
Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata)


Abstract
:1. Introduction
2. Materials and Methods
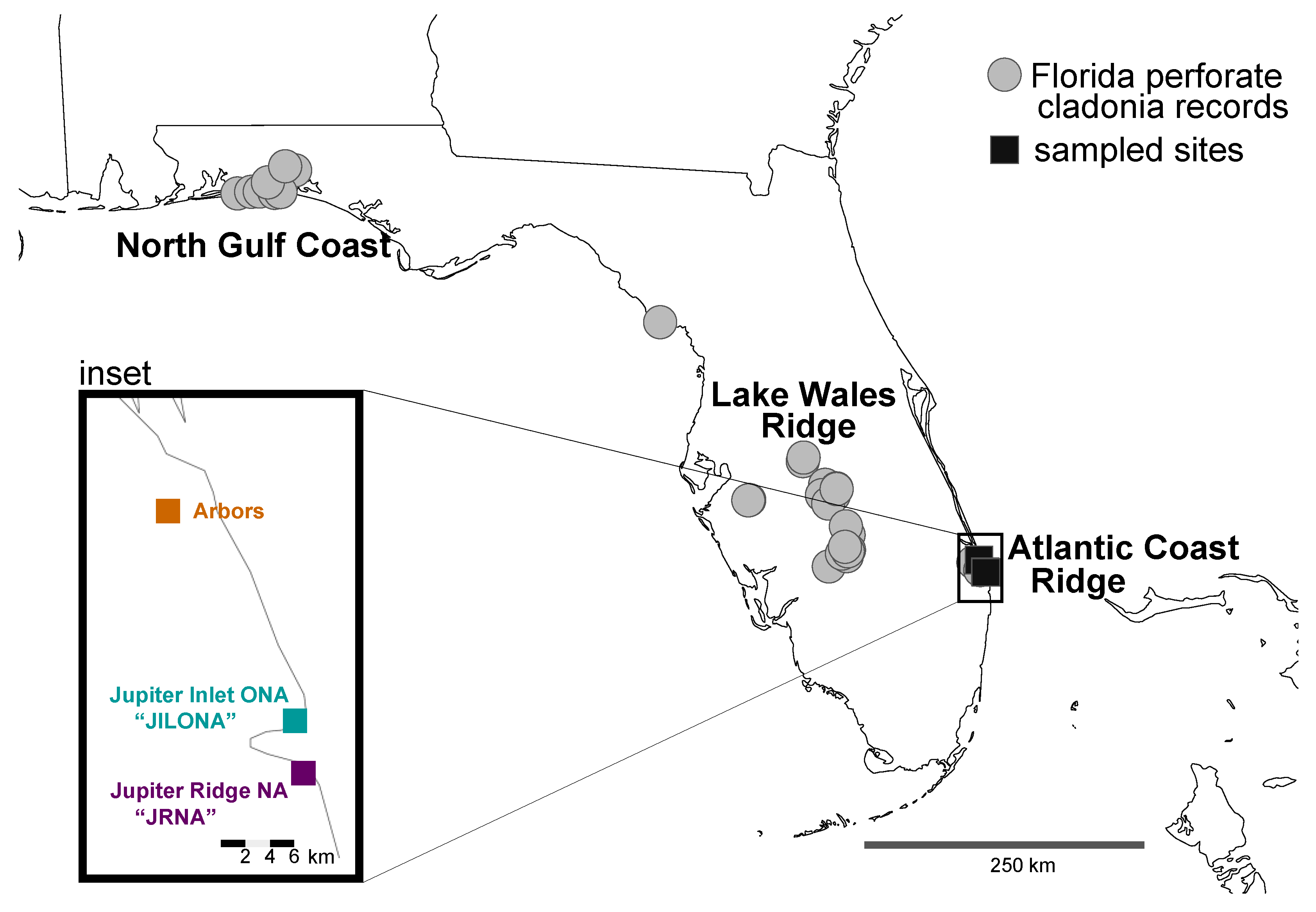
2.1. Field Sampling
2.2. High-Throughput Sequencing and Genome Assembly
2.3. Phylogenetic Datasets and Tree Inference
3. Results
3.1. Genomic Data
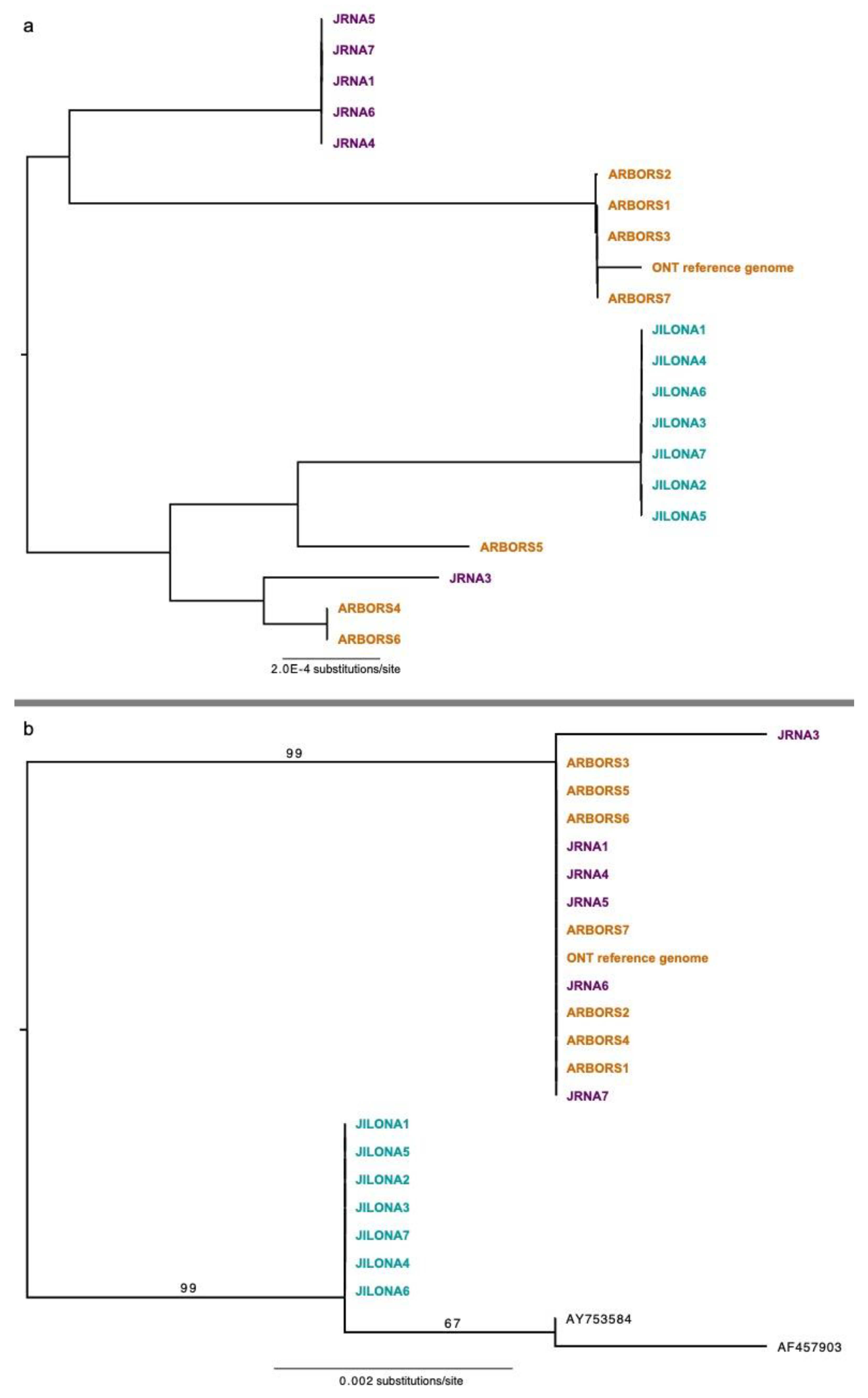
3.2. Phylogenomic Inferences
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Estill, J.C.; Cruzan, M.B. Phytogeography of rare plant species endemic to the Southeastern United States. Castanea 2001, 66, 3–23. [Google Scholar]
- Myers, R.L. Scrub and high pine. In Ecosystems of Florida; University of Central Florida Press: Orlando, FL, USA, 1990; Volume 765, pp. 150–193. [Google Scholar]
- Laessle, A.M. Relationship of sand pine scrub to former shore lines. Q. J. Fla. Acad. Sci. 1967, 30, 269–286. [Google Scholar]
- McCoy, E.D.; Britt, E.J.; Catenazzi, A.; Mushinsky, H.R. Fire and herpetofaunal diversity in the Florida scrub ecosystem. Nat. Areas J. 2013, 33, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Christman, S.P.; Judd, W.S. Notes on plants endemic to Florida scrub. Fla. Sci. 1990, 53, 52–73. [Google Scholar]
- Germain-Aubrey, C.C.; Soltis, P.S.; Neubig, K.M.; Thurston, T.; Soltis, D.E.; Gitzendanner, M.A. Using comparative biogeography to retrace the origins of an ecosystem: The case of four plants endemic to the Central Florida scrub. Int. J. Plant Sci. 2014, 175, 418–431. [Google Scholar] [CrossRef]
- Deyrup, M. Arthropods endemic to Florida scrub. Fla. Sci. 1989, 52, 254–270. [Google Scholar]
- Wilhelm, G.S.; Burkhalter, J.R. Cladonia Perforata, the Northwest Florida population. Bryologist 1990, 93, 66–68. [Google Scholar] [CrossRef]
- Noss, R.F.; Platt, W.J.; Sorrie, B.A.; Weakley, A.S.; Means, D.B.; Costanza, J.; Peet, R.K. How global biodiversity hotspots may go unrecognized: Lessons from the North American Coastal Plain. Divers. Distrib. 2015, 21, 236–244. [Google Scholar] [CrossRef]
- Menges, E.S.; Main, K.N.; Pickert, R.L.; Ewing, K. Evaluating a fire management plan for fire regime goals in a Florida landscape. Nat. Areas J. 2017, 37, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Yahr, R. Ecology and post-fire recovery of Cladonia perforata, an endangered Florida-scrub lichen. For. Snow Landsc. Res. 2000, 75, 339–356. [Google Scholar]
- Lindsay, M.N.; Lewis, D.B.; Halstead, N.; Gainsbury, A.M. Fire severity effects on the herpetofaunal diversity of the Florida scrub, a biodiversity hotspot. Biodivers. Conserv. 2023, 32, 1857–1878. [Google Scholar] [CrossRef]
- McCoy, E.D.; Mushinsky, H.R. Effects of fragmentation on the richness of vertebrates in the Florida scrub habitat. Ecology 1994, 75, 446–457. [Google Scholar] [CrossRef]
- Li, J.; Powel, T.L.; Seiler, T.J.; Johnson, D.P.; Anderson, H.P.; Bracho, R.; Hungate, B.A.; Hinkle, C.R.; Drake, B.G. Impacts of urricane Frances on Florida scrub-oak ecosystem processes: Defoliation, net CO2 exchange and interactions with elevated CO2. Glob. Chang. Biol. 2007, 13, 1101–1113. [Google Scholar] [CrossRef]
- Menges, E.S.; Kohfeldt, N. Life history strategies of Florida scrub plants in relation to rire. Bull. Torrey Bot. Club 1995, 122, 282–297. [Google Scholar] [CrossRef]
- Rosentreter, R.; DeBolt, A. Lichens as an indicator of sea-level rise. Evansia 2023, 39, 162–168. [Google Scholar] [CrossRef]
- Yahr, R.; Vilgalys, R.; Depriest, P.T. Strong fungal specificity and selectivity for algal symbionts in Florida scrub Cladonia lichens. Mol. Ecol. 2004, 13, 3367–3378. [Google Scholar] [CrossRef]
- DeBolt, A. Transplant Success of Cladonia perforata (Florida Perforate Cladonia) at the Jupiter Inlet Lighthouse Outstanding Natural Area. Evansia 2021, 38, 32–42. [Google Scholar] [CrossRef]
- Buckley, A.; Hendrickson, T.O. The distribution of Cladonia perforata Evans on the southern Lake Wales Ridge in Highlands County, Florida. Bryologist 1988, 91, 354–356. [Google Scholar] [CrossRef]
- Yahr, R.L.S.G. Cladonia perforata. The IUCN Red List of Threatened Species 2003: E.T43994A10838980. Available online: file:///C:/Users/MDPI/Downloads/10.2305_IUCN.UK.2003.RLTS.T43994A10838980.en-1.pdf (accessed on 28 April 2023). [CrossRef]
- Yahr, R. Evolutionary and Ecological Pattern and Process in Cladonia Lichen Symbioses: A Population Genetic Approach. Ph.D. Thesis, Duke University, Durham, NC, USA, 2004. [Google Scholar]
- Barrett, S.C.H. Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 8859–8866. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.L.; McMullin, R.T.; Tripp, E.A.; Lendemer, J.C. Lichen conservation in North America: A review of current practices and research in Canada and the United States. Biodivers. Conserv. 2019, 28, 3103–3138. [Google Scholar] [CrossRef]
- Formenti, G.; Theissinger, K.; Fernandes, C.; Bista, I.; Bombarely, A.; Bleidorn, C.; Ciofi, C.; Crottini, A.; Godoy, J.A.; Höglund, J.; et al. The era of reference genomes in conservation genomics. Trends Ecol. Evol. 2022, 37, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Holt, K.E. Polypolish: Short-read polishing of long-read bacterial genome assemblies. PLoS Comput. Biol. 2022, 18, e1009802. [Google Scholar] [CrossRef] [PubMed]
- Zimin, A.V.; Salzberg, S.L. The genome polishing tool POLCA makes fast and accurate corrections in genome assemblies. PLoS Comput. Biol. 2020, 16, e1007981. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [Green Version]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Bertels, F.; Silander, O.K.; Pachkov, M.; Rainey, P.B.; van Nimwegen, E. Automated reconstruction of whole-genome phylogenies from short-sequence reads. Mol. Biol. Evol. 2014, 31, 1077–1088. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Rozewicki, J.; Yamada, K.D.; Katoh, K. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, K.; Steenkamp, E.T.; Coetzee, M.P.A.; Wingfield, M.J.; Wingfield, B.D. Concerted evolution in the ribosomal RNA cistron. PLoS ONE 2013, 8, e59355. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree Version 1.4. 2008. Available online: http://tree.bio.ed.ac.uk/software/Wgtree/ (accessed on 9 January 2023).
- Pino-Bodas, R.; Stenroos, S. Global biodiversity patterns of the photobionts associated with the genus Cladonia (Lecanorales, Ascomycota). Microb. Ecol. 2021, 82, 173–187. [Google Scholar] [CrossRef]
- Blaxter, M.; Archibald, J.M.; Childers, A.K.; Coddington, J.A.; Crandall, K.A.; Di Palma, F.; Durbin, R.; Edwards, S.V.; Graves, J.A.M.; Hackett, K.J.; et al. Why sequence all eukaryotes? Proc. Natl. Acad. Sci. USA 2022, 119, e2115636118. [Google Scholar] [CrossRef]
- Pfeffer, B.; Lymbery, C.; Booth, B.; Allen, J.L. Chromosomal genome sequence assembly and mating-type (MAT) locus characterization of the leprose asexual lichenized fungus Lepraria neglecta (Nyl.) Erichsen. Lichenologist 2023, 55, 41–50. [Google Scholar] [CrossRef]
- Park, S.-Y.; Choi, J.; Lee, G.-W.; Kim, J.A.; Oh, S.-O.; Jeong, M.-H.; Yu, N.-H.; Kim, S.; Lee, Y.-H.; Hur, J.-S. Draft genome sequence of lichen-forming fungus Cladonia metacorallifera Strain KoLRI002260. Genome Announc. 2014, 2, e01065-13. [Google Scholar] [CrossRef] [Green Version]
- Brigham, L.M.; Allende, L.M.; Shipley, B.R.; Boyd, K.C.; Higgins, T.J.; Kelly, N.; Anderson Stewart, C.R.; Keepers, K.G.; Pogoda, C.S.; Lendemer, J.C.; et al. Genomic insights into the mitochondria of 11 eastern North American species of Cladonia. Mitochondrial DNA Part B 2018, 3, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widhelm, T.J.; Grewe, F.; Huang, J.-P.; Ramanauskas, K.; Mason-Gamer, R.; Lumbsch, H.T. Using RADseq to understand the circum-Antarctic distribution of a lichenized fungus, Pseudocyphellaria glabra. J. Biogeogr. 2021, 48, 78–90. [Google Scholar] [CrossRef]
- Lücking, R.; Leavitt, S.D.; Hawksworth, D.L. Species in lichen-forming fungi: Balancing between conceptual and practical considerations, and between phenotype and phylogenomics. Fungal Divers. 2021, 109, 99–154. [Google Scholar] [CrossRef]
- Grewe, F.; Lagostina, E.; Wu, H.; Printzen, C.; Lumbsch, H.T. Population genomic analyses of RAD sequences resolves the phylogenetic relationship of the lichen-forming fungal species Usnea antarctica and Usnea aurantiacoatra. MycoKeys 2018, 43, 91–113. [Google Scholar] [CrossRef] [Green Version]
- Hale, H.; Gardner, E.M.; Viruel, J.; Pokorny, L.; Johnson, M.G. Strategies for reducing per-sample costs in target capture sequencing for phylogenomics and population genomics in plants. Appl. Plant Sci. 2020, 8, e11337. [Google Scholar] [CrossRef] [Green Version]
- Widhelm, T.J.; Grewe, F.; Huang, J.-P.; Mercado-Díaz, J.A.; Goffinet, B.; Lücking, R.; Moncada, B.; Mason-Gamer, R.; Lumbsch, H.T. Multiple historical processes obscure phylogenetic relationships in a taxonomically difficult group (Lobariaceae, Ascomycota). Sci. Rep. 2019, 9, 8968. [Google Scholar] [CrossRef] [Green Version]
- DeSalle, R.; Goldstein, P. Review and interpretation of trends in DNA barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.R.; Karol, K.G.; Ohmura, Y.; Pogoda, C.S.; Keepers, K.G.; McMullin, R.T.; Lendemer, J.C. Mitochondrial genomes in the iconic reindeer lichens: Architecture, variation, and synteny across multiple evolutionary scales. Mycologia 2023, 115, 187–205. [Google Scholar] [CrossRef]
- Allen, J.L.; McKenzie, S.K.; Sleith, R.S.; Alter, S.E. First genome-wide analysis of the endangered, endemic lichen Cetradonia linearis reveals isolation by distance and strong population structure. Am. J. Bot. 2018, 105, 1556–1567. [Google Scholar] [CrossRef]
- Alonso-García, M.; Grewe, F.; Payette, S.; Villarreal, J.C.A. Population genomics of a reindeer lichen species from North American lichen woodlands. Am. J. Bot. 2021, 108, 159–171. [Google Scholar] [CrossRef]
- Alors, D.; Grande, F.D.; Cubas, P.; Crespo, A.; Schmitt, I.; Molina, M.C.; Divakar, P.K. Panmixia and dispersal from the Mediterranean Basin to Macaronesian Islands of a macrolichen species. Sci. Rep. 2017, 7, 40879. [Google Scholar] [CrossRef]
- Werth, S.; Gugerli, F.; Holderegger, R.; Wagner, H.H.; Csencsics, D.; Scheidegger, C. Landscape-level gene flow in Lobaria pulmonaria, an epiphytic lichen. Mol. Ecol. 2007, 16, 2807–2815. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.L.; McMullin, R.T.; Wiersma, Y.F.; Scheidegger, C. Population genetics and biogeography of the lungwort lichen in North America support distinct Eastern and Western gene pools. Am. J. Bot. 2021, 108, 2416–2424. [Google Scholar] [CrossRef]
- Menges, E.S.; Hawkes, C.V. Interactive effects of fire and microhabitat on plants of Florida scrub. Ecol. Appl. 1998, 8, 935–946. [Google Scholar] [CrossRef]
- Quintana-Ascencio, P.F.; Morales-Hernández, M. Fire-mediated effects of shrubs, lichens and herbs on the demography of Hypericum cumulicola in patchy Florida scrub. Oecologia 1997, 112, 263–271. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| # of Contigs | 72 |
|---|---|
| Total size | 33.61 Mb |
| Longest/shortest contig | 1.75 Mb/2 Kb |
| >10 kb/>100 kb/>1 Mb | 53/41/14 |
| N50/L50 | 1.1 Mb/12 |
| GC content | 47.05% |
| BUSCO complete single | 1250/1315 (95.1%) |
| BUSCO complete duplicate | 2/1315 (0.2%) |
| BUSCO fragmented | 40/1315 (3.0%) |
| BUSCO missing | 23/1315 (1.7%) |
| Total annotations | 13,980 |
| CAZYmes | 215 |
| antiSMASH clusters | 48 |
| antiSMASH biosynthetic enzymes | 99 |
| antiSMASH smCOGs | 141 |
| Data Matrix | MSA Size | Genetic Groups | Pairwise Identity (Identical Sites) |
|---|---|---|---|
| Nuclear Genome | 32.1 Mb | 6 | NA |
| Coding loci | 12.1 Mb | 6 | NA |
| BUSCO loci | 1.91 Mb | 6 | NA |
| Cistron | 9777 bp | 2 | 99.7% (99.3%) |
| ITS | 565 bp | 2 (3) 1 | 99.5% (98.1%) |
| Mitochondrial Genome | 59,014 bp | 1 | 100% (100%) |
| Sample Code | Illumina Reads | Nuclear Coverage | mt Coverage | nrDNA Coverage |
|---|---|---|---|---|
| Arbors1 | 3,204,340 | 9× | 169× | 32× |
| Arbors2 | 5,439,708 | 15× | 251× | 50× |
| Arbors3 | 4,727,072 | 12× | 186× | 65× |
| Arbors4 | 5,475,152 | 14× | 287× | 36× |
| Arbors5 | 1,927,270 | 4× | 61× | 11× |
| Arbors6 | 3,623,778 | 8× | 130× | 35× |
| Arbors7 1 | 3,646,488 | 10× | 159× | 53× |
| JILONA1 | 4,254,622 | 9× | 187× | 54× |
| JILONA2 | 4,915,262 | 12× | 185× | 64× |
| JILONA3 | 6,474,484 | 15× | 271× | 66× |
| JILONA4 | 4,708,534 | 12× | 215× | 52× |
| JILONA5 | 4,270,992 | 11× | 175× | 72× |
| JILONA6 | 4,627,850 | 11× | 209× | 36× |
| JILONA7 | 2,052,574 | 4× | 84× | 26× |
| JRNA1 | 4,086,004 | 9× | 170× | 16× |
| JRNA2 2 | 4,827,906 | 12× | 165× | 94× |
| JRNA3 | 3,255,906 | 8× | 133× | 26× |
| JRNA4 | 5,457,804 | 13× | 230× | 29× |
| JRNA5 | 4,800,882 | 12× | 225× | 30× |
| JRNA6 | 3,589,860 | 8× | 207× | 18× |
| JRNA7 | 5,092,998 | 13× | 245× | 40× |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leavitt, S.D.; DeBolt, A.; McQuhae, E.; Allen, J.L. Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata). J. Fungi 2023, 9, 698. https://doi.org/10.3390/jof9070698
Leavitt SD, DeBolt A, McQuhae E, Allen JL. Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata). Journal of Fungi. 2023; 9(7):698. https://doi.org/10.3390/jof9070698
Chicago/Turabian StyleLeavitt, Steven D., Ann DeBolt, Ethan McQuhae, and Jessica L. Allen. 2023. "Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata)" Journal of Fungi 9, no. 7: 698. https://doi.org/10.3390/jof9070698
APA StyleLeavitt, S. D., DeBolt, A., McQuhae, E., & Allen, J. L. (2023). Genomic Resources for the First Federally Endangered Lichen: The Florida Perforate Cladonia (Cladonia perforata). Journal of Fungi, 9(7), 698. https://doi.org/10.3390/jof9070698