

Suppression of Chitin-Triggered Immunity by Plant Fungal Pathogens: A Case Study of the Cucurbit Powdery Mildew Fungus Podosphaera xanthii

 , , and
, , and
Abstract
:1. Introduction
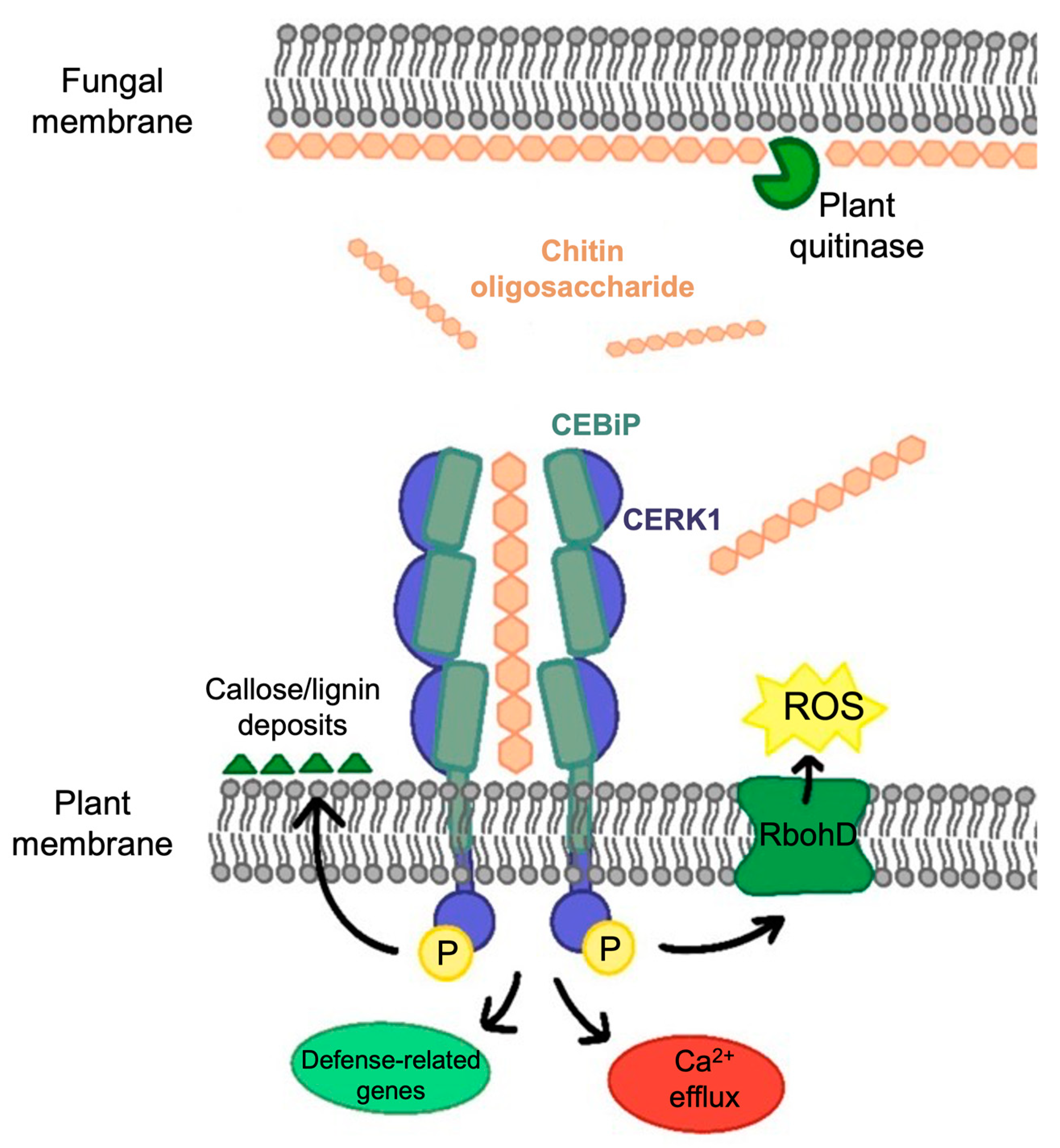
2. Chitin-Triggered Plant Immunity
3. Fungal Strategies to Overcome Chitin-Triggered Immunity
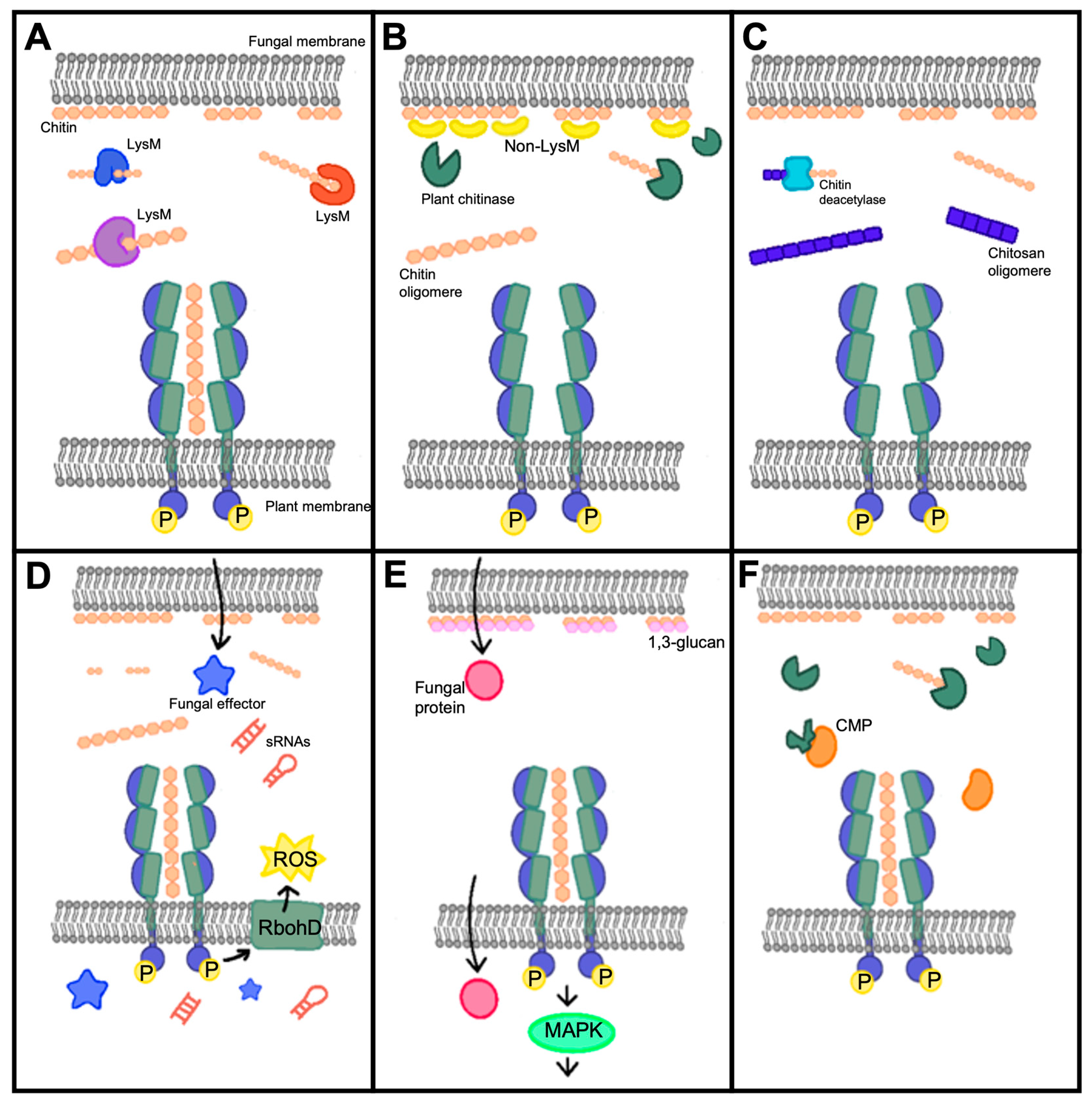
3.1. LysM and Non-LysM Containing Effectors
3.2. Chitin or Polysaccharide Deacetylases
3.3. Cytoplasmic Effectors
3.4. Chitinases and Other Mechanisms
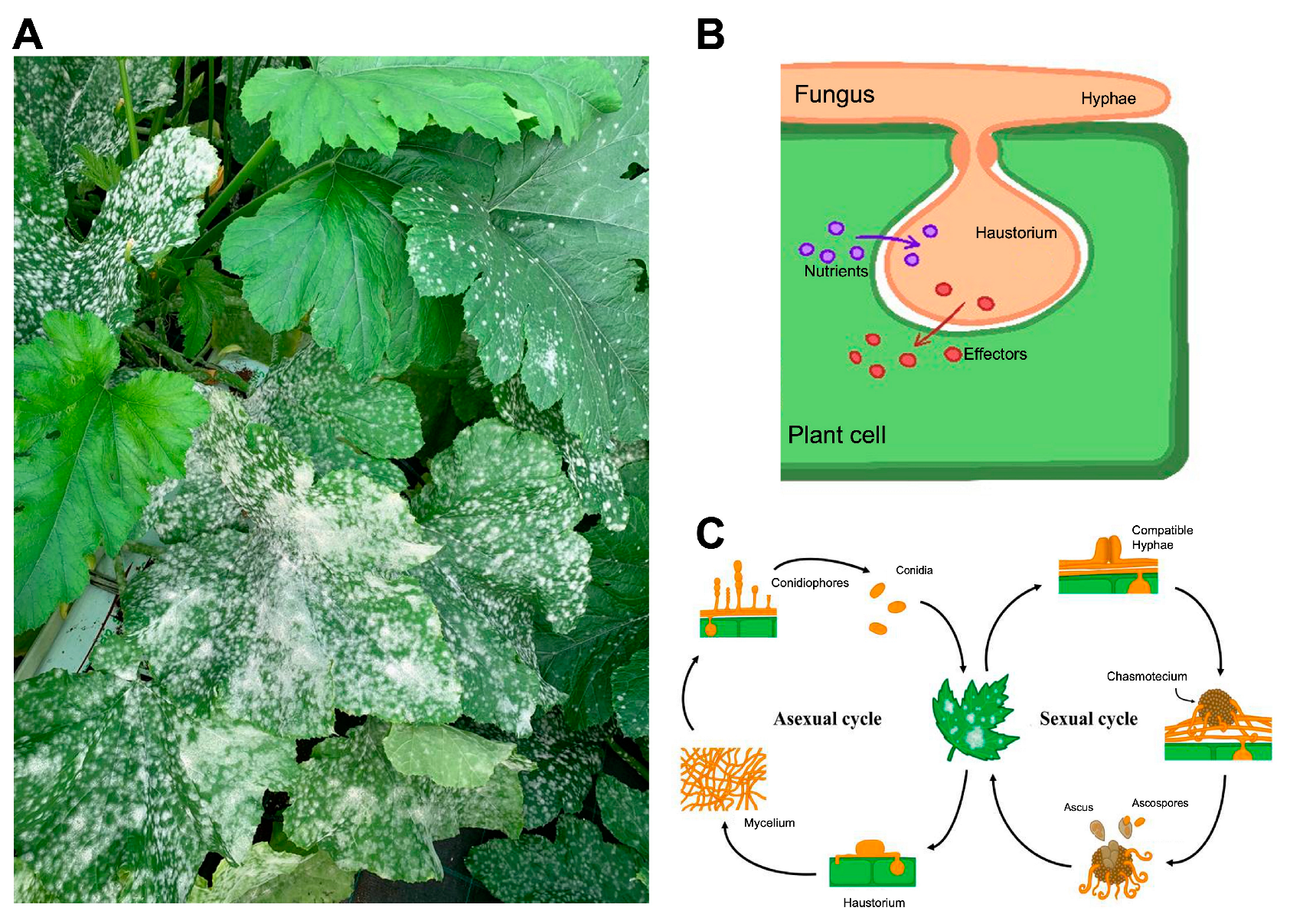
4. The Powdery Mildew Fungi and the Cucurbit Pathogen Podosphaera xanthii as a Model Species
5. Countermeasures of P. xanthii for Subverting Chitin Signaling
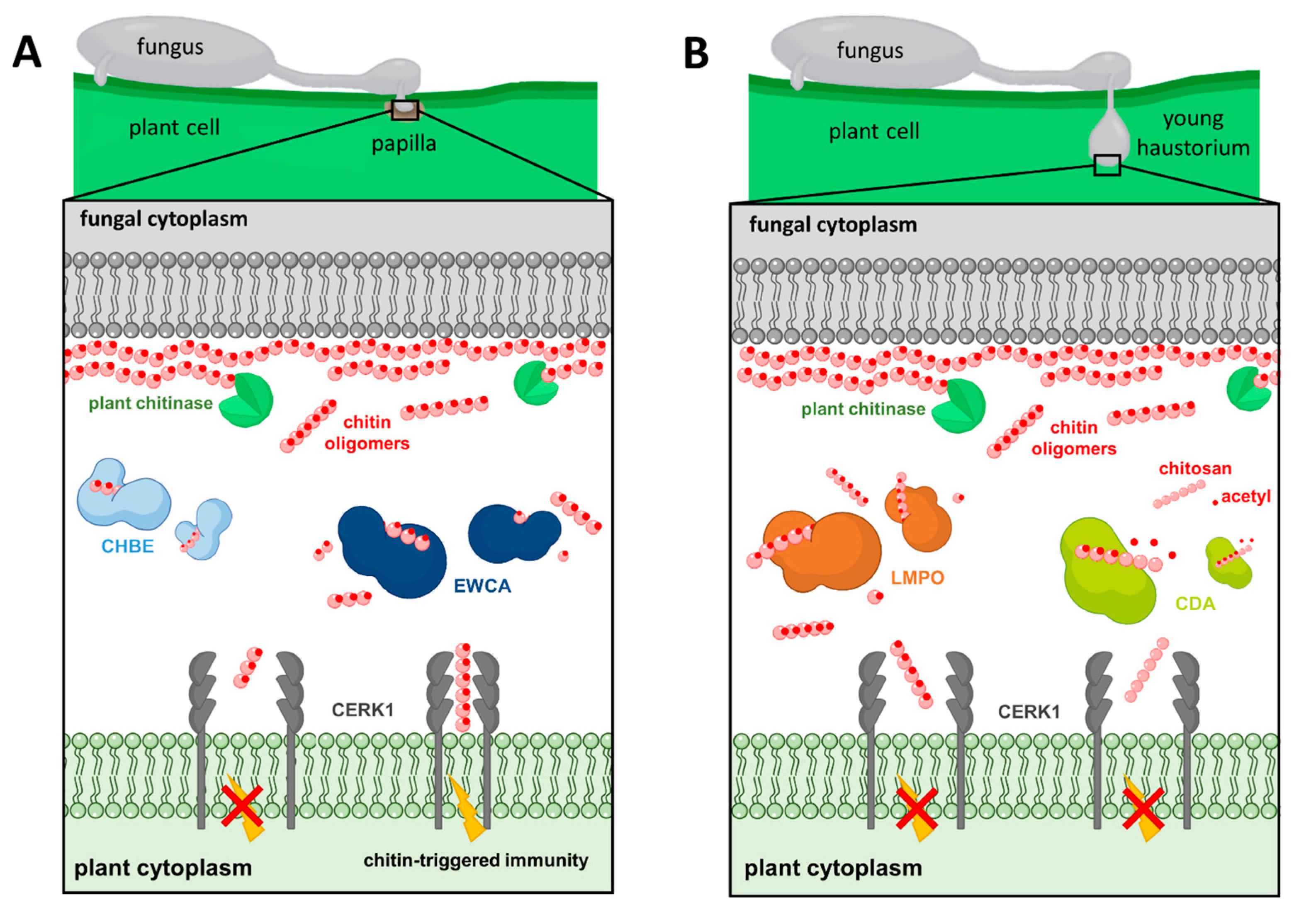
5.1. Degradation of Chitin Oligomers
5.2. Chitin Deacetylation
5.3. Binding of Chitin Oligomers
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Glawe, D.A. The powdery mildews: A review of the world’s most familiar (yet poorly known) plant pathogens. Annu. Rev. Phytopathol. 2008, 46, 27–51. [Google Scholar] [CrossRef] [PubMed]
- Kombrink, A.; Sánchez-Vallet, A.; Thomma, B.P. The role of chitin detection in plant–pathogen interactions. Microbes Infect. 2011, 13, 1168–1176. [Google Scholar] [CrossRef]
- Kamle, M.; Borah, R.; Bora, H.; Jaiswal, A.K.; Singh, R.K.; Kumar, P. Systemic acquired resistance (SAR) and induced systemic resistance (ISR): Role and mechanism of action against phytopathogens. In Fungal Biotechnology and Bioengineering; Springer: Cham, Switzerland, 2020; pp. 457–470. [Google Scholar]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, R.; Bolton, M.D.; Thomma, B.P. How filamentous pathogens co-opt plants: The ins and outs of fungal effectors. Curr. Opin. Plant Biol. 2011, 14, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Schwessinger, B.; Ronald, C. Plant innate immunity: Perception of conserved microbial signatures. Annu. Rev. Plant Biol. 2012, 63, 451–482. [Google Scholar] [CrossRef] [Green Version]
- Nürnberger, T.; Brunner, F. Innate immunity in plants and animals: Emerging parallels between the recognition of general elicitors and pathogen-associated molecular patterns. Curr. Opin. Plant Biol. 2002, 5, 318–324. [Google Scholar] [CrossRef]
- Pusztahelyi, T. Chitin and chitin-related compounds in plant–fungal interactions. Mycology 2018, 9, 189–201. [Google Scholar] [CrossRef]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Minami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Nakano, T.; Takamizawa, D.; Desaki, Y.; Ishii-Minami, N.; Nishizawa, Y.; Minami, E.; Okada, K.; Yamane, H.; Kaku, H.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 19613–19618. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vallet, A.; Mesters, J.R.; Thomma, B.P. The battle for chitin recognition in plant-microbe interactions. FEMS Microbiol. Rev. 2015, 39, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gow, N.A.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Free, S.J. Fungal cell wall organization and biosynthesis. Adv. Genet. 2013, 81, 33–82. [Google Scholar] [PubMed]
- Latgé, J.P.; Beauvais, A. Functional duality of the cell wall. Curr. Opin. Microbiol. 2014, 20, 111–117. [Google Scholar] [CrossRef]
- Cook, D.E.; Mesarich, C.H.; Thomma, B.P. Understanding plant immunity as a surveillance system to detect invasion. Annu. Rev. Phytopathol. 2015, 53, 541–563. [Google Scholar] [CrossRef]
- Thomma, B.P.; Penninckx, I.A.; Cammue, B.P.; Broekaert, W.F. The complexity of disease signaling in Arabidopsis. Curr. Opin. Immunol. 2001, 13, 63–68. [Google Scholar] [CrossRef]
- Felix, G.; Regenass, M.; Boller, T. Specific perception of subnanomolar concentrations of chitin fragments by tomato cells: Induction of extracellular alkalinization, changes in protein phosphorylation, and establishment of a refractory state. Plant J. 1993, 4, 307–316. [Google Scholar] [CrossRef]
- Kombrink, A.; Thomma, B.P. LysM effectors: Secreted proteins supporting fungal life. PLoS Pathog. 2013, 9, e1003769. [Google Scholar] [CrossRef] [Green Version]
- Schlumbaum, A.; Mauch, F.; Vögeli, U.; Boller, T. Plant chitinases are potent inhibitors of fungal growth. Nature 1986, 324, 365–367. [Google Scholar] [CrossRef]
- Grison, R.; Grezes-Besset, B.; Schneider, M.; Lucante, N.; Olsen, L.; Leguay, J.J.; Toppan, A. Field tolerance to fungal pathogens of Brassica napus constitutively expressing a chimeric chitinase gene. Nat. Biotechnol. 1996, 14, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, Y.; Kawakami, A.; Hibi, T.; He, D.Y.; Shibuya, N.; Minami, E. Regulation of the chitinase gene expression in suspension-cultured rice cells by N-acetylchitooligosaccharides: Differences in the signal transduction pathways leading to the activation of elicitor-responsive genes. Plant Mol. Biol. 1999, 39, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, D.; Robatzek, S. Pattern recognition receptors: From the cell surface to intracellular dynamics. Mol. Plant-Microbe Interact. 2007, 20, 1031–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of Microbe–associated molecular patterns and danger signals by pattern-recognition. Ann. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Doehlemann, G.; Hemetsberger, C. Apoplastic immunity and its suppression by filamentous plant pathogens. New Phytol. 2013, 198, 1001–1016. [Google Scholar] [CrossRef]
- Ito, Y.; Kaku, H.; Shibuya, N. Identification of a high-affinity binding protein for N-acetylchitooligosaccharide elicitor in the plasma membrane of suspension-cultured rice cells by affinity labeling. Plant J. 1997, 12, 347–356. [Google Scholar] [CrossRef]
- Buist, G.; Steen, A.; Kok, J.; Kuipers, O.P. LysM, a widely distributed protein motif for binding to (peptido) glycans. Mol. Microbiol. 2008, 68, 838–847. [Google Scholar] [CrossRef] [Green Version]
- Kouzai, Y.; Nakajima, K.; Hayafune, M.; Ozawa, K.; Kaku, H.; Shibuya, N.; Minami, E.; Nishizawa, Y. CEBiP is the major chitin oligomer-binding protein in rice and plays a main role in the perception of chitin oligomers. Plant Mol. Biol. 2014, 84, 519–528. [Google Scholar] [CrossRef]
- Kouzai, Y.; Kaku, H.; Shibuya, N.; Minami, E.; Nishizawa, Y. Expression of the chimeric receptor between the chitin elicitor receptor CEBiP and the receptor-like protein kinase Pi-d2 leads to enhanced responses to the chitin elicitor and disease resistance against Magnaporthe oryzae in rice. Plant Mol. Biol. 2013, 81, 287–295. [Google Scholar] [CrossRef]
- Shinya, T.; Motoyama, N.; Ikeda, A.; Wada, M.; Kamiya, K.; Hayafune, M.; Kaku, H.; Shibuya, N. Functional characterization of CEBiP and CERK1 homologs in Arabidopsis and rice reveals the presence of different chitin receptor systems in plants. Plant Cell Physiol. 2012, 53, 1696–1706. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Zhang, X.C.; Neece, D.; Ramonell, K.M.; Clough, S.; Kim, S.Y.; Stacey, M.G.; Stacey, G. A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis. Plant Cell 2008, 20, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Liu, Z.; Song, C.; Hu, Y.; Han, Z.; She, J.; Fan, F.; Wang, J.; Jin, C.; Chang, J.; et al. Chitin-induced dimerization activates a plant immune receptor. Science 2012, 336, 1160–1164. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Tanaka, K.; Zhang, X.C.; Son, G.H.; Brechenmacher, L.; Nguyen, T.H.N.; Stacey, G. LYK4, a lysin motif receptor-like kinase, is important for chitin signaling and plant innate immunity in Arabidopsis. Plant Physiol. 2012, 160, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. eLife 2014, 3, e03766. [Google Scholar] [CrossRef]
- Hayafune, M.; Berisio, R.; Marchetti, R.; Silipo, A.; Kayama, M.; Desaki, Y.; Arima, S.; Squeglia, F.; Ruggiero, A.; Tokuyasu, K.; et al. Chitin-induced activation of immune signaling by the rice receptor CEBiP relies on a unique sandwich-type dimerization. Proc. Natl. Acad. Sci. USA 2014, 111, E404–E413. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Park, C.H.; He, F.; Nagano, M.; Wang, M.O.; Bellizzi, M.; Zhang, K.; Zeng, X.; Liu, W.; Ning, Y.; et al. The RhoGAP SPIN6 associates with SPL11 and OsRac1 and negatively regulates programmed cell death and innate immunity in rice. PLoS Pathog. 2015, 11, e1004629. [Google Scholar] [CrossRef] [PubMed]
- Shinya, T.; Nakagawa, T.; Kaku, H.; Shibuya, N. Chitin-mediated plant–fungal interactions: Catching, hiding and handshaking. Curr. Opin. Plant Biol. 2015, 26, 64–71. [Google Scholar] [CrossRef]
- Naumann, T.A.; Wicklow, D.T.; Kendra, D.F. Maize seed chitinase is modified by a protein secreted by Bipolaris zeicola. Physiol. Mol. Plant Pathol. 2009, 74, 134–141. [Google Scholar] [CrossRef]
- Van den Burg, H.A.; Harrison, S.J.; Joosten, M.H.; Vervoort, J.; de Wit, J. Cladosporium fulvum Avr4 protects fungal cell walls against hydrolysis by plant chitinases accumulating during infection. Mol. Plant-Microbe Interact. 2006, 19, 1420–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, R.; Peter van Esse, H.; Kombrink, A.; Shinya, T.; Desaki, Y.; Bours, R.; Van Der Krol, S.; Shibuya, N.; Joosten, M.H.; Thomma, B.P. Conserved fungal LysM effector Ecp6 prevents chitin-triggered immunity in plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]
- Takahara, H.; Hacquard, S.; Kombrink, A.; Hughes, H.B.; Halder, V.; Robin, G.P.; Hiruma, K.; Neumann, U.; Shinya, T.; Kombrink, E.; et al. Colletotrichum higginsianum extracellular LysM proteins play dual roles in appressorial function and suppression of chitin-triggered plant immunity. New Phytol. 2016, 211, 1323–1337. [Google Scholar] [CrossRef] [Green Version]
- Irieda, H.; Inoue, Y.; Mori, M.; Yamada, K.; Oshikawa, Y.; Saitoh, H.; Uemura, A.; Terauchi, R.; Kitakura, S.; Kosaka, A.; et al. Conserved fungal effector suppresses PAMP-triggered immunity by targeting plant immune kinases. Proc. Natl. Acad. Sci. USA 2019, 116, 496–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jashni, M.K.; Dols, I.H.; Iida, Y.; Boeren, S.; Beenen, H.G.; Mehrabi, R.; Collemare, J.; de Wit, J. Synergistic action of a metalloprotease and a serine protease from Fusarium oxysporum f. sp. lycopersici cleaves chitin-binding tomato chitinases, reduces their antifungal activity, and enhances fungal virulence. Mol. Plant-Microbe Interact. 2015, 28, 996–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Zhang, B.S.; Zhao, J.H.; Huang, J.F.; Jia, S.; Wang, S.; Zhang, J.; Zhou, J.M.; Guo, H.S. Deacetylation of chitin oligomers increases virulence in soil-borne fungal pathogens. Nat. Plants 2019, 5, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Naumann, T.A.; Wicklow, D.T.; Price, N.P. Identification of a chitinase-modifying protein from Fusarium verticillioides: Truncation of a host resistance protein by a fungalysin metalloprotease. J. Biol. Chem. 2011, 286, 35358–35366. [Google Scholar] [CrossRef] [Green Version]
- Bai, P.; Park, C.H.; Shirsekar, G.; Songkumarn, P.; Bellizzi, M.; Wang, G.L. Role of lysine residues of the Magnaporthe oryzae effector AvrPiz-t in effector-and PAMP-triggered immunity. Mol. Plant Pathol. 2019, 20, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xing, J.; Cai, X.; Hendy, A.; He, W.; Yang, J.; Huang, J.; Peng, Y.L.; Ryder, L.; Chen, X.L. GPI7-mediated glycosylphosphatidylinositol anchoring regulates appressorial penetration and immune evasion during infection of Magnaporthe oryzae. Environ. Microbiol. 2020, 22, 2581–2595. [Google Scholar] [CrossRef]
- Yang, C.; Yu, Y.; Huang, J.; Meng, F.; Pang, J.; Zhao, Q.; Islam, M.A.; Xu, N.; Tian, Y.; Liu, J. Binding of the Magnaporthe oryzae chitinase MoChia1 by a rice tetratricopeptide repeat protein allows free chitin to trigger immune responses. Plant Cell 2019, 31, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P.; et al. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef] [Green Version]
- Fiorin, G.L.; Sanchéz-Vallet, A.; de Toledo Thomazella, D.P.; do Prado, F.V.; do Nascimento, L.C.; de Oliveira Figueira, A.V.; Thomma, B.P.; Pereira, G.A.G.; Teixeira, J.P.L. Suppression of plant immunity by fungal chitinase-like effectors. Curr. Biol. 2018, 28, 3023–3030. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; van den Burg, H.A.; Ökmen, B.; Beenen, H.G.; van Liere, S.; Kema, G.H.; de Wit, J. Tomato Cf resistance proteins mediate recognition of cognate homologous effectors from fungi pathogenic on dicots and monocots. Proc. Natl. Acad. Sci. USA 2010, 107, 7610–7615. [Google Scholar] [CrossRef]
- Liu, Z.; Gao, Y.; Kim, Y.M.; Faris, J.D.; Shelver, W.L.; de Wit, J.; Xu, S.S.; Friesen, T.L. SnTox1, a Parastagonospora nodorum necrotrophic effector, is a dual-function protein that facilitates infection while protecting from wheat-produced chitinases. New Phytol. 2016, 211, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Cord-Landwehr, S.; Melcher, R.L.; Kolkenbrock, S.; Moerschbacher, B.M. A chitin deacetylase from the endophytic fungus Pestalotiopsis sp. efficiently inactivates the elicitor activity of chitin oligomers in rice cells. Sci. Rep. 2016, 6, 38018. [Google Scholar] [CrossRef] [PubMed]
- Muirhead, K.; Pérez-López, E. Plasmodiophora brassicae CBM18 Proteins Bind Chitin and Suppress Chitin-Triggered Immunity. PhytoFrontiers 2022, 2, 21–29. [Google Scholar] [CrossRef]
- Martínez-Cruz, J.; Romero, D.; Hierrezuelo, J.; Thon, M.; de Vicente, A.; Pérez-García, A. Effectors with chitinase activity (EWCAs), a family of conserved, secreted fungal chitinases that suppress chitin-triggered immunity. Plant Cell 2021, 33, 1319–1340. [Google Scholar] [CrossRef]
- Polonio, Á.; Fernández-Ortuño, D.; de Vicente, A.; Pérez-García, A. A haustorial-expressed lytic polysaccharide monooxygenase from the cucurbit powdery mildew pathogen Podosphaera xanthii contributes to the suppression of chitin-triggered immunity. Mol. Plant Pathol. 2021, 22, 580–601. [Google Scholar] [CrossRef]
- Martínez-Cruz, J.M.; Polonio, Á.; Zanni, R.; Romero, D.; Gálvez, J.; Fernández-Ortuño, D.; Pérez-García, A. Chitin Deacetylase, a Novel Target for the Design of Agricultural Fungicides. J. Fungi 2021, 7, 1009. [Google Scholar] [CrossRef]
- Martínez-Cruz, J.M.; Polonio, Á.; Ruiz-Jiménez, L.; Vielba-Fernández, A.; Hierrezuelo, J.; Romero, D.; de Vicente, A.; Fernández-Ortuño, D.; Pérez-García, A. Suppression of Chitin-Triggered Immunity by a New Fungal Chitin-Binding Effector Resulting from Alternative Splicing of a Chitin Deacetylase Gene. J. Fungi 2022, 8, 1022. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, J.; Zhao, J.; Xu, J.; Sun, S.; Zhang, H.; Wu, J.; Tang, C.; Kang, Z.; Wang, X. A polysaccharide deacetylase from Puccinia striiformis f. sp. tritici is an important pathogenicity gene that suppresses plant immunity. Plant Biotechnol. J. 2020, 18, 1830–1842. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, S.; Cord-Landwehr, S.; Singh, R.; Bernard, F.; Kolkenbrock, S.; El Gueddari, N.E.; Moerschbacher, B.M. A recombinant fungal chitin deacetylase produces fully defined chitosan oligomers with novel patterns of acetylation. Appl. Environ. Microbiol. 2016, 82, 6645–6655. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Peng, X.; Wang, Y.; Hua, K.; Xing, F.; Zheng, Y.; Liu, W.; Sun, W.; Wei, S. The effector AGLIP1 in Rhizoctonia solani AG1 IA triggers cell death in plants and promotes disease development through inhibiting PAMP-triggered immunity in Arabidopsis thaliana. Front. Microbiol. 2019, 10, 2228. [Google Scholar] [CrossRef] [PubMed]
- Dölfors, F.; Holmquist, L.; Dixelius, C.; Tzelepis, G. A LysM effector protein from the basidiomycete Rhizoctonia solani contributes to virulence through suppression of chitin-triggered immunity. Mol. Genet. Genom. 2019, 294, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Rodriguez-Moreno, L.; Mansurkhodzaev, A.; Wang, P.; van den Berg, W.; Gasciolli, V.; Cottaz, S.; Fort, S.; Thomma, B.P.; Bono, J.J.; et al. A lysin motif effector subverts chitin-triggered immunity to facilitate arbuscular mycorrhizal symbiosis. New Phytol. 2020, 225, 448–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Contreras, Y.J.; Ramírez-Valdespino, C.A.; Guzmán-Guzmán, P.; Macías-Segoviano, J.I.; Villagómez-Castro, J.C.; Olmedo-Monfil, V. Tal6 from Trichoderma atroviride is a LysM effector involved in mycoparasitism and plant association. Front. Microbiol. 2019, 10, 2231. [Google Scholar] [CrossRef]
- 67. Fang, A.; Gao, H.; Zhang, N.; Zheng, X.; Qiu, S.; Li, Y.; Zhou, S.; Cui, F.; Sun, W. A novel effector gene SCRE2 contributes to full virulence of Ustilaginoidea virens to rice. Front. Microbiol. 2019, 10, 845. [Google Scholar] [CrossRef]
- Ökmen, B.; Kemmerich, B.; Hilbig, D.; Wemhöner, R.; Aschenbroich, J.; Perrar, A.; Huesgen, F.; Schipper, K.; Doehlemann, G. Dual function of a secreted fungalysin metalloprotease in Ustilago maydis. New Phytol. 2018, 220, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Han, L.B.; Li, Y.B.; Wang, F.X.; Wang, W.Y.; Liu, J.; Wu, J.H.; Zhong, N.Q.; Wu, S.J.; Jiao, G.L.; Wang, H.Y.; et al. The cotton apoplastic protein CRR1 stabilizes chitinase 28 to facilitate defense against the fungal pathogen Verticillium dahliae. Plant Cell 2019, 31, 520–536. [Google Scholar] [CrossRef]
- Kombrink, A.; Rovenich, H.; Shi-Kunne, X.; Rojas-Padilla, E.; van den Berg, G.C.; Domazakis, E.; De Jonge, R.; Valkenburg, D.J.; Sánchez-Vallet, A.; Seidl, M.F.; et al. Verticillium dahliae LysM effectors differentially contribute to virulence on plant hosts. Mol. Plant Pathol. 2017, 18, 596–608. [Google Scholar] [CrossRef] [Green Version]
- Volk, H.; Marton, K.; Flajšman, M.; Radišek, S.; Tian, H.; Hein, I.; Podlipnik, Č.; Thomma, B.P.; Košmelj, K.; Javornik, B.; et al. Chitin-binding protein of Verticillium nonalfalfae disguises fungus from plant chitinases and suppresses chitin-triggered host immunity. Mol. Plant Microbe Interact. 2019, 32, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.; Kombrink, A.; Motteram, J.; Loza-Reyes, E.; Lucas, J.; Hammond-Kosack, K.E.; Thomma, B.P.; Rudd, J.J. Analysis of two in planta expressed LysM effector homologs from the fungus Mycosphaerella graminicola reveals novel functional properties and varying contributions to virulence on wheat. Plant Physiol. 2011, 156, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Song, L.; Peng, C.; Liu, X.; Liu, L.; Zhang, Y.; Wang, W.; Zhou, J.; Wang, S.; Ebbole, D.; et al. A Magnaporthe chitinase interacts with a rice jacalin-related lectin to promote host colonization. Plant Physiol. 2019, 179, 1416–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.L.; Shi, T.; Yang, J.; Shi, W.; Gao, X.; Chen, D.; Xu, X.; Xu, J.R.; Talbot, N.J.; Peng, Y.L. N-glycosylation of effector proteins by an α-1, 3-mannosyltransferase is required for the rice blast fungus to evade host innate immunity. Plant Cell 2014, 26, 1360–1376. [Google Scholar] [CrossRef] [Green Version]
- Jian, J.; Liang, X. One small RNA of Fusarium graminearum targets and silences CEBiP gene in common wheat. Microorganisms 2019, 7, 425. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, T.; Sakaguchi, A.; Nishizawa, Y.; Kouzai, Y.; Minami, E.; Yano, S.; Koga, H.; Meshi, T.; Nishimura, M. Surface α-1, 3-glucan facilitates fungal stealth infection by interfering with innate immunity in plants. PLoS Pathog. 2012, 8, e1002882. [Google Scholar] [CrossRef] [Green Version]
- Ridout, C.J. Profiles in pathogenesis and mutualism: Powdery mildews. In Plant Relationships; Springer: Berlin/Heidelberg, Germany, 2009; pp. 51–68. [Google Scholar]
- Hückelhoven, R. Powdery mildew susceptibility and biotrophic infection strategies. FEMS Microbiol. Lett. 2005, 245, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Vielba-Fernández, A.; Polonio, Á.; Ruiz-Jiménez, L.; de Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Fungicide resistance in powdery mildew fungi. Microorganisms 2020, 8, 1431. [Google Scholar] [CrossRef]
- Hahn, M.; Mendgen, K. Characterization of in planta—Induced rust genes isolated from a haustorium-specific cDNA library. Mol. Plant-Microbe Interact. 1997, 10, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Fotopoulos, V.; Gilbert, M.J.; Pittman, J.K.; Marvier, A.C.; Buchanan, A.J.; Sauer, N.; Hall, J.L.; Williams, L.E. The monosaccharide transporter gene, AtSTP4, and the cell-wall invertase, At β fruct1, are induced in Arabidopsis during infection with the fungal biotroph Erysiphe cichoracearum. Plant Physiol. 2003, 132, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Both, M.; Csukai, M.; Stumpf, M.P.; Spanu, D. Gene expression profiles of Blumeria graminis indicate dynamic changes to primary metabolism during development of an obligate biotrophic pathogen. Plant Cell 2005, 17, 2107–2122. [Google Scholar] [CrossRef] [Green Version]
- Puthoff, D.P.; Neelam, A.; Ehrenfried, M.L.; Scheffler, B.E.; Ballard, L.; Song, Q.; Campbell, K.B.; Cooper, B.; Tucker, M.L. Analysis of expressed sequence tags from Uromyces appendiculatus hyphae and haustoria and their comparison to sequences from other rust fungi. Phytopathology 2008, 98, 1126–1135. [Google Scholar] [CrossRef] [Green Version]
- Spanu, D.; Abbott, J.C.; Amselem, J.; Burgis, T.A.; Soanes, D.M.; Stüber, K.; Loren van Themaat, E.V.; Brown, J.K.; Butcher, S.A.; Gurr, S.J.; et al. Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 2010, 330, 1543–1546. [Google Scholar] [CrossRef] [PubMed]
- Pliego, C.; Nowara, D.; Bonciani, G.; Gheorghe, D.M.; Xu, R.; Surana, P.; Whigham, E.; Nettleton, D.; Bogdanove, A.J.; Wise, R.P.; et al. Host-induced gene silencing in barley powdery mildew reveals a class of ribonuclease-like effectors. Mol. Plant-Microbe Interact. 2013, 26, 633–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.P.; Raab, T.K.; Somerville, C.R.; Somerville, S.C. Mutations in PMR5 result in powdery mildew resistance and altered cell wall composition. Plant J. 2004, 40, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Bindschedler, L.V.; Burgis, T.A.; Mills, D.J.; Ho, J.T.; Cramer, R.; Spanu, D. In planta proteomics and proteogenomics of the biotrophic barley fungal pathogen Blumeria graminis f. sp. hordei. Mol. Cell. Proteom. 2009, 8, 2368–2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindschedler, L.V.; McGuffin, L.J.; Burgis, T.A.; Spanu, D.; Cramer, R. Proteogenomics and in silico structural and functional annotation of the barley powdery mildew Blumeria graminis f. sp. hordei. Methods 2011, 54, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Pajonk, S.; Micali, C.; O’Connell, R.; Schulze-Lefert, P. Extracellular transport and integration of plant secretory proteins into pathogen-induced cell wall compartments. Plant J. 2009, 57, 986–999. [Google Scholar] [CrossRef]
- Grouffaud, S.; Whisson, S.C.; Birch, R.; van West, P. Towards an understanding on how RxLR-effector proteins are translocated from oomycetes into host cells. Fungal Biol. Rev. 2010, 24, 27–36. [Google Scholar] [CrossRef]
- Rafiqi, M.; Gan, H.; Ravensdale, M.; Lawrence, G.J.; Ellis, J.G.; Jones, D.A.; Hardham, A.R.; Dodds, N. Internalization of flax rust avirulence proteins into flax and tobacco cells can occur in the absence of the pathogen. Plant Cell 2010, 22, 2017–2032. [Google Scholar] [CrossRef] [Green Version]
- Koeck, M.; Hardham, A.R.; Dodds, N. The role of effectors of biotrophic and hemibiotrophic fungi in infection. Cell. Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Micali, C.O.; Neumann, U.; Grunewald, D.; Panstruga, R.; O’Connell, R. Biogenesis of a specialized plant–fungal interface during host cell internalization of Golovinomyces orontii haustoria. Cell. Microbiol. 2011, 13, 210–226. [Google Scholar] [CrossRef] [PubMed]
- Weßling, R.; Schmidt, S.M.; Micali, C.O.; Knaust, F.; Reinhardt, R.; Neumann, U.; van Themaat, E.V.L.; Panstruga, R. Transcriptome analysis of enriched Golovinomyces orontii haustoria by deep 454 pyrosequencing. Fungal Genet. Biol. 2012, 49, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Kracher, B.; Maekawa, T.; Vernaldi, S.; Schulze-Lefert, P.; Ver Loren van Themaat, E. Mosaic genome structure of the barley powdery mildew pathogen and conservation of transcriptional programs in divergent hosts. Proc. Natl. Acad. Sci. USA 2013, 110, E2219–E2228. [Google Scholar] [CrossRef] [PubMed]
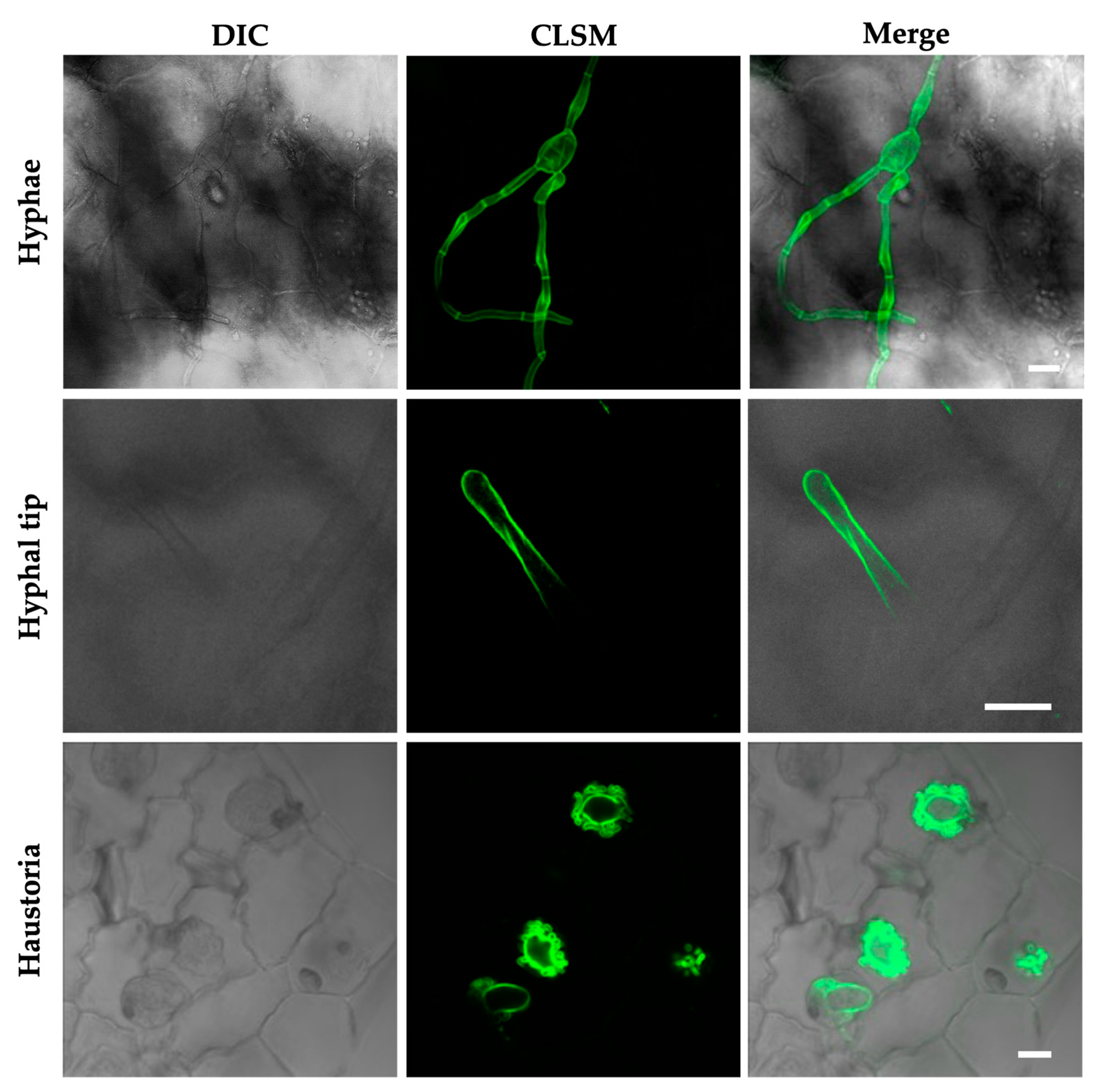
- Martínez-Cruz, J.; Romero, D.; Dávila, J.C.; Pérez-García, A. The Podosphaera xanthii haustorium, the fungal Trojan horse of cucurbit-powdery mildew interactions. Fungal Genet. Biol. 2014, 71, 21–31. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Van der Hoorn, R.A.; Terauchi, R.; Kamoun, S. Emerging concepts in effector biology of plant-associated organisms. Mol. Plant-Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.L.W.; Kunoh, H.; Thomas, B.J.; Nicholson, R.L. Release and visualization of the extracellular matrix of conidia of Blumeria graminis. Mycol. Res. 1999, 103, 547–560. [Google Scholar] [CrossRef]
- Both, M.; Eckert, S.E.; Csukai, M.; Müller, E.; Dimopoulos, G.; Spanu, D. Transcript profiles of Blumeria graminis development during infection reveal a cluster of genes that are potential virulence determinants. Mol. Plant-Microbe Interact. 2005, 18, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Romero, D.; Rivera, M.E.; Cazorla, F.M.; Codina, J.C.; Fernández-Ortuño, D.; Torés, J.A.; Pérez-García, A.; de Vicente, A. Comparative histochemical analyses of oxidative burst and cell wall reinforcement in compatible and incompatible melon–powdery mildew (Podosphaera fusca) interactions. J. Plant Physiol. 2008, 165, 1895–1905. [Google Scholar] [CrossRef]
- Pérez-Garcia, A.; Romero, D.; Fernández-Ortuño, D.; López-Ruiz, F.; de Vicente, A.; Tores, J.A. The powdery mildew fungus Podosphaera fusca (synonym Podosphaera xanthii), a constant threat to cucurbits. Mol. Plant Pathol. 2009, 10, 153–160. [Google Scholar] [CrossRef]
- Heffer, V.; Johnson, K.; Powelson, M.; Shishkoff, N. Identification of Powdery Mildew Fungi anno 2006. Plant Health Instr. 2006. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Gubler, W.D.; Grove, G.G. Epidemiology of Powdery mildews in Agricultural Pathosystems. In The Powdery Mildews: A Comprehensive Treatise; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 169–199. [Google Scholar]
- Bellón-Gómez, D.; Vela-Corcía, D.; Pérez-García, A.; Torés, J.A. Sensitivity of Podosphaera xanthii populations to anti-powdery-mildew fungicides in Spain. Pest Manag. Sci. 2015, 71, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ortuño, D.; Pérez-García, A.; López-Ruiz, F.; Romero, D.; De Vicente, A.; Torés, J.A. Occurrence and distribution of resistance to QoI fungicides in populations of Podosphaera fusca in south central Spain. Eur. J. Plant Pathol. 2006, 115, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Y.; Eyal, H.; Hanania, J. Ultrastructure, autofluorescence, callose deposition and lignification in susceptible and resistant muskmelon leaves infected with the powdery mildew fungus Sphaerotheca fuliginea. Physiol. Mol. Plant Pathol. 1990, 36, 191–204. [Google Scholar] [CrossRef]
- Kuzuya, M.; Yashiro, K.; Tomita, K.; Ezura, H. Powdery mildew (Podosphaera xanthii) resistance in melon is categorized into two types based on inhibition of the infection processes. J. Exp. Bot. 2006, 57, 2093–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fofana, B.; Benhamou, N.; McNally, D.J.; Labbé, C.; Séguin, A.; Bélanger, R.R. Suppression of induced resistance in cucumber through disruption of the flavonoid pathway. Phytopathology 2005, 95, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vela-Corcia, D.; Bautista, R.; de Vicente, A.; Spanu, D.; Perez-Garcia, A. De novo analysis of the epiphytic transcriptome of the cucurbit powdery mildew fungus Podosphaera xanthii and identification of candidate secreted effector proteins. PLoS ONE 2016, 11, e0163379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Miccolis Angelini, R.M.; Pollastro, S.; Rotondo, R.; Laguardia, C.; Abate, D.; Rotolo, C.; Faretra, F. Transcriptome sequence resource for the cucurbit powdery mildew pathogen Podosphaera xanthii. Sci. Data 2019, 6, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonio, Á.; Seoane, P.; Claros, M.G.; Pérez-García, A. The haustorial transcriptome of the cucurbit pathogen Podosphaera xanthii reveals new insights into the biotrophy and pathogenesis of powdery mildew fungi. BMC Genom. 2019, 20, 543. [Google Scholar] [CrossRef] [Green Version]
- Polonio, Á.; Díaz-Martínez, L.; Fernández-Ortuño, D.; de Vicente, A.; Romero, D.; López-Ruiz, F.J.; Pérez-García, A. A hybrid genome assembly resource for Podosphaera xanthii, the main causal agent of powdery mildew disease in cucurbits. Mol. Plant-Microbe Interact. 2021, 34, 319–324. [Google Scholar] [CrossRef]
- Kim, S.; Subramaniyam, S.; Jung, M.; Oh, E.A.; Kim, T.H.; Kim, J.G. Genome resource of Podosphaera xanthii, the host-specific fungal pathogen that causes cucurbit powdery mildew. Mol. Plant-Microbe Interact. 2021, 34, 457–459. [Google Scholar] [CrossRef]
- Polonio, Á.; Pineda, M.; Bautista, R.; Martínez-Cruz, J.; Pérez-Bueno, M.L.; Barón, M.; Pérez-García, A. RNA-seq analysis and fluorescence imaging of melon powdery mildew disease reveal an orchestrated reprogramming of host physiology. Sci. Rep. 2019, 9, 7978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.L.; Chen, B.H.; Chen, X.J.; Guo, Y.Y.; Yang, H.L.; Li, X.Z.; Wang, G.Y. Transcriptome profiling of pumpkin (Cucurbita moschata Duch.) leaves infected with powdery mildew. PLoS ONE 2018, 13, e0190175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Gao, P.; Wan, Y.; Cui, H.; Fan, C.; Liu, S.; Luan, F. Comparative transcriptome profiling of genes and pathways related to resistance against powdery mildew in two contrasting melon genotypes. Sci. Hortic. 2018, 227, 169–180. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Romero, D.; Torés, J.A.; De Vicente, A.; Pérez-García, A. Transient transformation of Podosphaera xanthii by electroporation of conidia. BMC Microbiol. 2015, 15, 20. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cruz, J.; Romero, D.; de Vicente, A.; Pérez-García, A. Transformation of the cucurbit powdery mildew pathogen Podosphaera xanthii by Agrobacterium tumefaciens. New Phytol. 2017, 213, 1961–1973. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cruz, J.; Romero, D.; De Vicente, A.; Pérez-García, A. Transformation by growth onto agro-infiltrated tissues (TGAT), a simple and efficient alternative for transient transformation of the cucurbit powdery mildew pathogen Podosphaera xanthii. Mol. Plant Pathol. 2018, 19, 2502–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Cruz, J.; Romero, D.; de la Torre, F.N.; Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. The functional characterization of Podosphaera xanthii candidate effector genes reveals novel target functions for fungal pathogenicity. Mol. Plant-Microbe Interact. 2018, 31, 914–931. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Jiménez, L.; Polonio, Á.; Vielba-Fernández, A.; Pérez-García, A.; Fernández-Ortuño, D. Gene mining for conserved, non-annotated proteins of Podosphaera xanthii identifies novel target candidates for controlling powdery mildews by spray-induced gene silencing. J. Fungi 2021, 7, 735. [Google Scholar] [CrossRef]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Langner, T.; Göhre, V. Fungal chitinases: Function, regulation, and potential roles in plant/pathogen interactions. Current Genetics 2016, 62, 243–254. [Google Scholar] [CrossRef]
- Jones, L.; Riaz, S.; Morales-Cruz, A.; Amrine, K.C.; McGuire, B.; Gubler, W.D.; Walker, M.A.; Cantu, D. Adaptive genomic structural variation in the grape powdery mildew pathogen, Erysiphe necator. BMC Genom. 2014, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.A.; Schwerdt, J.G.; Shirley, N.J.; Xing, X.; Hsieh, Y.S.; Srivastava, V.; Bulone, V.; Little, A. Analysis of cell wall synthesis and metabolism during early germination of Blumeria graminis f. sp. hordei conidial cells induced in vitro. Cell Surf. 2019, 5, 100030. [Google Scholar] [CrossRef]
- Blair, D.E.; Hekmat, O.; Schüttelkopf, A.W.; Shrestha, B.; Tokuyasu, K.; Withers, S.G.; Van Aalten, D.M. Structure and mechanism of chitin deacetylase from the fungal pathogen Colletotrichum lindemuthianum. Biochemistry 2006, 45, 9416–9426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanni, R.; Martínez-Cruz, J.; Gálvez-Llompart, M.; Fernández-Ortuño, D.; Romero, D.; García-Domènech, R.; Pérez-García, A.; Gálvez, J. Rational Design of Chitin Deacetylase Inhibitors for Sustainable Agricultural Use Based on Molecular Topology. J. Agric. Food Chem. 2022, 70, 13118–13131. [Google Scholar] [CrossRef]
- Wang, Z.; Du, Y.; Li, S.; Xu, X.; Chen, X. A Complete Genome Sequence of Podosphaera xanthii Isolate YZU573, the Causal Agent of Powdery Mildew Isolated from Cucumber in China. Pathogens 2023, 12, 561. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Joly, D.L.; Lin, Y.C.; Tisserant, E.; Feau, N.; Delaruelle, C.; Legué, V.; Kohler, A.; Tanguay, P.; Petre, B.; et al. A comprehensive analysis of genes encoding small secreted proteins identifies candidate effectors in Melampsora larici-populina (poplar leaf rust). Mol. Plant-Microbe Interact. 2012, 25, 279–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghag, S.B. Host induced gene silencing, an emerging science to engineer crop resistance against harmful plant pathogens. Physiol. Mol. Plant Pathol. 2017, 100, 242–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Plant Host | Effector | Strategy | Function | References |
|---|---|---|---|---|---|
| Bipolaris zeicola | Maize | Bz-Cmp | Chitinase-modifying protein | Converts the Zea mays seed chitinase ChitA into a modified form: ChitA-m | [40] |
| Cladosporium fulvum | Tomato | Avr4 | Non-LysM-containing effector | Associates with cell wall chitin to prevent hydrolysis by plant chitinases | [41] |
| Ecp6 | LysM-containing effector | Associates with chitin oligomers to prevent chitin perception by plants | [42] | ||
| Colletotrichum higginsianum | Arabidopsis thaliana | ChELP1 and ChELP2 | LysM-containing effectors | Binds to both chitin polymers and oligomers to protect fungal cell walls from plant digestion and mask chitin oligomers to avoid plant perception | [43] |
| Colletotrichum orbiculare | Nicotiana benthamiana | CoNIS1 | Cytoplasmic effector | Inhibits the kinase activity of BIK1 and BIK1–RbohD interaction to disrupt chitin-induced ROS burst | [44] |
| Fusarium oxysporum | Tomato | FoMep1 and FoSep1 | Proteases | Cleaves tomato chitinases to promote virulence | [45] |
| Cotton | FovPDA1 | Polysaccharide deacetylase | Transforms chitin oligomers into immunogenically inactive chitosan oligomers to circumvent chitin-induced plant immunity | [46] | |
| Fusarium verticillioides | Maize | Fv-Cmp | Chitinase-modifying protein | Truncates the plant chitinases ChitA and ChitB | [47] |
| Magnaporthe oryzae | Rice | AvrPiz-t | Cytoplasmic effector | Targets the host ubiquitin-proteasome system to manipulate plant defenses | [48] |
| GPI-anchored proteins | Non LysM effectors | Works as a camouflage for chitin polymers to help evade plant digestion | [49] | ||
| MoChia1 | LysM-containing effector | Sequesters chitin oligomers and hinders the perception by plant chitin receptors | [50] | ||
| MoNIS1 | Cytoplasmic effector | Harnesses the effector protein AvrPiz-t to interfere with chitin-induced ROS bursts by targeting the rice OsRac1 | [44] | ||
| Slp1 | LysM-containing effector | Associates with chitin oligomers to prevent chitin perception by plants | [51] | ||
| Moniliophthora perniciosa | Cacao | MpChi | LysM-containing effector | Prevents chitin-triggered immunity by sequestering immunogenic chitin fragments | [52] |
| Mycosphaerella fijiensis | Tomato | Avr4 | Non-LysM-containing effector | Associates with cell wall chitin to prevent hydrolysis by plant chitinases | [53] |
| Parastagonospora nodorum | Wheat | SnTox1 | Non-LysM-containing effector | Shields fungal cell walls from plant chitinases | [54] |
| Pestalotiopsis sp | Rice | PesCDA | Chitin deacetylase | Transforms chitin oligomers into immunogenically inactive chitosan oligomers to circumvent chitin-induced plant immunity | [55] |
| Plasmodiophora brassicae | Rapeseed | PbChiB2 and PbChiB4 | Chitinases | Binds to spores and chitin oligomers to suppress chitin-triggered activation of the map kinase proteins MPK3 and MPK6 in the host | [56] |
| Podosphaera xanthii | Melon | PxEWCAs | Chitinases | Released during the pathogen penetration sites to break down immunogenic chitin oligomers, preventing the activation of chitin-triggered immunity | [57] |
| PxLPMO | Monocopper enzyme | Binds and catalyzes colloidal chitin and chito-oligosaccharides generated by plant endochitinases during the growth of haustoria | [58] | ||
| PxCDA | Chitin deacetylase | Converts chitin to chitosan | [59] | ||
| PxCHBE | Chitin-binding effector | Binds to chitin oligomers and prevents the activation of chitin signaling | [60] | ||
| Puccinia striiformis | Wheat | Pst_13661 | Polysaccharide deacetylase | Modifies chitin polymers rendering them recalcitrant to plant digestion | [61] |
| Puccinia graminis | Cereal | PgtCDA | Chitin deacetylase | Converts chitin to chitosan | [62] |
| Rhizoctonia solani | Rice | AGLIP1 | Cytoplamic effector | Suppresses chitin-induced defense gene activation | [63] |
| Sugarbeet | RsLysM | LysM-containing effector | Associates with chitin oligomers to prevent chitin perception by plants | [64] | |
| Rhizophagus irregularis | Medicago truncatula | RiSLM | LysM-containing effector | Protects the hyphae from plant degradation and disguises chitin oligomers from plant detection | [65] |
| Stenocarpella maydis | Maize | Stm-Cmp | Chitinase-modifying protein | Modifies ChitA protein to ChitA-F | [40] |
| Trichoderma atroviride | Arabidopsis thaliana | Tal6 | LysM-containing effector | Protects the hyphae from plant degradation and disguises chitin oligomers from plant detection | [66] |
| Ustilaginoidea virens | Rice | SCRE2 | Cytoplasmic effector | Perturbs chitin-triggered immunity | [67] |
| Ustilago maydis | Maize | UmFly1 | Protease | Truncates the maize chitinase ZmChiA | [68] |
| Verticillium dahlia | Cotton | SSEP1 | Protease | Decapitates the cotton chitinase Chi28 | [69] |
| VdPDA1 | Polysaccharide deacetylase | Transforms chitin oligomers into immunogenically inactive chitosan oligomers to circumvent chitin-induced plant immunity | [46] | ||
| Tomato | Vd2LysM | LysM-containing effector | Binds to both chitin polymers and oligomers to protect fungal cell walls from plant digestion and masks chitin oligomers to avoid plant perception | [70] | |
| Verticillium nonalfalfae | Common hop | VnaChtBP | Non-LysM-containing effector | Interacts with both chitin polymers and oligomers, allowing the fungus to escape from both plant chitinases and chitin receptors | [71] |
| Zymoseptoria tritici | Wheat | Mg1LysM and Mg3LysM | LysM-containing effectors | Protects fungal hyphae against host chitinases | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakhat, N.; Vielba-Fernández, A.; Padilla-Roji, I.; Martínez-Cruz, J.; Polonio, Á.; Fernández-Ortuño, D.; Pérez-García, A. Suppression of Chitin-Triggered Immunity by Plant Fungal Pathogens: A Case Study of the Cucurbit Powdery Mildew Fungus Podosphaera xanthii. J. Fungi 2023, 9, 771. https://doi.org/10.3390/jof9070771
Bakhat N, Vielba-Fernández A, Padilla-Roji I, Martínez-Cruz J, Polonio Á, Fernández-Ortuño D, Pérez-García A. Suppression of Chitin-Triggered Immunity by Plant Fungal Pathogens: A Case Study of the Cucurbit Powdery Mildew Fungus Podosphaera xanthii. Journal of Fungi. 2023; 9(7):771. https://doi.org/10.3390/jof9070771
Chicago/Turabian StyleBakhat, Nisrine, Alejandra Vielba-Fernández, Isabel Padilla-Roji, Jesús Martínez-Cruz, Álvaro Polonio, Dolores Fernández-Ortuño, and Alejandro Pérez-García. 2023. "Suppression of Chitin-Triggered Immunity by Plant Fungal Pathogens: A Case Study of the Cucurbit Powdery Mildew Fungus Podosphaera xanthii" Journal of Fungi 9, no. 7: 771. https://doi.org/10.3390/jof9070771
APA StyleBakhat, N., Vielba-Fernández, A., Padilla-Roji, I., Martínez-Cruz, J., Polonio, Á., Fernández-Ortuño, D., & Pérez-García, A. (2023). Suppression of Chitin-Triggered Immunity by Plant Fungal Pathogens: A Case Study of the Cucurbit Powdery Mildew Fungus Podosphaera xanthii. Journal of Fungi, 9(7), 771. https://doi.org/10.3390/jof9070771