phosphorus cycling in deciduous forest soil differs between stands dominated by ecto- and arbuscular mycorrhizal trees. New Phytol. 2016, 209, 1184–1195. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.B.; Sweeney, S.; Brzostek, E.R.; Evans, T.P.; Johnson, D.J.; Myers, J.A.; Bourg, N.A.; Wolf, A.T.; Howe, R.W.; Phillips, R.P. Tree-mycorrhizal associations detected remotely from canopy spectral properties. Glob. Chang. Biol. 2016, 22, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.B.; Lajtha, K.; Crow, S.E.; Hugelius, G.; Kramer, M.G.; Piñeiro, G. The Ecology of Soil Carbon: Pools, Vulnerabilities, and Biotic and Abiotic Controls. Rev. Ecol. Evol. Syst. 2017, 48, 419–445. [Google Scholar] [CrossRef]
- Cortois, R.; Schroder-Georgi, T.; Weigelt, A.; van der Putten, W.H.; De Deyn, G.B. Plant–soil feedbacks: Role of plant functional group and plant traits. J. Ecol. 2016, 104, 1608–1617. [Google Scholar] [CrossRef]
- Hoeksema, J.D.; Chaudhary, V.B.; Cehring, G.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.J.; Wang, S.Y.; Liu, Y.; Li, J.; Shen, F.Y.; Yang, L.X. Responses of mycorrhizal colonization to nitrogen and phosphorus addition in fourteen woody and herbaceous species: The roles of hypodermal passage cells and penetration points. Plant Soil 2021, 469, 273–285. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, E.B.; Egerton-Warburton, L.M.; Hart, M.M.; Klironomos, J.N.; Maherali, H.; Tedersoo, L. Arbuscular mycorrhizal fungi as mediators of ecosystem responses to nitrogen deposition: A trait-based predictive framework. J. Ecol. 2018, 106, 480–489. [Google Scholar] [CrossRef]
- Luo, S.; Schmid, B.; De Deyn, G.B.; Yu, S. Soil microbes promote complementarity effects among co-existing trees through soil nitrogen partitioning. Funct. Ecol. 2018, 32, 1879–1889. [Google Scholar] [CrossRef]
- Turner, B.L. Resource partitioning for soil phosphorus: A hypothesis. J. Ecol. 2008, 96, 698–702. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis; Elsevier Ltd.: Amsterdam, The Netherlands, 2008; ISBN 9780123705266. [Google Scholar]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Michelsen, A.; Schmidt, I.K.; Jonasson, S.; Quarmby, C.; Sleep, D. Leaf 15N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non-and arbuscular mycorrhizal species access different sources of soil nitrogen. Oecologia 1996, 105, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Thormann, M.N.; Currah, R.S.; Bayley, S.E. The mycorrhizal status of the dominant vegetation along a peatland gradient in southern boreal Alberta, Canada. Wetlands 1999, 19, 438–450. [Google Scholar] [CrossRef]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon–nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef]
- Bennett, J.A.; Maherali, H.; Reinhart, K.O.; Lekberg, Y.; Hart, M.M.; Klironomos, J. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 2017, 355, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367, eaba1223. [Google Scholar] [CrossRef]
- Chapman, S.K.; Langley, J.A.; Hart, S.C.; Koch, G.W. Plants actively control nitrogen cycling: Uncorking the microbial bottleneck. New Phytol. 2006, 169, 27–34. [Google Scholar] [CrossRef]
- Cornelissen, J.; Aerts, R.; Cerabolini, B.; Werger, M.; Van der Heijden, M. Carbon cycling traits of plant species are linked with mycorrhizal strategy. Oecologia 2001, 129, 611–619. [Google Scholar] [CrossRef]
- Lin, G.; McCormack, M.L.; Ma, C.; Guo, D. Similar below-ground carbon cycling dynamics but contrasting modes of nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests. New Phytol. 2017, 213, 1440–1451. [Google Scholar] [CrossRef]
- Kobe, R.K.; Lepczyk, C.A.; Iyer, M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 2005, 86, 2780–2792. [Google Scholar] [CrossRef]
- Keller, A.B.; Phillips, R.P. Leaf litter decay rates differ between mycorrhizal groups in temperate, but not tropical, forests. New Phytol. 2019, 222, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Wurzburger, N.; Brookshire, E.N.J. Experimental evidence that mycorrhizal nitrogen strategies affect soil carbon. Ecology 2017, 98, 1491–1497. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Global trends in senesced-leaf nitrogen and phosphorus. Glob. Ecol. Biogeogr. 2009, 18, 532–542. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Akhmetzhanova, A.A.; Soudzilovskaia, N.A.; Onipchenko, V.G.; Cornwell, W.K.; Agafonov, V.A.; Selivanov, I.A.; Cornelissen, J.H.C. A rediscovered treasure: Mycorrhizal intensity database for 3000 vascular plant species across the former Soviet Union. Ecology 2012, 93, 689–690. [Google Scholar] [CrossRef]
- Hempel, S.; Gotzenberger, L.; Kuhn, I.; Michalski, S.G.; Rillig, M.C.; Zobel, M.; Moora, M. Mycorrhizas in the Central European flora: Relationships with plant life history traits and ecology. Ecology 2013, 94, 1389–1399. [Google Scholar] [CrossRef]
- Soudzilovskaia, N.A.; Vaessen, S.; Barcelo, M.; He, J.; Rahimlou, S.; Abarenkov, K.; Brundrett, M.C.; Gomes, S.I.F.; Merckx, V.; Tedersoo, L. FungalRoot: Global online database of plant mycorrhizal associations. New Phytol. 2020, 227, 955–966. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, X.; He, Y.; Fu, Y.; Du, Z.; Lu, M.; Sun, X.; Li, C.; Lu, C.; Liu, R.; et al. Global systematic review with meta-analysis shows that warming effects on terrestrial plant biomass allocation are influenced by precipitation and mycorrhizal association. Nat. Commun. 2022, 13, 4914. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornellssen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Xu, J.-W.; Lin, G.; Liu, B.; Mao, R. Linking leaf nutrient resorption and litter decomposition to plant mycorrhizal associations in boreal peatlands. Plant Soil 2020, 448, 413–424. [Google Scholar] [CrossRef]
- Yu, H.; Fan, J.; Harris, W.; Li, Y. Relationships between below-ground biomass and foliar N:P stoichiometry along climatic and altitudinal gradients of the Chinese grassland transect. Plant Ecol. 2017, 218, 661–671. [Google Scholar] [CrossRef]
- Zhao, N.; Yu, G.; He, N.; Xia, F.; Wang, Q.; Wang, R.; Xu, Z.; Jia, Y. Invariant allometric scaling of nitrogen and phosphorus in leaves, stems, and fine roots of woody plants along an altitudinal gradient. J. Plant Res. 2016, 129, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Toussaint, A.; Bueno, G.; Davison, J.; Moora, M.; Tedersoo, L.; Zobel, M.; Öpik, M.; Pärtel, M. Asymmetric patterns of global diversity among plants and mycorrhizal fungi. J. Veg. Sci. 2020, 31, 355–366. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Xiao-Tao, L.; Hartmann, H.; Keller, A.; Han, X.G.; Trumbore, S.; Phillips, R.P. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales. Glob. Ecol. Biogeogr. 2018, 27, 875–885. [Google Scholar] [CrossRef]
- Averill, C.; Hawkes, C.V. Ectomycorrhizal fungi slow soil carbon cycling. Ecol Lett. 2016, 19, 937–947. [Google Scholar] [CrossRef]
- Orwin, K.H.; Kirschbaum, M.U.F.; St John, M.G.; Dickie, I.A. Organic nutrient uptake by mycorrhizal fungi enhances ecosystem carbon storage: A model-based assessment. Ecol. Lett. 2011, 14, 493–502. [Google Scholar] [CrossRef]
- Wang, W.Q.; Sardans, J.; Wang, C.; Zeng, C.S.; Tong, C.; Asensio, D.; Peñuelas, J. Ecological stoichiometry of C, N, and P of invasive Phragmites australis and native Cyperus malaccensis species in the Minjiang River tidal estuarine wetlands of China. Plant Ecol. 2015, 216, 809–822. [Google Scholar] [CrossRef]
- Zand, E.; Soufizadeh, S.; Eskandari, A. Water stress and nitrogen limitation effects on corn (Zea mays L.) competition with a C3 and a C4 weed. Commun. Agric. Appl. Biol. Sci. 2006, 71, 753–760. [Google Scholar]
- Kocacinar, F.; Sage, R.F. Photosynthetic pathway alters xylem structure and hydraulic function in herbaceous plants. Plant. Cell Environ. 2003, 26, 2015–2026. [Google Scholar] [CrossRef]
- Dawes, M.A.; Hagedorn, F.; Handa, I.T.; Streit, K.; Ekblad, A.; Rixen, C.; Körner, C.; Hättenschwiler, S. An alpine treeline in a carbon dioxide-rich world: Synthesis of a nine-year free-air carbon dioxide enrichment study. Oecologia 2013, 171, 623–637. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; Van Der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Han, W.; Peñuelas, J.; Sardans, J.; Elser, J.J.; Du, E.; Reich, P.B.; Fang, J. Phosphorus accumulates faster than nitrogen globally in freshwater ecosystems under anthropogenic impacts. Ecol. Lett. 2016, 19, 1237–1246. [Google Scholar] [CrossRef]
- Shi, M.; Fisher, J.B.; Brzostek, E.R.; Phillips, R.P. Carbon cost of plant nitrogen acquisition: Global carbon cycle impact from an improved plant nitrogen cycle in the Community Land Model. Glob. Chang. Biol. 2016, 22, 1299–1314. [Google Scholar] [CrossRef] [PubMed]
- Terrer, C.; Vicca, S.; Stocker, B.D.; Hungate, B.A.; Phillips, R.P.; Reich, P.B.; Finzi, A.C.; Prentice, I.C. Ecosystem responses to elevated CO2 governed by plant-soil interactions and the cost of nitrogen acquisition. New Phytol. 2018, 217, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R. Nutrient Resorption from Senescing Leaves of Perennials: Are there General Patterns? J. Ecol. 1996, 84, 597. [Google Scholar] [CrossRef]
- Chuyong, G.B.; Newbery, D.M.; Songwe, N.C. Litter nutrients and retranslocation in a central African rain forest dominated by ectomycorrhizal trees. New Phytol. 2000, 148, 493–510. [Google Scholar] [CrossRef]
- Bothwell, L.D.; Selmants, P.C.; Giardina, C.P.; Litton, C.M. Leaf litter decomposition rates increase with rising mean annual temperature in Hawaiian tropical montane wet forests. PeerJ 2014, 2. [Google Scholar] [CrossRef]
- Chen, W.; Koide, R.T.; Adams, T.S.; DeForest, J.L.; Cheng, L.; Eissenstat, D.M. Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees. Proc. Natl. Acad. Sci. USA 2016, 113, 8741–8746. [Google Scholar] [CrossRef]
- Carrera, A.L.; Sain, C.L.; Bertiller, M.B. Patterns of nitrogen conservation in shrubs and grasses in the Patagonian Monte, Argentina. Plant Soil 2000, 224, 185–193. [Google Scholar] [CrossRef]
- Bertiller, M.B.; Sain, C.L.; Carrera, A.L.; Vargas, D.N. Patterns of nitrogen and phosphorus conservation in dominant perennial grasses and shrubs across an aridity gradient in Patagonia, Argentina. J. Arid. Environ. 2005, 62, 209–223. [Google Scholar] [CrossRef]
- Yuan, Z.; Liu, W.; Niu, S.; Wan, S. Plant Nitrogen Dynamics and Nitrogen-use Strategies under Altered Nitrogen Seasonality and Competition. Ann. Bot. 2007, 100, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Barceló, M.; van Bodegom, P.M.; Soudzilovskaia, N.A. Climate drives the spatial distribution of mycorrhizal host plants in terrestrial ecosystems. J. Ecol. 2019, 107, 2564–2573. [Google Scholar] [CrossRef]
- Norby, R.J.; Long, T.M.; Hartz-Rubin, J.S.; O’Neill, E.G. Nitrogen resorption in senescing tree leaves in a warmer, CO2-enriched atmosephere. Plant Soil 2000, 224, 15–29. [Google Scholar] [CrossRef]
- Wright, I.J.; Westoby, M. Nutrient concentration, resorption and lifespan: Leaf traits of Australian sclerophyll species. Funct. Ecol. 2003, 17, 10–19. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Chen, S.P.; Wang, Z.W.; Chen, Q.S.; Bai, W.M. Nitrogen response efficiency increased monotonically with decreasing soil resource availability: A case study from a semiarid grassland in northern China. Oecologia 2006, 148, 564–572. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H. Soil water status influences plant nitrogen use: A case study. Plant Soil 2007, 301, 303–313. [Google Scholar] [CrossRef]
- Bennett, A.E.; Classen, A.T. Climate change influences mycorrhizal fungal–plant interactions, but conclusions are limited by geographical study bias. Ecology 2020, 101, e02978. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomes | Mycorrhiza Types | C (mg∙g−1) | N (mg∙g−1) | P (mg∙g−1) | N:P Ratio | C:N Ratio | C:P Ratio |
|---|---|---|---|---|---|---|---|
| Boreal | AM | 460.1 ± 27.2 b | 8.8 ± 3.6 b | 1.7 ± 0.3 a | 17.1 ± 10.4 a | 54.1 ± 21.1 a | 999.2 ± 742.4 a |
| AM + ECM | 501.4 ± 24.1 a | 11.7 ± 4.5 a | 1.4 ± 0.3 a | 18.6 ± 14.7 a | 40.9 ± 18.3 a | 1035.0 ± 365.6 a | |
| ECM | 512.3 ± 18.5 a | 7.3 ± 3.5 c | 1.4 ± 0.3 a | 11.8 ± 6.5 b | 50.3 ± 14.8 a | 1068.5 ± 554.3 a | |
| Temperate | AM | 494.5 ± 53.7 a | 10.5 ± 4.2 a | 1.3 ± 0.3 a | 17.0 ± 6.4 a | 54.6 ± 22.9 a | 702.1 ± 341.3 a |
| AM + ECM | 498.5 ± 47.6 a | 9.8 ± 3.5 a | 1.5 ± 0.7 a | 16.8 ± 6.8 a | 59.8 ± 22.4 a | 1216.0 ± 573.6 a | |
| ECM | 503.0 ± 37.0 a | 9.5 ± 3.5 a | 1.4 ± 0.9 a | 17.6 ± 9.4 a | 52.8 ± 23.4 a | 947.9 ± 795.4 a | |
| Tropical | AM | 420.5 ± 51.4 a | 12.2 ± 4.2 a | - | 28.1 ± 16.4 a | 39.7 ± 9.7 b | 1325.0 ± 842.9 a |
| AM + ECM | 466.5 ± 37.1 a | 13.3 ± 5.5 a | - | 33.4 ± 11.1 a | 53.0 ± 5.4 a | 1780.0 ± 739.8 a | |
| ECM | 439.0 ± 46.7 a | 12.7 ± 3.8 a | - | 28.5 ± 17.7 a | 35.3 ± 8.5 b | 2016.5 ± 2260.6 a |
| Plant Functional Types | Mycorrhiza Types | C (mg∙g−1) | N (mg∙g−1) | P (mg∙g−1) | N:P Ratio | C:N Ratio | C:P Ratio |
|---|---|---|---|---|---|---|---|
| Deciduous | AM | 434.7 ± 58.3 b | 11.5 ± 4.2 a | 1.6 ± 0.4 a | 16.6 ± 8.4 a | 43.3 ± 18.3 a | 817.6 ± 610.7 a |
| AM + ECM | 499.2 ± 37.5 a | 10.7 ± 4.5 a | 1.5 ± 0.5 a | 16.9 ± 11.1 a | 48.6 ± 15.8 a | 1165.8 ± 468.9 a | |
| ECM | 493.1 ± 36.9 a | 10.4 ± 2.9 a | 1.4 ± 0.4 a | 14.8 ± 10.1 a | 49.6 ± 14.7 a | 772.9 ± 533.5 a | |
| Evergreen | AM | 456.0 ± 68.3 b | 10.8 ± 4.2 a | 1.2 ± 0.2 a | 25.4 ± 16.5 a | 48.8 ± 17.6 a | 1534.6 ± 877.6 a |
| AM + ECM | 490.1 ± 46.3 a | 11.1 ± 4.2 a | 1.3 ± 0.2 a | 23.6 ± 11.6 a | 58.9 ± 26.0 a | 1427.9 ± 657.1 a | |
| ECM | 512.0 ± 39.2 a | 7.8 ± 3.9 b | 1.2 ± 0.3 a | 17.2 ± 9.9 b | 56.9 ± 32.5 a | 1555.7 ± 1043.6 a | |
| Broadleaf | AM | 437.8 ± 59.2 b | 11.3 ± 4.2 a | 1.5 ± 0.4 a | 21.2 ± 13.3 a | 44.6 ± 16.5 a | 1190.4 ± 830.5 ab |
| AM + ECM | 493.3 ± 46.1 a | 10.8 ± 4.4 a | 1.5 ± 0.5 a | 21.2 ± 12.0 a | 53.6 ± 17.9 a | 1459.1 ± 556.2 a | |
| ECM | 492.8 ± 39.9 a | 10.4 ± 3.0 a | 1.4 ± 0.4 a | 17.5 ± 11.2 a | 51.2 ± 20.8 a | 780.6 ± 523.4 b | |
| Conifer | AM | 535.3 ± 53.1 a | 10.3 ± 4.0 a | 1.4 ± 0.1 a | 20.1 ± 17.5 a | 60.7 ± 26.8 a | - |
| AM + ECM | 497.3 ± 29.7 a | 11.5 ± 4.3 a | 1.3 ± 0.2 a | 16.4 ± 10.2 a | 54.2 ± 33.8 a | 956.8 ± 532.9 | |
| ECM | 518.6 ± 30.7 a | 7.2 ± 3.7 b | 1.3 ± 0.3 a | 13.8 ± 7.2 a | 55.9 ± 30.0 a | 1733 ± 1080.8 |
| Leaf Index | Mycorrhizal Type | Leaf Shape | Mycorrhizal Type × Leaf Shape | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| C | 35.86 | <0.001 | 7.58 | 0.03 | 6.70 | 0.001 |
| N | 15.74 | <0.001 | 13.62 | 0.03 | 10.79 | <0.001 |
| P | 2.50 | 0.09 | 0.58 | 0.45 | 0.05 | 0.96 |
| N:P | 12.51 | <0.001 | 32.62 | <0.001 | 0.52 | 0.60 |
| C:N | 0.36 | 0.70 | 0.55 | 0.48 | 0.33 | 0.72 |
| C:P | 0.11 | 0.90 | 0.04 | 0.84 | 5.92 | 0.02 |
| Mycorrhizal Types | C and MAT | N and MAT | P and MAT | N:P and MAT | C:N and MAT | C:P and MAT |
|---|---|---|---|---|---|---|
| AM | −4.97 × 10−3 b | 2.39 × 10−2 b | −4.8 × 10−2 b | 0.88 c | −1.01 b | 18.46 b |
| AM + ECM | −4.1 × 10−3 a | 1.32 × 10−2 c | 7.87 × 10−3 a | 1.14 a | 0.73 a | 49.85 b |
| ECM | −1.51 × 10−3 a | 4.20 × 10−2 a | −7.80 × 10−3 a | 1.02 b | −1.02 a | 173.72 a |
| Mycorrhizal Types | C and MAP | N and MAP | P and MAP | N:P and MAP | C:N and MAP | C:P and MAP |
|---|---|---|---|---|---|---|
| AM | 2.49 × 10−6 a | 1.82 × 10−4 c | −1.89 × 10−4 b | 9.4 × 10−3 a | −1.16 × 10−2 b | 0.45 c |
| AM + ECM | −4.57 × 10−5 c | 1.32 × 10−2 a | 2.97 × 10−4 a | 8.25 × 10−3 b | −5.18 × 10−3 a | 0.58 b |
| ECM | −4.73 × 10−5 b | 3.06 × 10−4 b | −2.03 × 10−4 b | 4.08 × 10−3 c | −1.46 × 10−2 a | 0.77 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.-W.; Shi, Z.-Y.; Huang, M.; Yang, S.; Yang, W.-Y.; Li, Y.-J. Influence of Mycorrhiza on C:N:P Stoichiometry in Senesced Leaves. J. Fungi 2023, 9, 588. https://doi.org/10.3390/jof9050588
Wu S-W, Shi Z-Y, Huang M, Yang S, Yang W-Y, Li Y-J. Influence of Mycorrhiza on C:N:P Stoichiometry in Senesced Leaves. Journal of Fungi. 2023; 9(5):588. https://doi.org/10.3390/jof9050588
Chicago/Turabian StyleWu, Shan-Wei, Zhao-Yong Shi, Ming Huang, Shuang Yang, Wen-Ya Yang, and You-Jun Li. 2023. "Influence of Mycorrhiza on C:N:P Stoichiometry in Senesced Leaves" Journal of Fungi 9, no. 5: 588. https://doi.org/10.3390/jof9050588
APA StyleWu, S. -W., Shi, Z. -Y., Huang, M., Yang, S., Yang, W. -Y., & Li, Y. -J. (2023). Influence of Mycorrhiza on C:N:P Stoichiometry in Senesced Leaves. Journal of Fungi, 9(5), 588. https://doi.org/10.3390/jof9050588