
Improvement of DOPA-Melanin Production by Aspergillus nidulans Using Eco-Friendly and Inexpensive Substrates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Corn Steep Liquor, Vinasse, and Sugarcane Molasses
2.3. Fungal Cultivation
2.4. Optimization of MEL1 Mutant Culture Conditions for Melanin Production
2.5. Melanin Extraction
2.6. Melanin Purification and Quantification
2.7. Statistical Analysis
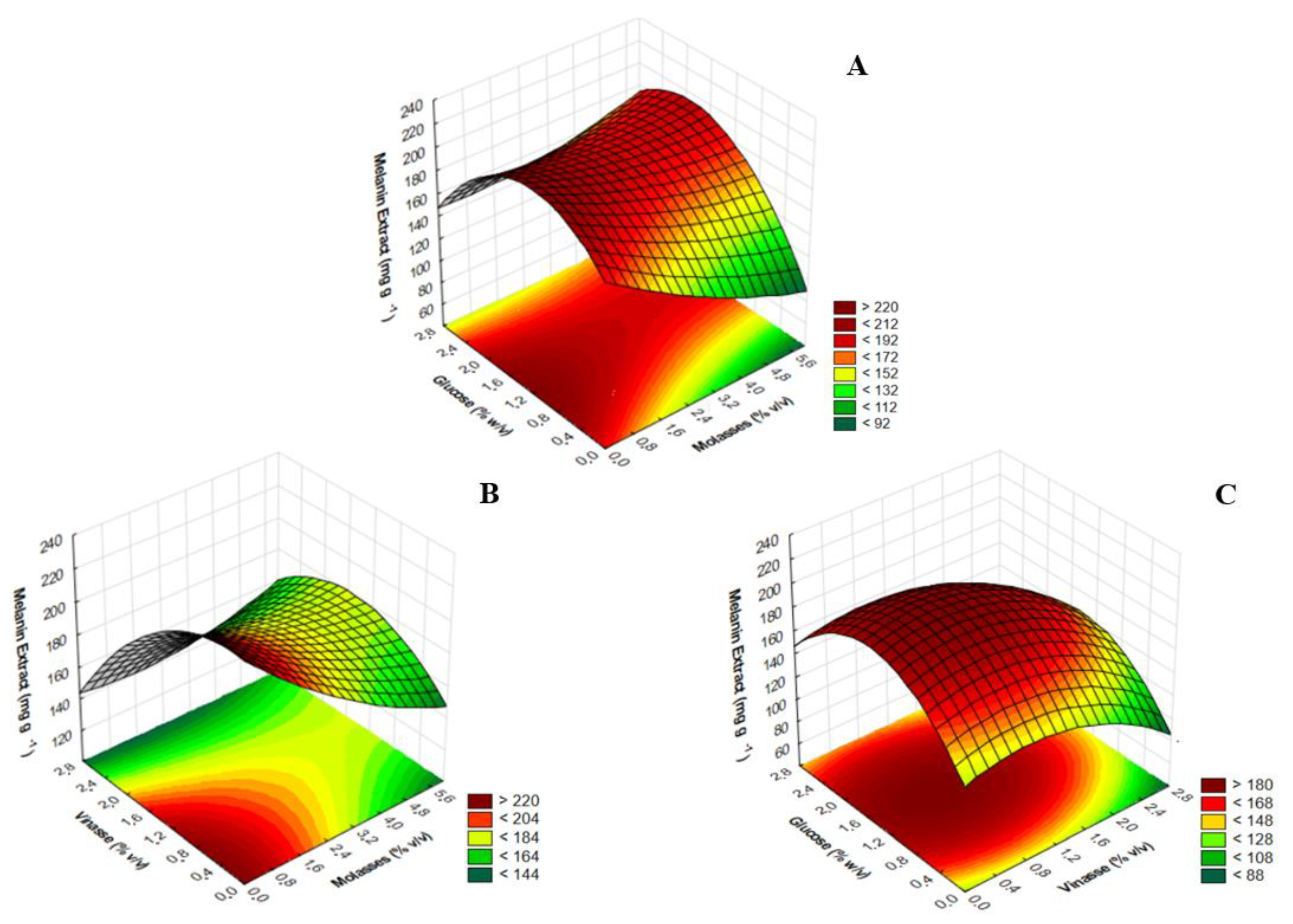
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Narsing Rao, M.P.; Xiao, M.; Li, W.J. Fungal and bacterial pigments: Secondary metabolites with wide applications. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Lyu, Y.; Yu, H.; Chen, W.; Ye, L.; Yang, R. Biotechnological advances for improving natural pigment production: A state-of-the-art review. Bioresour. Bioprocess. 2022, 9, 8. [Google Scholar] [CrossRef]
- Dufosse, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Panesar, R.; Kaur, S.; Panesar, P.S. Production of microbial pigments utilizing agro-industrial waste: A review. Curr. Opin. Food Sci. 2015, 1, 70–76. [Google Scholar] [CrossRef]
- Tuli, H.S.; Chaudhary, P.; Beniwal, V.; Sharma, A.K. Microbial pigments as natural color sources: Current trends and future perspectives. J. Food Sci. Technol. 2015, 52, 4669–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatragadda, R.; Dufossé, L. Ecological and biotechnological aspects of pigmented microbes: A way forward in development of food and pharmaceutical grade pigments. Microorganisms 2021, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Tran-Ly, A.N.; Reyes, C.; Schwarze, F.W.; Ribera, J. Microbial production of melanin and its various applications. World J. Microbiol. Biotechnol. 2020, 36, 170. [Google Scholar] [CrossRef] [PubMed]
- Mattoon, E.R.; Cordero, R.J.; Casadevall, A. Fungal Melanins and Applications in Healthcare, Bioremediation and Industry. J. Fungi 2021, 7, 488. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Nimse, S.B.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; Ghadge, V.A.; Kumar, P.; Shinde, P.B. Microbial melanin: Recent advances in biosynthesis, extraction, characterization, and applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar] [CrossRef]
- Toledo, A.V.; Franco, M.E.E.; Lopez, S.M.Y.; Troncozo, M.I.; Saparrat, M.C.N.; Balatti, P.A. Melanins in fungi: Types, localization and putative biological roles. Physiol. Mol. Plant Pathol. 2017, 99, 2–6. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; Saber, W.I. Natural Melanin: Current Trends, and Future Approaches, with Especial Reference to Microbial Source. Polymers 2022, 14, 1339. [Google Scholar] [CrossRef]
- Ghattavi, K.; Homaei, A.; Kamrani, E.; Kim, S.K. Melanin pigment derived from marine organisms and its industrial applications. Dye. Pigment. 2022, 201, 110214. [Google Scholar] [CrossRef]
- Caldas, M.; Santos, A.C.; Veiga, F.; Rebelo, R.; Reis, R.L.; Correlo, V.M. Melanin nanoparticles as a promising tool for biomedical applications—A review. Acta Biomater. 2020, 105, 26–43. [Google Scholar] [CrossRef] [PubMed]
- Pombeiro-Sponchiado, S.R.; Sousa, G.S.; Andrade, J.C.R.; Lisboa, H.F.; Goncalves, R.C.R. Production of Melanin Pigment by Fungi and its Biotechnological Applications. In Melanin; Blumenberg, M., Ed.; IntechOpen: London, UK, 2017; pp. 47–75. [Google Scholar]
- Solano, F. Melanin and melanin-related polymers as materials with biomedical and biotechnological applications—Cuttlefish ink and mussel foot proteins as inspired biomolecules. Int. J. Mol. Sci. 2017, 18, 1561. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Rhim, J.W. New insight into melanin for food packaging and biotechnology applications. Crit. Rev. Food Sci. Nutr. 2022, 62, 4629–4655. [Google Scholar] [CrossRef] [PubMed]
- El Obeid, A.S.; Kamal-Eldin, A.; Abdelhalim, M.A.K.; Haseeb, A.M. Pharmacological Properties of Melanin and its Function in Health. Basic Clin. Pharmacol. Toxicol. 2017, 120, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.A.; Wheeler, M.H. Biosynthesis and Functions of Fungal Melanins. Annu. Rev. Phytopathol. 1986, 24, 411–451. [Google Scholar] [CrossRef]
- Ellis, D.H.; Griffiths, D.A. The location and analysis of melanins in the cell walls of some soil fungi. Can. J. Microbiol. 1974, 20, 1379–1386. [Google Scholar] [CrossRef]
- Gómez, B.L.; Nosanchuk, J.D. Melanin and fungi. Curr. Opin. Infect. Dis. 2003, 16, 91–96. [Google Scholar] [CrossRef]
- Belozerskaya, T.A.; Gessler, N.N.; Aver‘yanov, A.A. Melanin Pigments of Fungi. In Fungal Metabolites; Mérillon, J.M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 263–291. [Google Scholar] [CrossRef]
- Gonçalves, R.C.R.; Lisboa, H.C.F.; Pombeiro-Sponchiado, S.R. Characterization of melanin pigment produced by Aspergillus nidulans. World J. Microbiol. Biotechnol. 2012, 28, 1467–1474. [Google Scholar] [CrossRef]
- Gonçalves, R.C.R.; Pombeiro-Sponchiado, S.R. Antioxidant Activity of the Melanin Pigment Extracted from Aspergillus nidulans. Biol. Pharm. Bull. 2005, 28, 1129–1131. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, R.D.C.R.; Kitagawa, R.R.; Raddi, M.S.G.; Carlos, I.Z.; Pombeiro-Sponchiado, S.R. Inhibition of nitric oxide and tumour necrosis factor-α production in peritoneal macrophages by Aspergillus nidulans melanin. Biol. Pharm. Bull. 2013, 36, 1915–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, R.D.C.R.; Kitagawa, R.R.; Varanda, E.A.; Raddi, M.S.G.; Leite, C.A.; Pombeiro-Sponchiado, S.R. Effect of biotransformation by liver S9 enzymes on the mutagenicity and cytotoxicity of melanin extracted from Aspergillus nidulans. Pharm. Biol. 2015, 54, 1014–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amini, Z.; Self, R.; Strong, J.; Speight, R.; O’Hara, I.; Harrison, M.D. Valorization of sugarcane biorefinery residues using fungal biocatalysis. Biomass Convers. Biorefinery 2021, 12, 997–1011. [Google Scholar] [CrossRef]
- Freitas, L.C.; Barbosa, J.R.; da Costa, A.L.C.; Bezerra, F.W.F.; Pinto, R.H.H.; de Carvalho Junior, R.N. From waste to sustainable industry: How can agro-industrial wastes help in the development of new products? Resour. Conserv. Recycl. 2021, 169, 105466. [Google Scholar] [CrossRef]
- Srianta, I.; Kusdiyantini, E.; Zubaidah, E.; Ristiarini, S.; Nugerahani, I.; Alvin, A.; Iswanto, N. Utilization of agro-industrial by-products in Monascus fermentation: A review. Bioresour. Bioprocess. 2021, 8, 129. [Google Scholar] [CrossRef]
- Ramesh, C.; Prasastha, V.R.; Venkatachalam, M.; Dufossé, L. Natural Substrates and Culture Conditions to Produce Pigments from Potential Microbes in Submerged Fermentation. Fermentation 2022, 8, 460. [Google Scholar] [CrossRef]
- Da Silva, V.L.; Ienczak, J.L.; Moritz, D. Agro-industrial residues for the production of red biopigment by Monascus ruber: Rice flour and sugarcane molasses. Braz. J. Microbiol. 2021, 52, 587–596. [Google Scholar] [CrossRef]
- Sodhi, A.S.; Sharma, N.; Bhatia, S.; Verma, A.; Soni, S.; Batra, N. Insights on sustainable approaches for production and applications of value added products. Chemosphere 2022, 286, 131623. [Google Scholar] [CrossRef]
- Lopes, F.C.; Ligabue-Braun, R. Agro-Industrial Residues: Eco-Friendly and Inexpensive Substrates for Microbial Pigments Production. Front. Sustain. Food Syst. 2021, 5, 589414. [Google Scholar] [CrossRef]
- Molelekoa, T.B.J.; Regnier, T.; da Silva, L.S.; Augustyn, W. Production of pigments by filamentous fungi cultured on agro-industrial by-products using submerged and solid-state fermentation methods. Fermentation 2021, 7, 295. [Google Scholar] [CrossRef]
- Zou, Y.; Tian, M. Fermentative Production of Melanin by Auricularia auricula. J. Food Process. Preserv. 2017, 41, e12909. [Google Scholar] [CrossRef]
- Vasanthabharathi, V.; Lakshminarayanan, R.; Jayalakshmi, S. Melanin production from marine Streptomyces. African J. Biotechnol. 2011, 10, 11224–11234. [Google Scholar] [CrossRef] [Green Version]
- Bezirhan Arikan, E.; Canli, O.; Caro, Y.; Dufossé, L.; Dizge, N. Production of bio-based pigments from food processing industry by-products (apple, pomegranate, black carrot, red beet pulps) using Aspergillus carbonarius. J. Fungi 2020, 6, 240. [Google Scholar] [CrossRef] [PubMed]
- Rani, M.H.S.; Ramesh, T.; Subramanian, J.; Kalaiselvam, M. Production and Characterization of Melanin Pigment from Halophilic Black Yeast Hortaea werneckii. Int. J. Pharma Res. Rev. 2013, 2, 9–17. Available online: https://www.ijpr.in/Data/Archives/2013/August/0406201301.pdf (accessed on 14 March 2023).
- Pretti, T.S. Otimização Das Condições De Cultivo Do Fungo Aspergillus nidulans Para Produção De Melanina Usando Resíduos Agroindustriais. Mater’s Thesis, Sao Paulo State University, São Paulo, Brazil, 2009. [Google Scholar]
- Freitas, M.G.M.D. Estudo Comparativo da Biossorção Dos Metais Terras-Raras (Nd+3, La+3 E Ce+3) Pela Biomassa Melanizada E Pigmento, Na Forma Livre E Imobilizada, Obtidos Do Fungo Aspergillus nidulans. Mater’s Thesis, Sao Paulo State University, São Paulo, Brazil, 2013. [Google Scholar]
- De Sousa, G.S. Otimização da Produção De Melanina Pelo Mutante MEL1 Do Fungo Aspergillus nidulans Usando Planejamento Fatorial. Mater’s Thesis, Sao Paulo State University, São Paulo, Brazil, 2017. [Google Scholar]
- Cove, D.J. The induction and repression of nitrate reductase in the fungus Aspergillus nidulans. Biochim. Biophys. Acta—Enzymol. Biol. Oxid. 1966, 113, 51–56. [Google Scholar] [CrossRef]
- Sponchiado, S.R.P.; De Sousa, G.S.; Lisboa, H.C.F.; Gonçalves, R.d.C.R. Processo de Produção do Pigmento Melanina Pelo Fungo Aspergillus nidulans. Brazil Patent BR1020180079522A2, 19 April 2018. [Google Scholar]
- Sava, V.M.; Galkin, B.N.; Hong, M.Y.; Yang, P.C.; Huang, G.S. A novel melanin-like pigment derived from black tea leaves with immuno-stimulating activity. Food Res. Int. 2001, 34, 337–343. [Google Scholar] [CrossRef]
- Bull, A.T. Inhibition of polysaccharases by melanin: Enzyme inhibition in relation to mycolysis. Arch. Biochem. Biophys. 1970, 137, 345–356. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, X.; Sun, S.; Zhang, L.; Shan, S.; Zhu, H. Production of natural melanin by Auricularia auricula and study on its molecular structure. Food Chem. 2016, 190, 801–807. [Google Scholar] [CrossRef]
- Galhaup, C.; Haltrich, D. Enhanced formation of laccase activity by the white-rot fungus Trametes pubescens in the presence of copper. Appl. Microbiol. Biotechnol. 2001, 56, 225–232. [Google Scholar] [CrossRef]
- Hoarau, J.; Caro, Y.; Grondin, I.; Petit, T. Sugarcane vinasse processing: Toward a status shift from waste to valuable resource. A review. J. Water Process Eng. 2018, 24, 11–25. [Google Scholar] [CrossRef]
- Espana-Gamboa, E.; Mijangos-Cortes, J.; Barahona-Perez, L.; Dominguez-Maldonado, J.; Hernández-Zarate, G.; Alzate-Gaviria, L. Vinasses: Characterization and treatments. Waste Manag. Res. 2011, 29, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, W.B.D. Estudo do efeito de indutores na melanização do mutante MEL1 do fungo Aspergillus nidulans. Microb. Cell Fact. 2020, 21, 278. [Google Scholar] [CrossRef] [PubMed]
- Mujdeci, G.N. Natural melanin synthesized by Aureobasidium pullulans using food wastes and its characterization. Appl. Food Biotechnol. 2021, 8, 307–318. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Levels | ||||
|---|---|---|---|---|
| Variables | Coded Value | −1 | 0 | +1 |
| Corn steep liquor (% v/v) | X1 | 0 | 0.50 | 1.00 |
| Molasses (% v/v) | X2 | 0 | 1.00 | 2.00 |
| Vinasse (% v/v) | X3 | 0 | 0.50 | 1.00 |
| Sodium nitrate (% w/v) | X4 | 0 | 0.30 | 0.60 |
| Glucose (% w/v) | X5 | 0 | 0.50 | 1.00 |
| Levels | ||||||
|---|---|---|---|---|---|---|
| Variables | Coded Value | −1.68 | −1 | 0 | +1 | +1.68 |
| Molasses (% v/v) | X1 | 0.01 | 1.00 | 2.45 | 3.90 | 4.89 |
| Vinasse (% v/v) | X2 | 0.01 | 0.50 | 1.23 | 1.95 | 2.44 |
| Glucose (% w/v) | X3 | 0.01 | 0.50 | 1.23 | 1.95 | 2.44 |
| Run | Corn Steep Liquor (v/v) | Molasses (v/v) | Vinasse (v/v) | Sodium Nitrate (w/v) | Glucose (w/v) | Melanin Extract (a) (mg g−1 of Biomass) |
|---|---|---|---|---|---|---|
| 1 | −1 (0%) | −1 (0%) | −1 (0%) | −1 (0%) | 1 (1%) | 173.26 |
| 2 | 1 (1%) | −1 (0%) | −1 (0%) | −1 (0%) | −1 (0%) | 22.86 |
| 3 | −1 (0%) | 1 (2%) | −1 (0%) | −1 (0%) | −1 (0%) | 133.33 |
| 4 | 1 (1%) | 1 (2%) | −1 (0%) | −1 (0%) | 1 (1%) | 151.04 |
| 5 | −1 (0%) | −1 (0%) | 1 (1%) | −1 (0%) | −1 (0%) | 32.97 |
| 6 | 1 (1%) | −1 (0%) | 1 (1%) | −1 (0%) | 1 (1%) | 46.46 |
| 7 | −1 (0%) | 1 (2%) | 1 (1%) | −1 (0%) | 1 (1%) | 192.64 |
| 8 | 1 (1%) | 1 (2%) | 1 (1%) | −1 (0%) | −1 (0%) | 74.98 |
| 9 | −1 (0%) | −1 (0%) | −1 (0%) | 1 (0.6%) | −1 (0%) | 25.97 |
| 10 | 1 (1%) | −1 (0%) | −1 (0%) | 1 (0.6%) | 1 (1%) | 53.43 |
| 11 | −1 (0%) | 1 (2%) | −1 (0%) | 1 (0.6%) | 1 (1%) | 158.21 |
| 12 | 1 (1%) | 1 (2%) | −1 (0%) | 1 (0.6%) | −1 (0%) | 57.08 |
| 13 | −1 (0%) | −1 (0%) | 1 (1%) | 1 (0.6%) | 1 (1%) | 181.22 |
| 14 | 1 (1%) | −1 (0%) | 1 (1%) | 1 (0.6%) | −1 (0%) | 12.23 |
| 15 | −1 (0%) | 1 (2%) | 1 (1%) | 1 (0.6%) | −1 (0%) | 174.39 |
| 16 | 1 (1%) | 1 (2%) | 1 (1%) | 1 (0.6%) | 1 (1%) | 99.75 |
| 17 | 0 (0.5%) | 0 (1%) | 0 (0.5%) | 0 (0.3%) | 0 (0.5%) | 96.52 |
| 18 | 0 (0.5%) | 0 (1%) | 0 (0.5%) | 0 (0.3%) | 0 (0.5%) | 94.90 |
| 19 | 0 (0.5%) | 0 (1%) | 0 (0.5%) | 0 (0.3%) | 0 (0.5%) | 90.52 |
| Factors | Coded Value | Effect | Pure Error | t (2) | p(a) |
|---|---|---|---|---|---|
| Mean | 98.51 | 0.71 | 138.34 | 0.000052 | |
| Corn steep liquor (% v/v) | X1 | −69.27 | 1.55 | −44.63 | 0.000502 |
| Molasses (% v/v) | X2 | 61.63 | 1.55 | 39.71 | 0.000634 |
| Vinasse (% v/v) | X3 | 4.93 | 1.55 | 3.18 | 0.086373 |
| Sodium Nitrate (% w/v) | X4 | −8.16 | 1.55 | −5.26 | 0.034350 |
| Glucose (% w/v) | X5 | 65.27 | 1.55 | 42.06 | 0.000565 |
| Runs | Molasses (v/v) | Vinasse (v/v) | Glucose (w/v) | Melanin Extract (a) (mg g−1 of Biomass) |
|---|---|---|---|---|
| 1 | −1 (1.00%) | −1 (0.50%) | −1 (0.50%) | 197.92 |
| 2 | 1 (3.90%) | −1 (0.50%) | −1 (0.50%) | 129.85 |
| 3 | −1 (1.00%) | 1 (1.95%) | −1 (0.50%) | 164.59 |
| 4 | 1 (3.90%) | 1 (1.95%) | −1 (0.50%) | 135.58 |
| 5 | −1 (1.00%) | −1 (0.50%) | 1 (1.95%) | 198.81 |
| 6 | 1 (3.90%) | −1 (0.50%) | 1 (1.95%) | 174.80 |
| 7 | −1 (1.00%) | 1(1.95%) | 1(1.95%) | 177.29 |
| 8 | 1 (3.90%) | 1 (1.95%) | 1 (1.95%) | 165.11 |
| 9 * | −1.68 (0.00%) | 0 (1.23%) | 0 (1.23%) | 218.21 |
| 10 * | 1.68 (4.89%) | 0 (1.23%) | 0 (1.23%) | 203.56 |
| 11 * | 0 (2.45%) | −1.68 (0.00%) | 0 (1.23%) | 202.85 |
| 12 * | 0 (2.45%) | 1.68 (2.40%) | 0 (1.23%) | 166.52 |
| 13 * | 0 (2.45%) | 0 (1.23%) | −1.68 (0.00%) | 148.83 |
| 14 * | 0 (2.45%) | 0 (1.23%) | 1.68 (2.44%) | 180.14 |
| 15 ** | 0 (2.45%) | 0 (1.23%) | 0 (1.23%) | 189.74 |
| 16 ** | 0 (2.45%) | 0 (1.23%) | 0 (1.23%) | 191.29 |
| 17 ** | 0 (2.45%) | 0 (1.23%) | 0 (1.23%) | 199.00 |
| Variation Factor | Sum of Squares | Degrees of Freedom | Medium Square | F Calc. | F Crit. | p-Value |
|---|---|---|---|---|---|---|
| Regression | 8157.928 | 8 | 1019.741 | 4.639118 | 3.44 | 0.021948 * |
| Residual | 1758.508 | 8 | 219.8136 | |||
| Lack of fit | 1709.302 | 6 | 284.8836 | 11.57904 | 19.33 | 0.081619 |
| Pure error | 49.20678 | 2 | 24.60339 | |||
| Total | 9916.437 | 16 |
| Conditions | Biomass (g L−1) | Purified Melanin (mg g−1 of Biomass) |
|---|---|---|
| Before optimization | 3.92 ± 0.06 | 100.32 ± 28.13 |
| After optimization | 4.57 ± 0.16 | 225.39 ± 4.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campanhol, B.S.; Ribeiro, B.D.; Casellato, F.; Medina, K.J.D.; Sponchiado, S.R.P. Improvement of DOPA-Melanin Production by Aspergillus nidulans Using Eco-Friendly and Inexpensive Substrates. J. Fungi 2023, 9, 714. https://doi.org/10.3390/jof9070714
Campanhol BS, Ribeiro BD, Casellato F, Medina KJD, Sponchiado SRP. Improvement of DOPA-Melanin Production by Aspergillus nidulans Using Eco-Friendly and Inexpensive Substrates. Journal of Fungi. 2023; 9(7):714. https://doi.org/10.3390/jof9070714
Chicago/Turabian StyleCampanhol, Beatriz Silva, Beatriz Dias Ribeiro, Fernando Casellato, Kelly Johana Dussán Medina, and Sandra Regina Pombeiro Sponchiado. 2023. "Improvement of DOPA-Melanin Production by Aspergillus nidulans Using Eco-Friendly and Inexpensive Substrates" Journal of Fungi 9, no. 7: 714. https://doi.org/10.3390/jof9070714
APA StyleCampanhol, B. S., Ribeiro, B. D., Casellato, F., Medina, K. J. D., & Sponchiado, S. R. P. (2023). Improvement of DOPA-Melanin Production by Aspergillus nidulans Using Eco-Friendly and Inexpensive Substrates. Journal of Fungi, 9(7), 714. https://doi.org/10.3390/jof9070714