
Meyerozyma caribbica Isolated from Vinasse-Irrigated Sugarcane Plantation Soil: A Promising Yeast for Ethanol and Xylitol Production in Biorefineries

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Characteristics and Sampling
2.2. Soil Processing and Yeast Isolation
2.3. Yeast Molecular Identification
2.4. Physiological Tests
2.5. Acid Hydrolysates Production and Molasses
2.6. Fermentation Assays
2.7. Substrate Detoxification
2.8. Effect of Acetate on Fermentation
2.9. Analytical Methods
2.10. Statistical Analysis
3. Results
3.1. Yeast Isolation and Identification
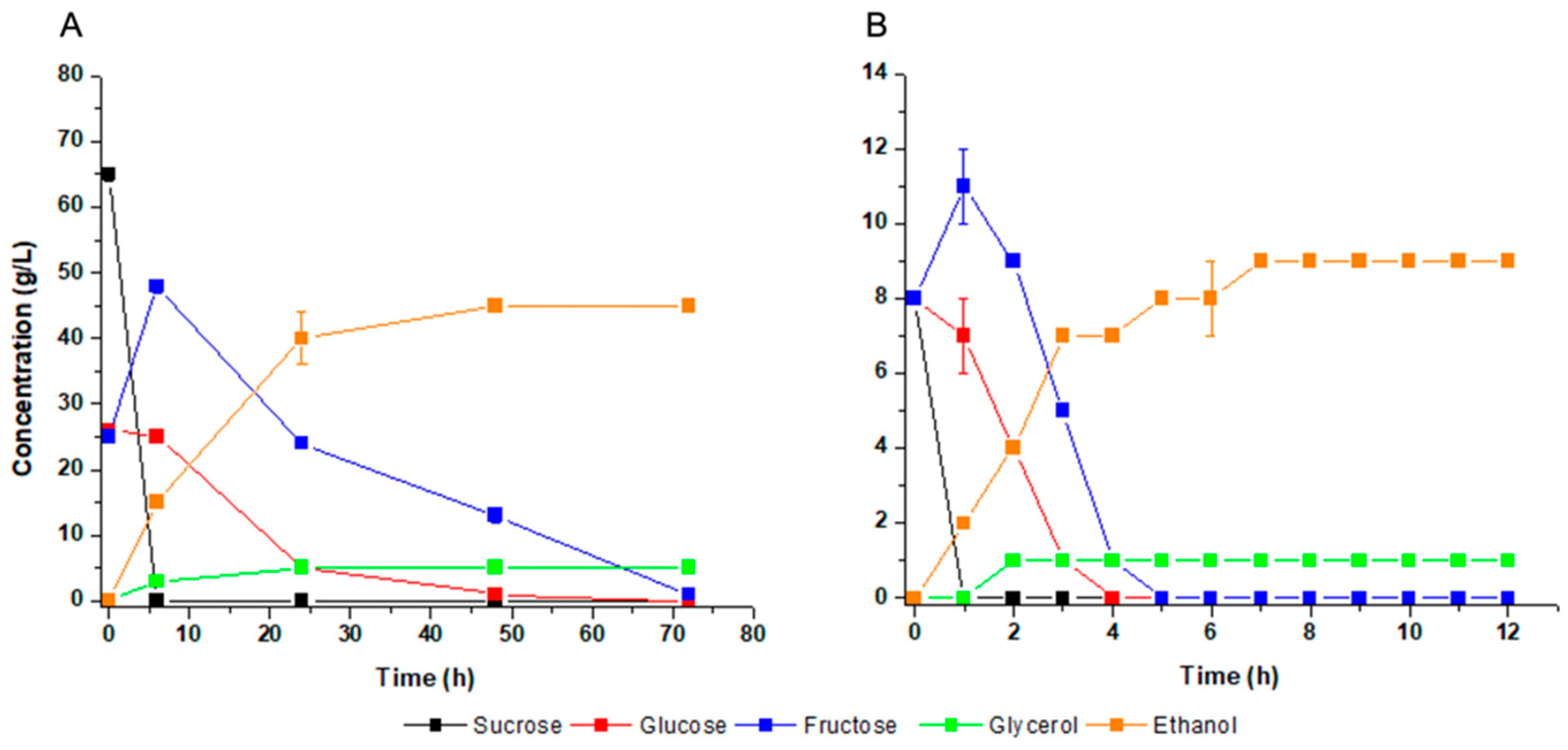
3.2. Ethanol Fermentation of Mineral Medium and Carbon Distribution Analysis
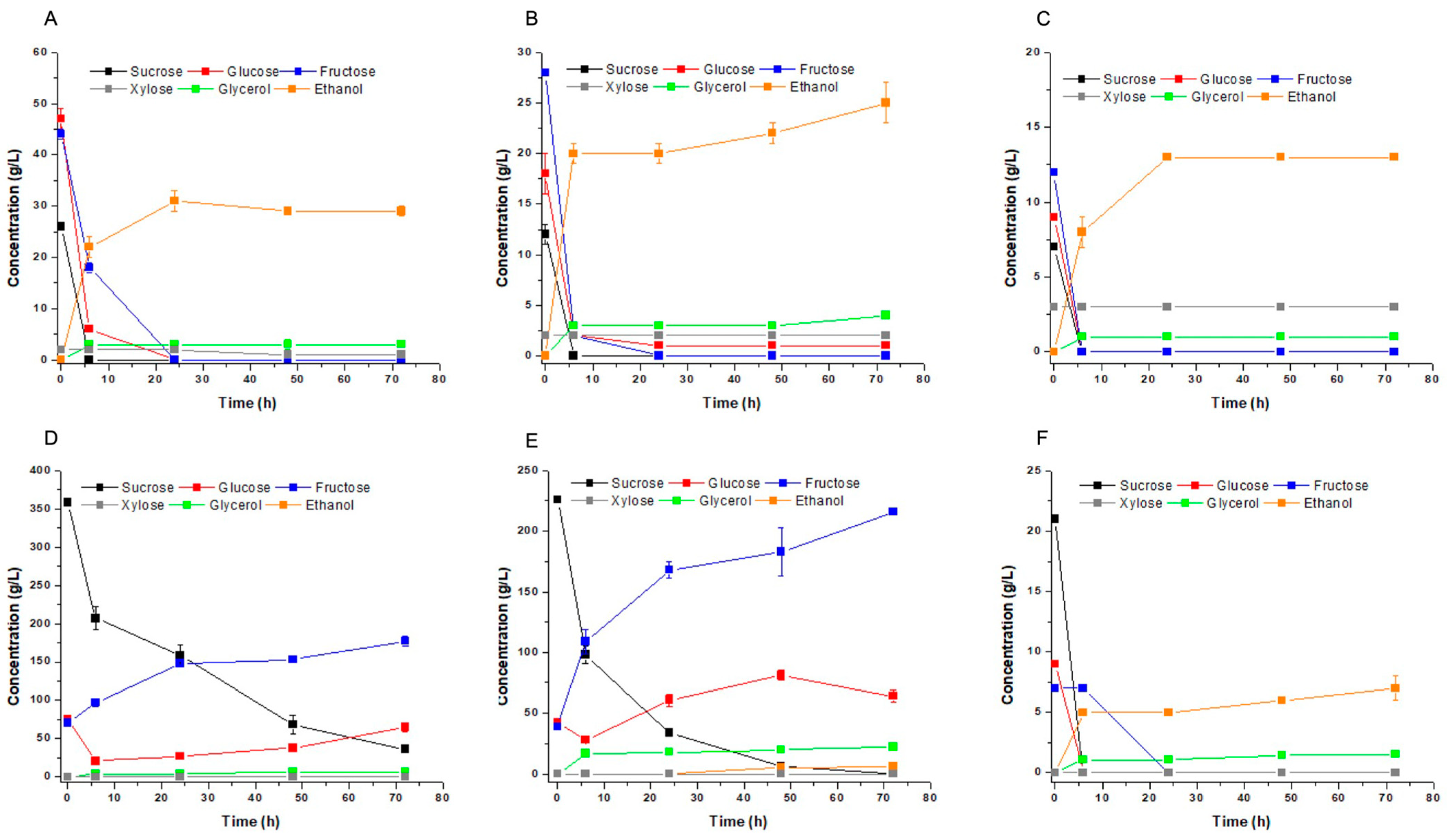
3.3. Ethanol Fermentation of Industrial Substrates
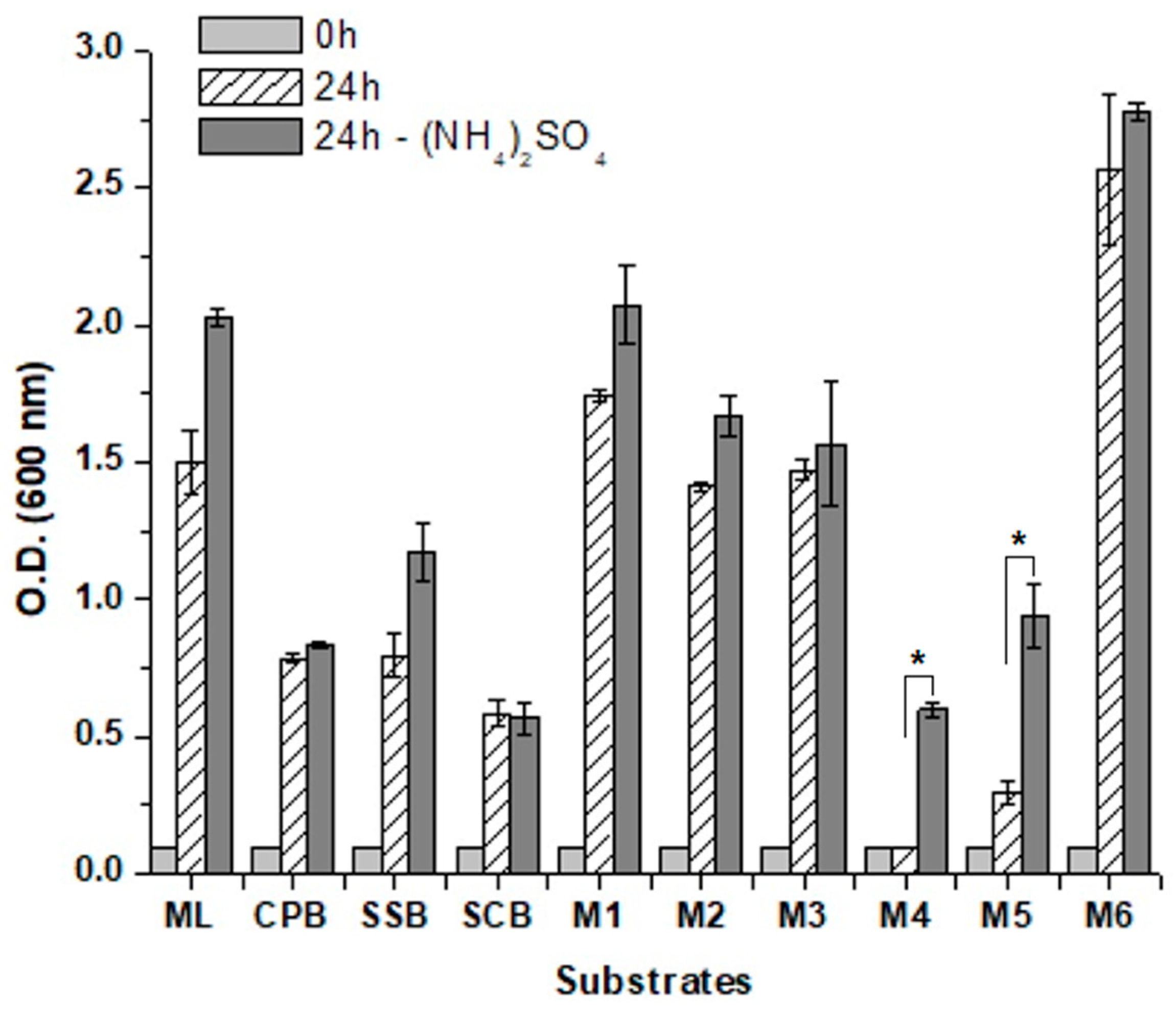
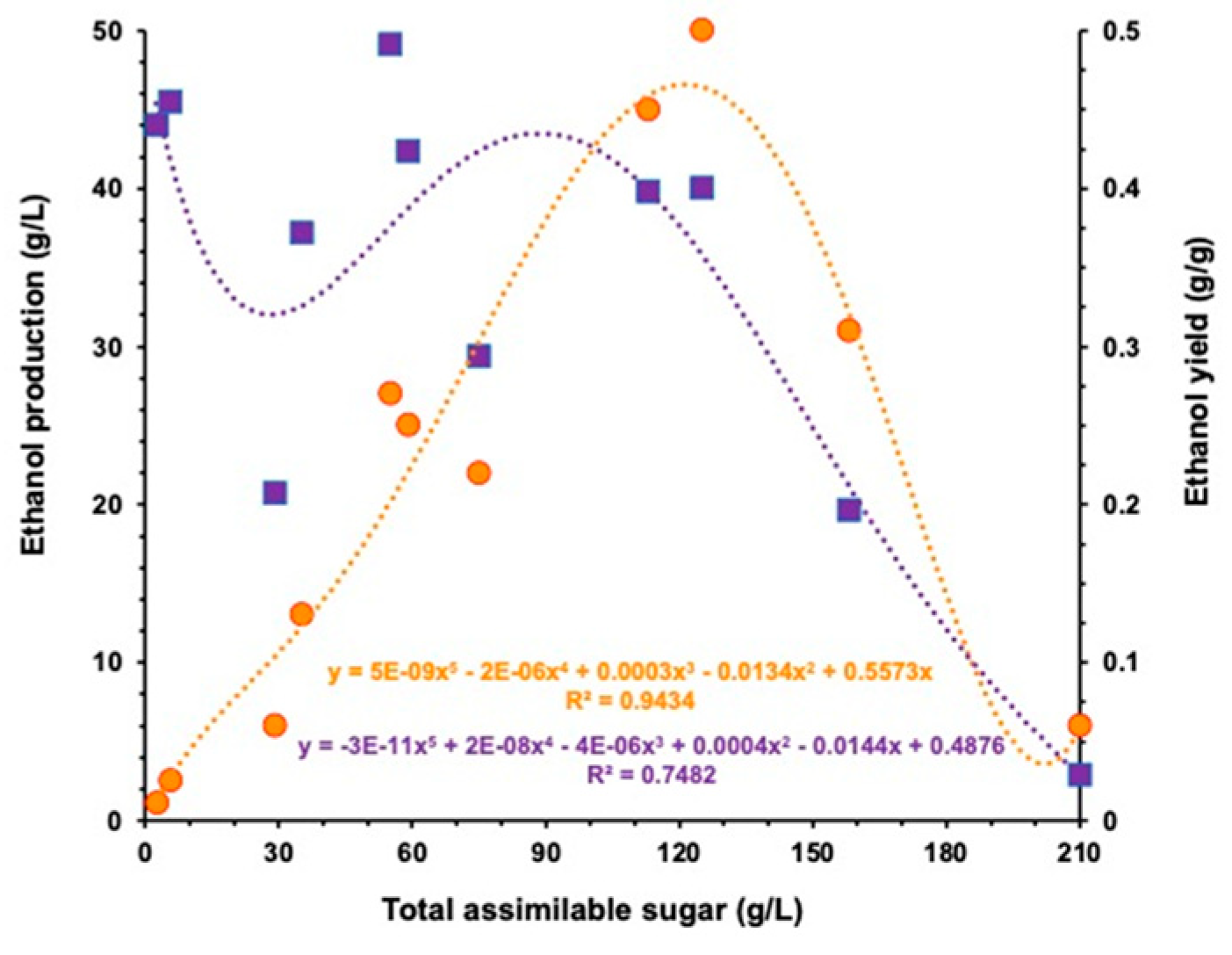
3.4. Fermentation of Mixed Industrial Substrates
3.5. Xylitol Production
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yaashikaa, P.R.; Kumar, P.S.; Saravanan, A.; Karishma, S.; Rangasamy, G. A biotechnological roadmap for decarbonization systems combined into bioenergy production: Prelude of environmental life-cycle assessment. Chemosphere 2023, 329, 138670. [Google Scholar] [CrossRef]
- Singh, N.; Singhania, R.R.; Nigam, P.S.; Dong, C.D.; Patel, A.K.; Puri, M. Global status of lignocellulosic biorefinery: Challenges and perspectives. Bioresour. Technol. 2022, 344, 126415. [Google Scholar] [CrossRef]
- Freitas, J.V.; Bilatto, S.; Squinca, P.; Pinto, A.S.; Brondi, M.G.; Bonaci, T.J.; Farinas, C.S. Sugarcane biorefineries: Potential opportunities towards shifting from wastes to products. Ind. Crops Prod. 2021, 172, 114057. [Google Scholar]
- Shapiro, A.J.; O’Dea, R.M.; Li, S.C.; Ajah, J.C.; Bass, G.F.; Epps, T.H. Engineering Innovations, Challenges, and Opportunities for Lignocellulosic Biorefineries: Leveraging Biobased Polymer Production. Ann. Rev. Chem. Biomol. Eng. 2023, 14, 109–140. [Google Scholar] [CrossRef]
- Yadav, A.; Sharma, V.; Tsai, M.L.; Chen, C.W.; Sun, P.P.; Nargotra, P.; Wang, J.X.; Dong, C.D. Development of lignocellulosic biorefineries for the sustainable production of biofuels: Towards circular bioeconomy. Bioresour. Technol. 2023, 381, 129145. [Google Scholar] [CrossRef] [PubMed]
- Solarte-Toro, J.C.; Cardona Alzate, C.A. Sustainability of Biorefineries: Challenges and Perspectives. Energies 2023, 16, 3786. [Google Scholar] [CrossRef]
- Rossetto, R.; Ramos, N.P.; de Matos Pires, R.C.; Xavier, M.A.; Cantarella, H.; Landell, M.G.A. Sustainability in sugarcane supply chain in Brazil: Issues and way forward. Sugar Tech 2022, 24, 941–966. [Google Scholar] [CrossRef]
- Parsaee, M.; Kiani, M.K.D.; Karimi, K. A review of biogas production from sugarcane vinasse. Biomass Bioenergy 2019, 122, 117–125. [Google Scholar] [CrossRef]
- Antunes, F.A.; Thomé, L.C.; Santos, J.C.; Ingle, A.P.; Costa, C.B.; Dos Anjos, V.; Da Silva, S.S. Multi-scale study of the integrated use of the carbohydrate fractions of sugarcane bagasse for ethanol and xylitol production. Renew. Energy 2021, 163, 1343–1355. [Google Scholar] [CrossRef]
- Escobar, E.L.N.; Suota, M.J.; Ramos, L.P.; Corazza, M.L. Combination of green solvents for efficient sugarcane bagasse fractionation. Biomass Bioenergy 2022, 161, 106482. [Google Scholar] [CrossRef]
- Dutra, E.D.; Neto, A.G.B.; de Souza, R.B.; de Morais, M.A., Jr.; Tabosa, J.N.; Menezes, R.S.C. Ethanol production from the stem juice of different sweet sorghum cultivars in the state of Pernambuco, Northeast of Brazil. Sugar Tech 2013, 15, 316–321. [Google Scholar] [CrossRef]
- Reshmy, R.; Paulose, T.A.P.; Philip, E.; Thomas, D.; Madhavan, A.; Sirohi, R.; Sindhu, R. Updates on high value products from cellulosic biorefinery. Fuel 2022, 308, 122056. [Google Scholar] [CrossRef]
- Dutra, E.D.; Alencar, B.R.A.; Galdino, J.J.; Tabosa, J.N.; Menezes, R.S.C.; de Araújo Filho, R.N.; Costa Primo, D.; Piscoya, V.C.; Antunes, J.E.L. First and Second Generation of Ethanol Production for Five Sweet Sorghum Cultivars during Soft Dough Grain. J. Exp. Agric. Int. 2018, 25, 1–12. [Google Scholar] [CrossRef]
- Reis, A.L.S.; Damilano, E.D.; Menezes, R.S.C.; de Morais Jr, M.A. Second-generation ethanol from sugarcane and sweet sorghum bagasses using the yeast Dekkera bruxellensis. Ind. Crops Prod. 2016, 92, 255–262. [Google Scholar] [CrossRef]
- Vaz, F.L.; da Rocha Lins, J.; Alencar, B.R.A.; de Abreu, I.B.S.; Vidal, E.E.; Ribeiro, E.; Dutra, E.D. Chemical pretreatment of sugarcane bagasse with liquid fraction recycling. Renew. Energy 2021, 174, 666–673. [Google Scholar] [CrossRef]
- Santos, T.N.; Dutra, E.D.; do Prado, A.G.; Leite, F.C.B.; de Souza, R.D.F.R.; dos Santos, D.C.; Menezes, R.S.C. Potential for biofuels from the biomass of prickly pear cladodes: Challenges for bioethanol and biogas production in dry areas. Biomass Bioenergy 2016, 85, 215–222. [Google Scholar] [CrossRef]
- Alencar, B.R.A.; Dutra, E.D.; Sampaio, E.V.D.S.B.; Menezes, R.S.C.; Morais Jr, M.A. Enzymatic hydrolysis of cactus pear varieties with high solids loading for bioethanol production. Bioresour. Technol. 2018, 250, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Cadete, R.M.; Melo, M.A.; Dussán, K.J.; Rodrigues, R.C.; Silva, S.S.; Zilli, J.E.; Vital, M.J.; Gomes, F.C.; Lachance, M.A.; Rosa, C.A. Diversity and physiological characterization of D-xylose-fermenting yeasts isolated from the Brazilian Amazonian Forest. PLoS ONE 2012, 7, e43135. [Google Scholar] [CrossRef]
- De Souza, R.F.R.; Dutra, E.D.; Leite, F.C.B.; Cadete, R.M.; Rosa, C.A.; Stambuk, B.U.; Sramford, T.L.M.; de Morais, M.A., Jr. Production of ethanol fuel from enzyme-treated sugarcane bagasse hydrolysate using d-xylose-fermenting wild yeast isolated from Brazilian biomes. 3 Biotech 2018, 8, 312. [Google Scholar] [CrossRef]
- Rech, F.R.; Fontana, R.C.; Rosa, C.A.; Camassola, M.; Ayub, M.A.Z.; Dillon, A.J.P. Fermentation of hexoses and pentoses from sugarcane bagasse hydrolysates into ethanol by Spathaspora hagerdaliae. Bioprocess Biosyst. Eng. 2019, 42, 83–92. [Google Scholar] [CrossRef]
- Campos, V.J.; Ribeiro, L.E.; Albuini, F.M.; de Castro, A.G.; Fontes, P.P.; da Silveira, W.B.; Rosa, C.A.; Fietto, L.G. Physiological comparisons among Spathaspora passalidarum, Spathaspora arborariae, and Scheffersomyces stipitis reveal the bottlenecks for their use in the production of second-generation ethanol. Braz. J. Microbiol. 2022, 53, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Tadioto, V.; Milani, L.M.; Barrilli, É.T.; Baptista, C.W.; Bohn, L.; Dresch, A.; Harakava, R.; Fogolari, O.; Mibielli, G.M.; Bender, J.P.; et al. Analysis of glucose and xylose metabolism in new indigenous Meyerozyma caribbica strains isolated from corn residues. World J. Microbiol. Biotechnol. 2022, 38, 35. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.E.; Lockhart, S.R. Recent Taxonomic Developments with Candida and Other Opportunistic Yeasts. Curr. Fungal Infect. Rep. 2012, 6, 170–177. [Google Scholar] [CrossRef]
- Limtong, S.; Kaewwichian, R.; Yongmanitchai, W.; Kawasaki, H. Diversity of culturable yeasts in phylloplane of sugarcane in Thailand and their capability to produce indole-3-acetic acid. World J. Microbiol. Biotechnol. 2014, 30, 1785–1796. [Google Scholar] [CrossRef]
- Basílio, A.C.; de Araújo, P.R.; de Morais, J.O.F.; da Silva Filho, E.A.; de Morais, M.A., Jr.; Simões, D.A. Detection and identification of wild yeast contaminants of the industrial fuel ethanol fermentation process. Curr. Microbiol. 2008, 56, 322–326. [Google Scholar] [CrossRef]
- Vila Nova, M.X.; Schuler, A.R.P.; Brasileiro, B.T.R.V.; Morais Jr, M.A. Yeast species involved in artisanal cachaça fermentation in three stills with different technological levels in Pernambuco, Brazil. Food Microbiol. 2009, 26, 460–466. [Google Scholar] [CrossRef]
- Martinez, S.J.; Bressani, A.P.P.; Simão, J.B.P.; Pylro, V.S.; Dias, D.R.; Schwan, R.F. Dominant microbial communities and biochemical profile of pulped natural fermented coffees growing in different altitudes. Food Res. Int. 2022, 159, 111605. [Google Scholar] [CrossRef]
- Tiwari, S.; Jadhav, R.; Avchar, R.; Lanjekar, V.; Datar, M.; Baghela, A. Nectar Yeast Community of Tropical Flowering Plants and Assessment of Their Osmotolerance and Xylitol-Producing Potential. Curr. Microbiol. 2021, 79, 28. [Google Scholar] [CrossRef] [PubMed]
- Matos, Í.T.S.R.; de Souza, V.A.; D’Angelo, G.D.R.; Astolfi Filho, S.; do Carmo, E.J.; Vital, M.J.S. Yeasts with Fermentative Potential Associated with Fruits of Camu-Camu (Myrciaria dubia, Kunth) from North of Brazilian Amazon. Sci. World J. 2021, 2021, 9929059. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. Online 2015, 11, 43–48. [Google Scholar] [CrossRef]
- Nylander, J.A.A.; Ronquist, F.; Huelsenbeck, J.P.; Nieves-Aldrey, J.L. Bayesian phylogenetic analysis of combined data. Syst. Biol. 2004, 53, 47–67. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Zhang, J.; Ling, H.; Ping, W.; Huang, W.; Ge, J.; Xu, J. Optimization of pH and acetic acid concentration for bioconversion of hemicellulose from corncobs to xylitol by Candida tropicalis. Biochem. Eng. J. 2009, 43, 203–207. [Google Scholar] [CrossRef]
- Teles, G.H.; da Silva, J.M.; Mendonça, A.A.; de Morais, M.A., Jr.; de Barros Pita, W. First aspects on acetate metabolism in the yeast Dekkera bruxellensis: A few keys for improving ethanol fermentation. Yeast 2018, 35, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.F.; Bassi, A.P.; Avansini, S.H.; Neto, A.G.; Brasileiro, B.T.; Ceccato-Antonini, S.R.; de Morais, M.A., Jr. The physiological characteristics of the yeast Dekkera bruxellensis in fully fermentative conditions with cell recycling and in mixed cultures with Saccharomyces cerevisiae. Antonie Van Leeuwenhoek 2012, 101, 529–539. [Google Scholar] [CrossRef]
- Pereira, L.F.; Lucatti, E.; Basso, L.C.; de Morais, M.A., Jr. The fermentation of sugarcane molasses by Dekkera bruxellensis and the mobilization of reserve carbohydrates. Antonie Van Leeuwenhoek 2014, 105, 481–489. [Google Scholar] [CrossRef]
- Ahuja, V.; Bhatt, A.K.; Mehta, S.; Sharma, V.; Rathour, R.K.; Sheetal. Xylitol production by Pseudomonas gessardii VXlt-16 from sugarcane bagasse hydrolysate and cost analysis. Bioprocess Biosyst. Eng. 2022, 45, 1019–1031. [Google Scholar] [CrossRef]
- Vaughan-Martini, A.; Kurtzman, C.P.; Meyer, S.A.; O’Neill, E.B. Two new species in the Pichia guilliermondii clade: Pichia caribbica sp. nov., the ascosporic state of Candida fermentati, and Candida carpophila comb. nov. FEMS Yeast Res. 2005, 5, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romi, W.; Keisam, S.; Ahmed, G.; Jeyaram, K. Reliable differentiation of Meyerozyma guilliermondii from Meyerozyma caribbica by internal transcribed spacer restriction fingerprinting. BMC Microbiol. 2014, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavana, B.K.; Mudliar, S.N.; Bokade, V.V.; Debnath, S. Effect of furfural, acetic acid and 5-hydroxymethylfurfural on yeast growth and xylitol fermentation using Pichia stipitis NCIM 3497. Biomass Convers. Biorefinery 2022. [Google Scholar] [CrossRef]
- Nagarajan, A.; Thulasinathan, B.; Arivalagan, P.; Alagarsamy, A.; Muthuramalingam, J.B.; Thangarasu, S.D.; Thangavel, K. Particle size influence on the composition of sugars in corncob hemicellulose hydrolysate for xylose fermentation by Meyerozyma caribbica. Bioresour. Technol. 2021, 340, 125677. [Google Scholar] [CrossRef]
- Veras, H.C.T.; Parachin, N.S.; Almeida, J.R.M. Comparative assessment of fermentative capacity of different xylose-consuming yeasts. Microb. Cell Factories 2017, 16, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, S.; Xu, H.; Oh, E.J.; Kim, H.; Lesmana, A.; Jeong, D.; Zhang, G.; Tsai, C.S.; Jin, Y.S.; Kim, S.R. Glucose repression can be alleviated by reducing glucose phosphorylation rate in Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trichez, D.; Steindorff, A.S.; Soares, C.E.V.F.; Formighieri, E.F.; Almeida, J.R.M. Physiological and comparative genomic analysis of new isolated yeasts Spathaspora sp. JA1 and Meyerozyma caribbica JA9 reveal insights into xylitol production. FEMS Yeast Res. 2019, 19, foz034. [Google Scholar] [CrossRef]
- Parawira, W.; Tekere, M. Biotechnological strategies to overcome inhibitors in lignocellulose hydrolysates for ethanol production: Review. Crit. Rev. Biotechnol. 2011, 31, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Chandrasekaran, G.; Brul, S.; Smits, G.J. Yeast adaptation to weak acids prevents futile energy expenditure. Front. Microbiol. 2013, 4, 142. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TFC | Total N | Conductivity | Composition | |||||
|---|---|---|---|---|---|---|---|---|
| Substrate | (g/L) | (%) | C/N | (ms/cm) | ML | SCB | SSB | CPB |
| ML | 107.51 | 4.01 | 160.8 | 33.1 | 100% | -- | -- | -- |
| SCB | 21.27 | 1.81 | 70.5 | 122.0 | -- | 100% | -- | -- |
| SSB | 17.96 | 1.96 | 55.0 | 109.8 | -- | -- | 100% | -- |
| CPB | 64.09 | 4.21 | 91.4 | 57.0 | -- | -- | -- | 100% |
| M1 | 127.42 | 3.21 | 238.2 | 54.7 | 70% * | 10% | 10% | 10% |
| M2 | 94.66 | 3.31 | 171.6 | 64.8 | 50% * | 16.7% | 16.7% | 16.6% |
| M3 | 52.31 | 2.46 | 127.5 | 83.8 | 25% * | 25% | 25% | 25% |
| M4 | 306.97 | 6.76 | 272.5 | 29.9 | 50% | 16.7% | 16.7% | 16.6% |
| M5 | 108.8 | 5.11 | 127.7 | 56.4 | 25% | 25% | 25% | 25% |
| M6 | 36.63 | 3.26 | 67.4 | 93.4 | 10% | 30% | 30% | 30% |
| Parameter | Glucose | Xylose | Glucose + Xylose |
|---|---|---|---|
| Initial glucose (g L−1) | 37.85 (±0.90) | - | 17.00 (±0.23) |
| Initial xylose (g L−1) | - | 38.82 (±0.96) | 18.95 (±0.20) |
| Final glucose (g L−1) | 0.0 (±0.00) | - | 0.0 (±0.00) |
| Final xylose (g L−1) | - | 38.62 (±0.99) | 14.84 (±0.31) |
| Ethanol produced (g L−1) | 16.19 (±1.09) | 0.00 | 9.29 (±1.73) |
| Glycerol produced (g L−1) | 0.67 (±0.12) | 0.00 | 0.00 |
| Ethanol yield (g/g) | 0.43 (±0.03) | 0.00 | 0.44 (±0.04) |
| Glycerol yield (g/g) | 0.02 (>0.00) | 0.00 | 0.00 |
| CO2 yield (g/g) * | 0.41 | 0.00 | 0.42 |
| Biomass yield (g/g) | 0.03 | 0.00 | 0.09 |
| Carbon balance (%) | 89% | 0.00 | 95% |
| Parameters | Mineral Medium | +Acetic Acid 1.6 g/L | +Acetic Acid 3.7 g/L |
|---|---|---|---|
| Xylose consumed (g/L) | 63.58 ± 5.26 | 50.30 ± 3.05 | 0.00 |
| Glucose consumed (g/L) | 11.10 ± 0.69 | 11.74 ± 0.966 | 2.50 ± 0.63 |
| Ethanol produced (g/L) | 11.67 ± 1.01 | 12.41 ± 1.33 | 0.00 |
| Xylitol produced (g/L) | 11.67 ± 1.1 | 4.85 ± 0.57 | 0.00 |
| Xylitol yield (g/g) | 0.18 ± 0.04 | 0.1 ± 0.01 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alencar, B.R.A.; de Freitas, R.A.A.; Guimarães, V.E.P.; Silva, R.K.; Elsztein, C.; da Silva, S.P.; Dutra, E.D.; de Morais Junior, M.A.; de Souza, R.B. Meyerozyma caribbica Isolated from Vinasse-Irrigated Sugarcane Plantation Soil: A Promising Yeast for Ethanol and Xylitol Production in Biorefineries. J. Fungi 2023, 9, 789. https://doi.org/10.3390/jof9080789
Alencar BRA, de Freitas RAA, Guimarães VEP, Silva RK, Elsztein C, da Silva SP, Dutra ED, de Morais Junior MA, de Souza RB. Meyerozyma caribbica Isolated from Vinasse-Irrigated Sugarcane Plantation Soil: A Promising Yeast for Ethanol and Xylitol Production in Biorefineries. Journal of Fungi. 2023; 9(8):789. https://doi.org/10.3390/jof9080789
Chicago/Turabian StyleAlencar, Bárbara Ribeiro Alves, Renan Anderson Alves de Freitas, Victor Emanuel Petrício Guimarães, Rayssa Karla Silva, Carolina Elsztein, Suzyanne Porfírio da Silva, Emmanuel Damilano Dutra, Marcos Antonio de Morais Junior, and Rafael Barros de Souza. 2023. "Meyerozyma caribbica Isolated from Vinasse-Irrigated Sugarcane Plantation Soil: A Promising Yeast for Ethanol and Xylitol Production in Biorefineries" Journal of Fungi 9, no. 8: 789. https://doi.org/10.3390/jof9080789
APA StyleAlencar, B. R. A., de Freitas, R. A. A., Guimarães, V. E. P., Silva, R. K., Elsztein, C., da Silva, S. P., Dutra, E. D., de Morais Junior, M. A., & de Souza, R. B. (2023). Meyerozyma caribbica Isolated from Vinasse-Irrigated Sugarcane Plantation Soil: A Promising Yeast for Ethanol and Xylitol Production in Biorefineries. Journal of Fungi, 9(8), 789. https://doi.org/10.3390/jof9080789