
Ubiquitin Degradation of the AICAR Transformylase/IMP Cyclohydrolase Ade16 Regulates the Sexual Reproduction of Cryptococcus neoformans



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Quantitative Real-Time PCR
2.3. Generation of Fluorescence Strains
2.4. Yeast Two-Hybrid Assays
2.5. Generation of Tagged Protein Strains
2.6. Generation of ADE16 Interference and Overexpression Strains
2.7. Analysis of Melanin and Capsule
2.8. Mating Assay
2.9. Statistical Analysis
3. Results
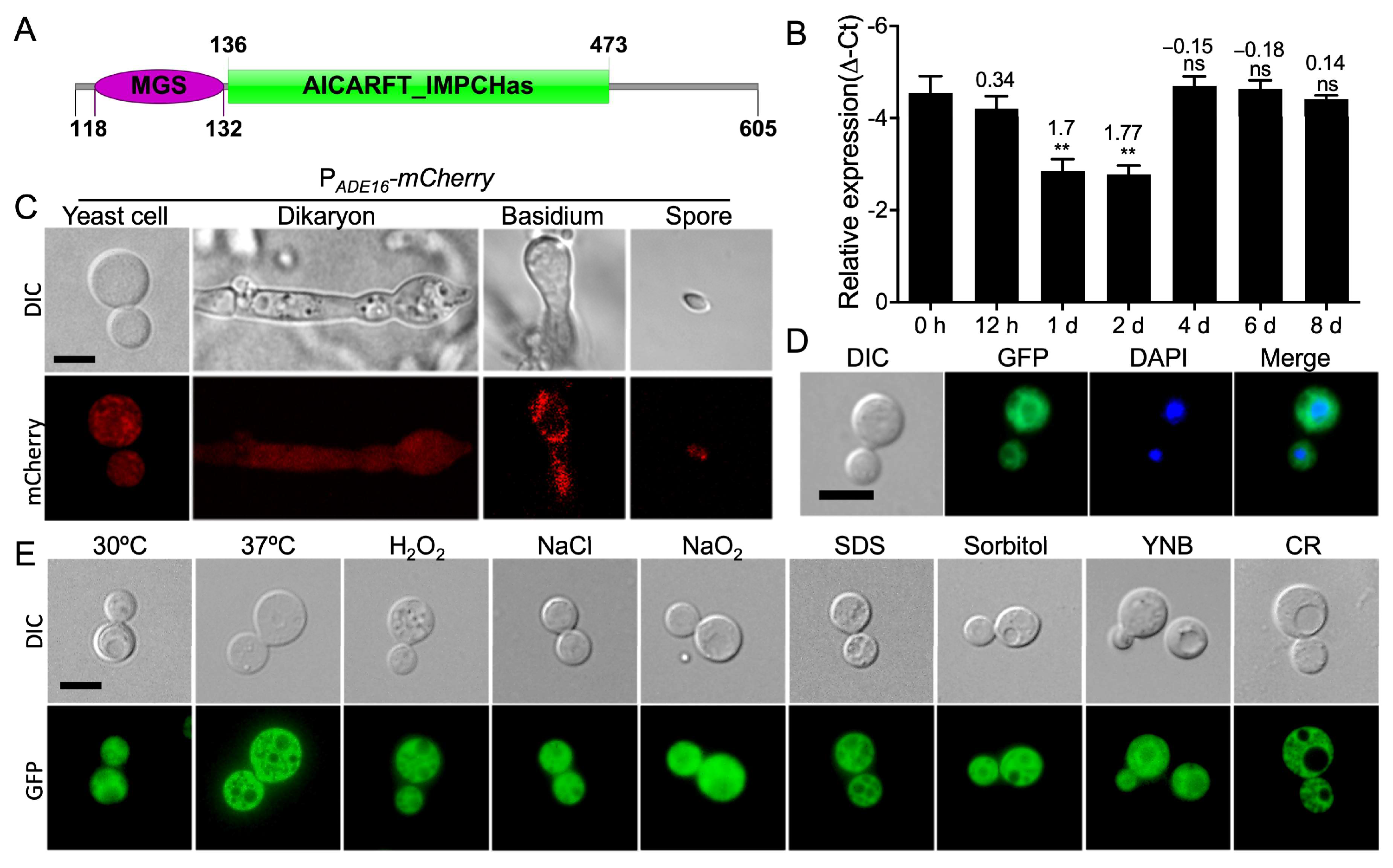
3.1. Interaction Profile and Expression Pattern of ADE16 during Mating of C. neoformans
3.2. Ade16 Interacts with Fbp1
3.3. The Ade16 Stability Depends on Fbp1 Function
3.4. ADE16 Is Required for Normal Growth of C. neoformans
3.5. ADE16 Overexpression Increases Capsule Size in C. neoformans
3.6. Ade16 Regulates Sexual Reproduction of C. neoformans
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Idnurm, A.; Bahn, Y.S.; Nielsen, K.; Lin, X.; Fraser, J.A.; Heitman, J. Deciphering the model pathogenic fungus Cryptococcus neoformans. Nat. Rev. Microbiol. 2005, 3, 753–764. [Google Scholar] [CrossRef]
- Beardsley, J.; Sorrell, T.C.; Chen, S.C. Central Nervous System Cryptococcal Infections in Non-HIV Infected Patients. J. Fungi 2019, 5, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.L.; Alviano, C.S.; Travassos, L.R. Pathogenicity of Cryptococcus neoformans: Virulence factors and immunological mechanisms. Microbes Infect. 1999, 1, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Govender, N.P.; Jordan, A.; Loyse, A.; Shroufi, A.; Denning, D.W.; Meya, D.B.; Chiller, T.M.; Boulware, D.R. The global burden of HIV-associated cryptococcal infection in adults in 2020: A modelling analysis. Lancet Infect. Dis. 2022, 22, 1748–1755. [Google Scholar] [CrossRef] [PubMed]
- Alspaugh, J.A.; Davidson, R.C.; Heitman, J. Morphogenesis of Cryptococcus neoformans. Contrib. Microbiol. 2000, 5, 217–238. [Google Scholar] [CrossRef]
- Kozubowski, L.; Heitman, J. Profiling a killer, the development of Cryptococcus neoformans. FEMS Microbiol. Rev. 2012, 36, 78–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Heitman, J. The biology of the Cryptococcus neoformans species complex. Annu. Rev. Microbiol. 2006, 60, 69–105. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, J.; Fan, Y.; Lin, X. Life Cycle of Cryptococcus neoformans. Annu. Rev. Microbiol. 2019, 73, 17–42. [Google Scholar] [CrossRef]
- Lin, X.; Hull, C.M.; Heitman, J. Sexual reproduction between partners of the same mating type in Cryptococcus neoformans. Nature 2005, 434, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Sun, S.; Billmyre, R.B.; Roach, K.C.; Heitman, J. Unisexual versus bisexual mating in Cryptococcus neoformans: Consequences and biological impacts. Fungal Genet. Biol. 2015, 78, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lin, X. Mechanisms of unisexual mating in Cryptococcus neoformans. Fungal Genet. Biol. 2011, 48, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.C.; Nichols, C.B.; Cox, G.M.; Perfect, J.R.; Heitman, J. A MAP kinase cascade composed of cell type specific and non-specific elements controls mating and differentiation of the fungal pathogen Cryptococcus neoformans. Mol. Microbiol. 2003, 49, 469–485. [Google Scholar] [CrossRef]
- Lin, X.; Jackson, J.C.; Feretzaki, M.; Xue, C.; Heitman, J. Transcription factors Mat2 and Znf2 operate cellular circuits orchestrating opposite- and same-sex mating in Cryptococcus neoformans. PLoS Genet. 2010, 6, e1000953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.C.; Davidson, R.C.; Cox, G.M.; Heitman, J. Pheromones stimulate mating and differentiation via paracrine and autocrine signaling in Cryptococcus neoformans. Eukaryot. Cell 2002, 1, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Feretzaki, M.; Heitman, J. Genetic Circuits that Govern Bisexual and Unisexual Reproduction in Cryptococcus neoformans. PLoS Genet. 2013, 9, e1003688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.L.; Han, L.T.; Jiang, S.T.; Chang, A.N.; Zhou, Z.Y.; Liu, T.B. The Cys2His2 zinc finger protein Zfp1 regulates sexual reproduction and virulence in Cryptococcus neoformans. Fungal Genet. Biol. 2019, 124, 59–72. [Google Scholar] [CrossRef]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci. 2006, 31, 137–155. [Google Scholar] [CrossRef]
- Liu, T.B.; Xue, C. The Ubiquitin-Proteasome System and F-box Proteins in Pathogenic Fungi. Mycobiology 2011, 39, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.B.; Wang, Y.; Stukes, S.; Chen, Q.; Casadevall, A.; Xue, C. The F-Box protein Fbp1 regulates sexual reproduction and virulence in Cryptococcus neoformans. Eukaryot. Cell 2011, 10, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Fan, C.L.; Han, L.T.; Guo, Y.B.; Liu, T.B. Role of F-box Protein Cdc4 in Fungal Virulence and Sexual Reproduction of Cryptococcus neoformans. Front. Cell. Infect. Microbiol. 2021, 11, 806465. [Google Scholar] [CrossRef]
- Brown, A.M.; Hoopes, S.L.; White, R.H.; Sarisky, C.A. Purine biosynthesis in archaea: Variations on a theme. Biol. Direct. 2011, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Wizrah, M.S.I.; Chua, S.M.H.; Luo, Z.; Manik, M.K.; Pan, M.; Whyte, J.M.L.; Robertson, A.A.B.; Kappler, U.; Kobe, B.; Fraser, J.A. AICAR transformylase/IMP cyclohydrolase (ATIC) is essential for de novo purine biosynthesis and infection by Cryptococcus neoformans. J. Biol. Chem. 2022, 298, 102453. [Google Scholar] [CrossRef] [PubMed]
- Ory, J.J.; Griffith, C.L.; Doering, T.L. An efficiently regulated promoter system for Cryptococcus neoformans utilizing the CTR4 promoter. Yeast 2004, 21, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Han, L.T.; Wu, Y.J.; Liu, T.B. The F-Box Protein Fbp1 Regulates Virulence of Cryptococcus neoformans Through the Putative Zinc-Binding Protein Zbp1. Front. Cell. Infect. Microbiol. 2021, 11, 794661. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Liu, T.; Chen, L.; Li, W.; Liu, I.; Kronstad, J.W.; Seyfang, A.; Heitman, J. Role of an expanded inositol transporter repertoire in Cryptococcus neoformans sexual reproduction and virulence. mBio 2010, 1, e00084-10. [Google Scholar] [CrossRef] [Green Version]
- Davidson, R.C.; Cruz, M.C.; Sia, R.A.; Allen, B.; Alspaugh, J.A.; Heitman, J. Gene disruption by biolistic transformation in serotype D strains of Cryptococcus neoformans. Fungal Genet. Biol. 2000, 29, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Osman, A. Yeast two-hybrid assay for studying protein-protein interactions. Methods Mol. Biol. 2004, 270, 403–422. [Google Scholar] [CrossRef]
- Liu, T.B.; Xue, C. Fbp1-mediated ubiquitin-proteasome pathway controls Cryptococcus neoformans virulence by regulating fungal intracellular growth in macrophages. Infect. Immun. 2014, 82, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, O.; Casadevall, A. Experimental modulation of capsule size in Cryptococcus neoformans. Biol. Proced. Online 2004, 6, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, A.J.; Frases, S.; Cordero, R.J.; Nimrichter, L.; Casadevall, A.; Nosanchuk, J.D. Cryptococcus neoformans responds to mannitol by increasing capsule size in vitro and in vivo. Cell Microbiol. 2010, 12, 740–753. [Google Scholar] [CrossRef] [Green Version]
- Tibbetts, A.S.; Appling, D.R. Characterization of two 5-aminoimidazole-4-carboxamide ribonucleotide transformylase/inosine monophosphate cyclohydrolase isozymes from Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 20920–20927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campuzano, A.; Wormley, F.L. Innate Immunity against Cryptococcus, from Recognition to Elimination. J. Fungi 2018, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Pareek, V.; Pedley, A.M.; Benkovic, S.J. Human de novo purine biosynthesis. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 1–16. [Google Scholar] [CrossRef]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohnmeyer, H.; Elling, L. Enzyme cascades for the synthesis of nucleotide sugars: Updates to recent production strategies. Carbohydr. Res. 2023, 523, 108727. [Google Scholar] [CrossRef]
- Serezani, C.H.; Ballinger, M.N.; Aronoff, D.M.; Peters-Golden, M. Cyclic AMP: Master regulator of innate immune cell function. Am. J. Respir. Cell. Mol. Biol. 2008, 39, 127–132. [Google Scholar] [CrossRef]
- Pukkila-Worley, R.; Gerrald, Q.D.; Kraus, P.R.; Boily, M.J.; Davis, M.J.; Giles, S.S.; Cox, G.M.; Heitman, J.; Alspaugh, J.A. Transcriptional network of multiple capsule and melanin genes governed by the Cryptococcus neoformans cyclic AMP cascade. Eukaryot. Cell 2005, 4, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Coelho, M.A.; David-Palma, M.; Priest, S.J.; Heitman, J. The Evolution of Sexual Reproduction and the Mating-Type Locus: Links to Pathogenesis of Cryptococcus Human Pathogenic Fungi. Annu. Rev. Genet. 2019, 53, 417–444. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Carter, D.A.; Dyer, P.S.; Soll, D.R. Sexual reproduction of human fungal pathogens. Cold Spring Harb. Perspect. Med. 2014, 4, a019281. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Feretzaki, M.; Li, W.; Floyd-Averette, A.; Mieczkowski, P.; Dietrich, F.S.; Heitman, J. Unisexual and heterosexual meiotic reproduction generate aneuploidy and phenotypic diversity de novo in the yeast Cryptococcus neoformans. PLoS Biol. 2013, 11, e1001653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibbetts, A.S.; Appling, D.R. Saccharomyces cerevisiae expresses two genes encoding isozymes of 5-aminoimidazole-4-carboxamide ribonucleotide transformylase. Arch. Biochem. Biophys. 1997, 340, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Rebora, K.; Laloo, B.; Daignan-Fornier, B. Revisiting purine-histidine cross-pathway regulation in Saccharomyces cerevisiae: A central role for a small molecule. Genetics 2005, 170, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, W.B.; Appling, D.R. Cloning and characterization of methenyltetrahydrofolate synthetase from Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 20205–20213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briza, P.; Bogengruber, E.; Thur, A.; Rutzler, M.; Munsterkotter, M.; Dawes, I.W.; Breitenbach, M. Systematic analysis of sporulation phenotypes in 624 non-lethal homozygous deletion strains of Saccharomyces cerevisiae. Yeast 2002, 19, 403–422. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Average fbp1Δ/H99 | No. of PEST Domain |
|---|---|---|---|
| CNAG_05514 | Uncharacterized protein | 1.899038 | 0 |
| CNAG_01974 | Ribosomal protein | 1.631707 | 2 |
| CNAG_06195 | Uncharacterized protein | 1.595274 | 0 |
| CNAG_00700 | ATIC Ade16 | 1.544354 | 0 |
| CNAG_05498 | Uncharacterized protein | 1.541769 | 1 |
| CNAG_02860 | Endo-1,3(4)-beta-glucanase | 1.522101 | 2 |
| CNAG_01019 | Superoxide dismutase | 1.470508 | 0 |
| CNAG_02344 | Uracil phosphoribosyltransferase | 1.431079 | 1 |
| CNAG_02738 | Uncharacterized protein | 1.400934 | 0 |
| CNAG_01109 | Uncharacterized protein | 1.384226 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, L.; Wu, Y.; Xiong, S.; Liu, T. Ubiquitin Degradation of the AICAR Transformylase/IMP Cyclohydrolase Ade16 Regulates the Sexual Reproduction of Cryptococcus neoformans. J. Fungi 2023, 9, 699. https://doi.org/10.3390/jof9070699
Han L, Wu Y, Xiong S, Liu T. Ubiquitin Degradation of the AICAR Transformylase/IMP Cyclohydrolase Ade16 Regulates the Sexual Reproduction of Cryptococcus neoformans. Journal of Fungi. 2023; 9(7):699. https://doi.org/10.3390/jof9070699
Chicago/Turabian StyleHan, Liantao, Yujuan Wu, Sichu Xiong, and Tongbao Liu. 2023. "Ubiquitin Degradation of the AICAR Transformylase/IMP Cyclohydrolase Ade16 Regulates the Sexual Reproduction of Cryptococcus neoformans" Journal of Fungi 9, no. 7: 699. https://doi.org/10.3390/jof9070699
APA StyleHan, L., Wu, Y., Xiong, S., & Liu, T. (2023). Ubiquitin Degradation of the AICAR Transformylase/IMP Cyclohydrolase Ade16 Regulates the Sexual Reproduction of Cryptococcus neoformans. Journal of Fungi, 9(7), 699. https://doi.org/10.3390/jof9070699