
Effect of Different Light Wavelengths on Zymoseptoria tritici Development and Leaf Colonization in Bread Wheat

 ,
,  ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Inoculum Production
2.2. Light Treatments
2.3. In Vitro Mycelial Growth Assay under Different Light Wavelengths
2.4. Plant Material and Inoculation
2.5. Disease Evaluation
2.6. Z. tritici DNA Quantification
2.7. Statistical Analysis
3. Results
3.1. Effect of the Different Light Conditions on the Morphology and Growth of Z. tritici Mycelium
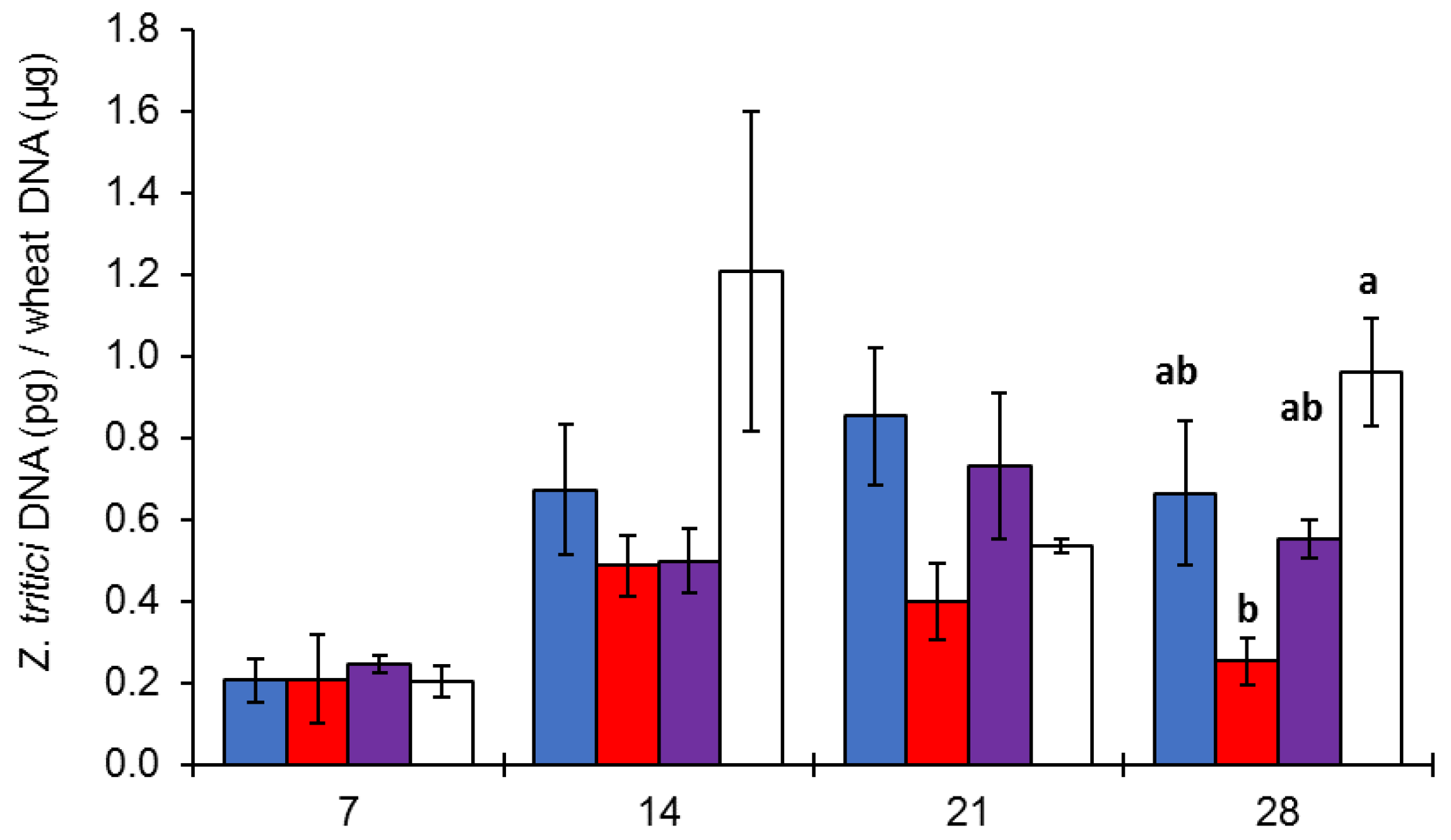
3.2. Effect of Light on Z. tritici Colonization of Bread Wheat Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sliney, D.H. What Is Light? The Visible Spectrum and Beyond. Eye 2016, 30, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Chory, J. Phytochrome Signaling Mechanisms and the Control of Plant Development. Trends Cell Biol. 2011, 21, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.M.; Blackwood, L.; Petersen, J.; Sullivan, S. Plant Flavoprotein Photoreceptors. Plant Cell Physiol. 2015, 56, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Mawphlang, O.I.L.; Kharshiing, E.V. Photoreceptor Mediated Plant Growth Responses: Implications for Photoreceptor Engineering toward Improved Performance in Crops. Front. Plant Sci. 2017, 8, 1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möglich, A.; Yang, X.; Ayers, R.A.; Moffat, K. Structure and Function of Plant Photoreceptors. Annu. Rev. Plant Biol. 2010, 61, 21–47. [Google Scholar] [CrossRef] [Green Version]
- Corrochano, L.M. Light in the Fungal World: From Photoreception to Gene Transcription and Beyond. Annu. Rev. Genet. 2019, 53, 149–170. [Google Scholar] [CrossRef]
- Herrera-Estrella, A.; Horwitz, B.A. Looking through the Eyes of Fungi: Molecular Genetics of Photoreception. Mol. Microbiol. 2007, 64, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Thind, T.S.; Schilder, A.C. Understanding Photoreception in Fungi and Its Role in Fungal Development with Focus on Phytopathogenic Fungi. Indian Phytopathol. 2018, 71, 169–182. [Google Scholar] [CrossRef]
- Yu, Z.; Fischer, R. Light Sensing and Responses in Fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef]
- Cerón-Bustamante, M.; Balducci, E.; Beccari, G.; Nicholson, P.; Covarelli, L.; Benincasa, P. Effect of Light Spectra on Cereal Fungal Pathogens, a Review. Fungal Biol. Rev. 2022, 43, 100291. [Google Scholar] [CrossRef]
- Kim, H.; Ridenour, J.B.; Dunkle, L.D.; Bluhm, B.H. Regulation of Pathogenesis by Light in Cercospora zeae-maydis: An Updated Perspective. Plant Pathol. J. 2011, 27, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.M.; Ramkumar, G.; Lee, Y.H. Light Quality Influences the Virulence and Physiological Responses of Colletotrichum acutatum Causing Anthracnose in Pepper Plants. J. Appl. Microbiol. 2013, 115, 509–516. [Google Scholar] [CrossRef]
- Islam, S.Z.; Honda, Y.; Arase, S. Light-Induced Resistance of Broad Bean against Botrytis cinerea. J. Phytopathol. 1998, 146, 479–485. [Google Scholar] [CrossRef]
- Telli, O.; Jimenez-Quiros, C.; McDowell, J.M.; Tör, M. Effect of Light and Dark on the Growth and Development of Downy Mildew Pathogen Hyaloperonospora arabidopsidis. Plant Pathol. 2020, 69, 1291–1300. [Google Scholar] [CrossRef]
- Solanki, S.; Ameen, G.; Borowicz, P.; Brueggeman, R.S. Shedding Light on Penetration of Cereal Host Stomata by Wheat Stem Rust Using Improved Methodology. Sci. Rep. 2019, 9, 7939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, J. How Light Affects the Life of Botrytis. Fungal Genet. Biol. 2017, 106, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Judelson, H.S. Myb Transcription Factors and Light Regulate Sporulation in the Oomycete Phytophthora infestans. PLoS ONE 2014, 9, e92086. [Google Scholar] [CrossRef] [Green Version]
- Balsells-Llauradó, M.; Torres, R.; Vall-llaura, N.; Casals, C.; Teixidó, N.; Usall, J. Light Intensity Alters the Behavior of Monilinia spp. in vitro and the Disease Development on Stone Fruit-Pathogen Interaction. Front. Plant Sci. 2021, 12, 666985. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Singh, P.; Park, J.; Park, S.; Friedman, A.; Zheng, T.; Lee, Y.H.; Lee, K. Genetic and Molecular Characterization of a Blue Light Photoreceptor MGWC-1 in Magnaporth oryzae. Fungal Genet. Biol. 2011, 48, 400–407. [Google Scholar] [CrossRef]
- Canessa, P.; Schumacher, J.; Hevia, M.A.; Tudzynski, P.; Larrondo, L.F. Assessing the Effects of Light on Differentiation and Virulence of the Plant Pathogen Botrytis cinerea: Characterization of the White Collar Complex. PLoS ONE 2013, 8, e84223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, J.; Simon, A.; Cohrs, K.C.; Viaud, M.; Tudzynski, P. The Transcription Factor BcLTF1 Regulates Virulence and Light Responses in the Necrotrophic Plant Pathogen Botrytis cinerea. PLoS Genet. 2014, 10, e1004040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fones, H.; Gurr, S. The Impact of Septoria Tritici Blotch Disease on Wheat: An EU Perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torriani, S.F.F.; Melichar, J.P.E.; Mills, C.; Pain, N.; Sierotzki, H.; Courbot, M. Zymoseptoria tritici: A Major Threat to Wheat Production, Integrated Approaches to Control. Fungal Genet. Biol. 2015, 79, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.E.; Goodwin, S.B. MVE1, Encoding the Velvet Gene Product Homolog in Mycosphaerella graminicola, Is Associated with Aerial Mycelium Formation, Melanin Biosynthesis, Hyphal Swelling, and Light Signaling. Appl. Environ. Microbiol. 2011, 77, 942–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiley, A.M.M.; Lawless, C.; Pilo, P.; Karki, S.J.; Lu, J.; Long, Z.; Gibriel, H.; Bailey, A.M.; Feechan, A. The Zymoseptoria tritici White Collar-1 Gene, ZtWco-1, Is Required for Development and Virulence on Wheat. Fungal Genet. Biol. 2022, 161, 103715. [Google Scholar] [CrossRef]
- Tiley, A.M.M.; White, H.J.; Foster, G.D.; Bailey, A.M. The ZtvelB Gene Is Required for Vegetative Growth and Sporulation in the Wheat Pathogen Zymoseptoria tritici. Front. Microbiol. 2019, 10, 2210. [Google Scholar] [CrossRef] [Green Version]
- Cascant-Lopez, E.; Crosthwaite, S.K.; Johnson, L.J.; Harrison, R.J. No Evidence That Homologs of Key Circadian Clock Genes Direct Circadian Programs of Development or MRNA Abundance in Verticillium dahliae. Front. Microbiol. 2020, 11, 1977. [Google Scholar] [CrossRef]
- McCorison, C.B.; Goodwin, S.B. The Wheat Pathogen Zymoseptoria tritici Senses and Responds to Different Wavelengths of Light. BMC Genom. 2020, 21, 513. [Google Scholar] [CrossRef]
- Kilaru, S.; Fantozzi, E.; Cannon, S.; Schuster, M.; Chaloner, T.M.; Guiu-Aragones, C.; Gurr, S.J.; Steinberg, G. Zymoseptoria tritici White-Collar Complex Integrates Light, Temperature and Plant Cues to Initiate Dimorphism and Pathogenesis. Nat. Commun. 2022, 13, 5625. [Google Scholar] [CrossRef]
- Tosti, G.; Benincasa, P.; Cortona, R.; Falcinelli, B.; Farneselli, M.; Guiducci, M.; Onofri, A.; Pannacci, E.; Tei, F.; Giulietti, M. Growing Lettuce under Multispectral Light-Emitting Diodes Lamps with Adjustable Light Intensity. Ital. J. Agron. 2017, 11, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Chungu, C.; Gilbert, J.; Townley-Smith, F. Septoria Tritici Blotch Development as Affected by Temperature, Duration of Leaf Wetness, Inoculum Concentration, and Host. Plant Dis. 2001, 85, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium Species, Chemotype Characterisation and Trichothecene Contamination of Durum and Soft Wheat in an Area of Central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.R.; Schnieder, F.; Verreet, J.A. Presymptomatic and Quantitative Detection of Mycosphaerella graminicola Development in Wheat Using a Real-Time PCR Assay. FEMS Microbiol. Lett. 2006, 262, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaisen, M.; Supronienė, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-Time PCR for Quantification of Eleven Individual Fusarium Species in Cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: https://www.R-project.org/ (accessed on 1 February 2023).
- Russell Lenth. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.5.2-1. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 February 2023).
- De Lucca, A.J.; Carter-Wientjes, C.; Williams, K.A.; Bhatnagar, D. Blue Light (470 nm) Effectively Inhibits Bacterial and Fungal Growth. Lett. Appl. Microbiol. 2012, 55, 460–466. [Google Scholar] [CrossRef] [Green Version]
- Tomb, R.M.; White, T.A.; Coia, J.E.; Anderson, J.G.; MacGregor, S.J.; Maclean, M. Review of the Comparative Susceptibility of Microbial Species to Photoinactivation Using 380-480 nm Violet-Blue Light. Photochem. Photobiol. 2018, 94, 445–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzaska, W.J.; Wrigley, H.E.; Thwaite, J.E.; May, R.C. Species-Specific Antifungal Activity of Blue Light. Sci. Rep. 2017, 7, 4605. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, P.; Huang, C.; Wang, W. Dual Effect of Blue Light on Fusarium solani Clinical Corneal Isolates in vitro. Lasers Med Sci. 2020, 35, 1299–1305. [Google Scholar] [CrossRef]
- Lafuente, M.T.; Alférez, F. Effect of LED Blue Light on Penicillium digitatum and Penicillium italicum Strains. Photochem. Photobiol. 2015, 91, 1412–1421. [Google Scholar] [CrossRef]
- Cheng, C.W.; Chen, C.K.; Chang, C.J.; Chen, L.Y. Effect of Colour LEDs on Mycelia Growth of Aspergillus ficuum and Phytase Production in Photo-Fermentations. J. Photochem. Photobiol. B Biol. 2012, 106, 81–86. [Google Scholar] [CrossRef]
- Haridas, D.; Atreya, C.D. The Microbicidal Potential of Visible Blue Light in Clinical Medicine and Public Health. Front. Med. 2022, 9, 606. [Google Scholar] [CrossRef]
- Imada, K.; Tanaka, S.; Ibaraki, Y.; Yoshimura, K.; Ito, S. Antifungal Effect of 405-nm Light on Botrytis cinerea. Lett. Appl. Microbiol. 2014, 59, 670–676. [Google Scholar] [CrossRef]
- Madden, L.V.; Hughes, G.; Van Den Bosch, F. The Study of Plant Disease Epidemics; The American Phytopathological Society: Saint Paul, MN, USA, 2017; ISBN 978-0-89054-505-8. [Google Scholar]
- Lovell, D.J.; Hunter, T.; Powers, S.J.; Parker, S.R.; Van Den Bosch, F. Effect of Temperature on Latent Period of Septoria Leaf Blotch on Winter Wheat under Outdoor Conditions. Plant Pathol. 2004, 53, 170–181. [Google Scholar] [CrossRef]
- Pfender, W.F. Role of Phenology in Host Susceptibility and Within-Plant Spread of Stem Rust During Reproductive Development of Perennial Ryegrass. Phytopathology 2007, 94, 308–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Précigout, P.-A.; Claessen, D.; Makowski, D.; Robert, C. Does the Latent Period of Leaf Fungal Pathogens Reflect Their Trophic Type? A Meta-Analysis of Biotrophs, Hemibiotrophs, and Necrotrophs. Phytopathology 2020, 110, 345–361. [Google Scholar] [CrossRef] [Green Version]
- Lannou, C. Variation and Selection of Quantitative Traits in Plant Pathogens. Annu. Rev. Phytopathol. 2012, 50, 319–338. [Google Scholar] [CrossRef]
- Fantozzi, E.; Kilaru, S.; Gurr, S.J.; Steinberg, G. Asynchronous Development of Zymoseptoria tritici Infection in Wheat. Fungal Genet. Biol. 2021, 146, 103504. [Google Scholar] [CrossRef] [PubMed]
- Fones, H.N.; Eyles, C.J.; Kay, W.; Cowper, J.; Gurr, S.J. A Role for Random, Humidity-Dependent Epiphytic Growth Prior to Invasion of Wheat by Zymoseptoria tritici. Fungal Genet. Biol. 2017, 106, 51–60. [Google Scholar] [CrossRef]
- Islam, S.; Babadoost, M.; Honda, Y.; Sawa, Y. Characterization of Antifungal Glycoprotein in Red-Light-Irradiated Broadbean Leaflets. Mycoscience 2002, 43, 471–473. [Google Scholar] [CrossRef]
- Xu, H.; Fu, Y.N.; Li, T.L.; Wang, R. Effects of Different LED Light Wavelengths on the Resistance of Tomato against Botrytis cinerea and the Corresponding Physiological Mechanisms. J. Integr. Agric. 2017, 16, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jiang, Y.P.; Yu, H.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Light Quality Affects Incidence of Powdery Mildew, Expression of Defence-Related Genes and Associated Metabolism in Cucumber Plants. Eur. J. Plant Pathol. 2010, 127, 125–135. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Khanam, H.; Ueno, M.; Kihara, J.; Honda, Y.; Arase, S. Suppression by Red Light Irradiation of Corynespora Leaf Spot of Cucumber Caused by Corynespora cassiicola. J. Phytopathol. 2010, 158, 378–381. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Honda, Y.; Arase, S. Red-Light-Induced Resistance in Broad Bean (Vicia faba L.) to Leaf Spot Disease Caused by Alternaria tenuissima. J. Phytopathol. 2003, 151, 86–91. [Google Scholar] [CrossRef]
- Shirasawa, H.; Ueno, M.; Kihara, J.; Arase, S. Protective Effect of Red Light against Blast Disease Caused by Magnaporthe oryzae in Rice. Crop Prot. 2012, 39, 41–44. [Google Scholar] [CrossRef]
- Ueno, M.; Imaoka, A.; Kihara, J.; Arase, S. Effects of Light Quality on Induction of Tryptamine-Mediated Resistance in Lesion Mimic Mutant of Rice Infected with Magnaporthe grisea. J. Phytopathol. 2007, 155, 228–235. [Google Scholar] [CrossRef]
- Parada, R.Y.; Mon-nai, W.; Ueno, M.; Kihara, J.; Arase, S. Red-Light-Induced Resistance to Brown Spot Disease Caused by Bipolaris oryzae in Rice. J. Phytopathol. 2015, 163, 116–123. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Tóth, L.; Galgóczy, L.; Poór, P. Pest and Disease Management by Red Light. Plant. Cell Environ. 2021, 44, 3197–3210. [Google Scholar] [CrossRef]
- Genoud, T.; Buchala, A.J.; Chua, N.-H.; Métraux, J.-P. Phytochrome Signalling Modulates the SA-Perceptive Pathway in Arabidopsis. Plant J. 2002, 31, 87–95. [Google Scholar] [CrossRef]
- Griebel, T.; Zeier, J. Light Regulation and Daytime Dependency of Inducible Plant Defenses in Arabidopsis: Phytochrome Signaling Controls Systemic Acquired Resistance Rather Than Local Defense. Plant Physiol. 2008, 147, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Robson, F.; Okamoto, H.; Patrick, E.; Harris, S.-R.; Wasternack, C.; Brearley, C.; Turner, J.G. Jasmonate and Phytochrome A Signaling in Arabidopsis Wound and Shade Responses Are Integrated through JAZ1 Stability. Plant Cell 2010, 22, 1143–1160. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.-Z.; Xue, Y.-J.; Zhou, J.-J.; Zhang, B.; Chang, H.; Takano, M. Phytochromes Regulate SA and JA Signaling Pathways in Rice and Are Required for Developmentally Controlled Resistance to Magnaporthe grisea. Mol. Plant 2011, 4, 688–696. [Google Scholar] [CrossRef]
- Westermann, A.J.; Gorski, S.A.; Vogel, J. Dual RNA-Seq of Pathogen and Host. Nat. Rev. Microbiol. 2012, 10, 618–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic Acid Biosynthesis in Plants. Front. Plant Sci. 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Kramer, B.; Thines, E.; Foster, A.J. MAP Kinase Signalling Pathway Components and Targets Conserved between the Distantly Related Plant Pathogenic Fungi Mycosphaerella graminicola and Magnaporthe grisea. Fungal Genet. Biol. 2009, 46, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Mehrabi, R.; van der Lee, T.; Waalwijk, C.; Kema, G.H.J. MgSlt2, a Cellular Integrity MAP Kinase Gene of the Fungal Wheat Pathogen Mycosphaerella graminicola, Is Dispensable for Penetration but Essential for Invasive Growth. Mol. Plant-Microbe Interact. 2006, 19, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Mirzadi Gohari, A.; Mehrabi, R.; Robert, O.; Ince, I.A.; Boeren, S.; Schuster, M.; Steinberg, G.; de Wit, P.J.G.M.; Kema, G.H.J. Molecular Characterization and Functional Analyses of ZtWor1, a Transcriptional Regulator of the Fungal Wheat Pathogen Zymoseptoria tritici. Mol. Plant Pathol. 2014, 15, 394–405. [Google Scholar] [CrossRef]
- Marshall, R.; Kombrink, A.; Motteram, J.; Loza-Reyes, E.; Lucas, J.; Hammond-Kosack, K.E.; Thomma, B.P.H.J.; Rudd, J.J. Analysis of Two in Planta Expressed LysM Effector Homologs from the Fungus Mycosphaerella graminicola Reveals Novel Functional Properties and Varying Contributions to Virulence on Wheat. Plant Physiol. 2011, 156, 756–769. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerón-Bustamante, M.; Tini, F.; Beccari, G.; Benincasa, P.; Covarelli, L. Effect of Different Light Wavelengths on Zymoseptoria tritici Development and Leaf Colonization in Bread Wheat. J. Fungi 2023, 9, 670. https://doi.org/10.3390/jof9060670
Cerón-Bustamante M, Tini F, Beccari G, Benincasa P, Covarelli L. Effect of Different Light Wavelengths on Zymoseptoria tritici Development and Leaf Colonization in Bread Wheat. Journal of Fungi. 2023; 9(6):670. https://doi.org/10.3390/jof9060670
Chicago/Turabian StyleCerón-Bustamante, Minely, Francesco Tini, Giovanni Beccari, Paolo Benincasa, and Lorenzo Covarelli. 2023. "Effect of Different Light Wavelengths on Zymoseptoria tritici Development and Leaf Colonization in Bread Wheat" Journal of Fungi 9, no. 6: 670. https://doi.org/10.3390/jof9060670
APA StyleCerón-Bustamante, M., Tini, F., Beccari, G., Benincasa, P., & Covarelli, L. (2023). Effect of Different Light Wavelengths on Zymoseptoria tritici Development and Leaf Colonization in Bread Wheat. Journal of Fungi, 9(6), 670. https://doi.org/10.3390/jof9060670