
Optimization and Process Effect for Microalgae Carbon Dioxide Fixation Technology Applications Based on Carbon Capture: A Comprehensive Review




Abstract
:1. Introduction
2. Mechanism of Carbon Dioxide Fixation Technology by Microalgae
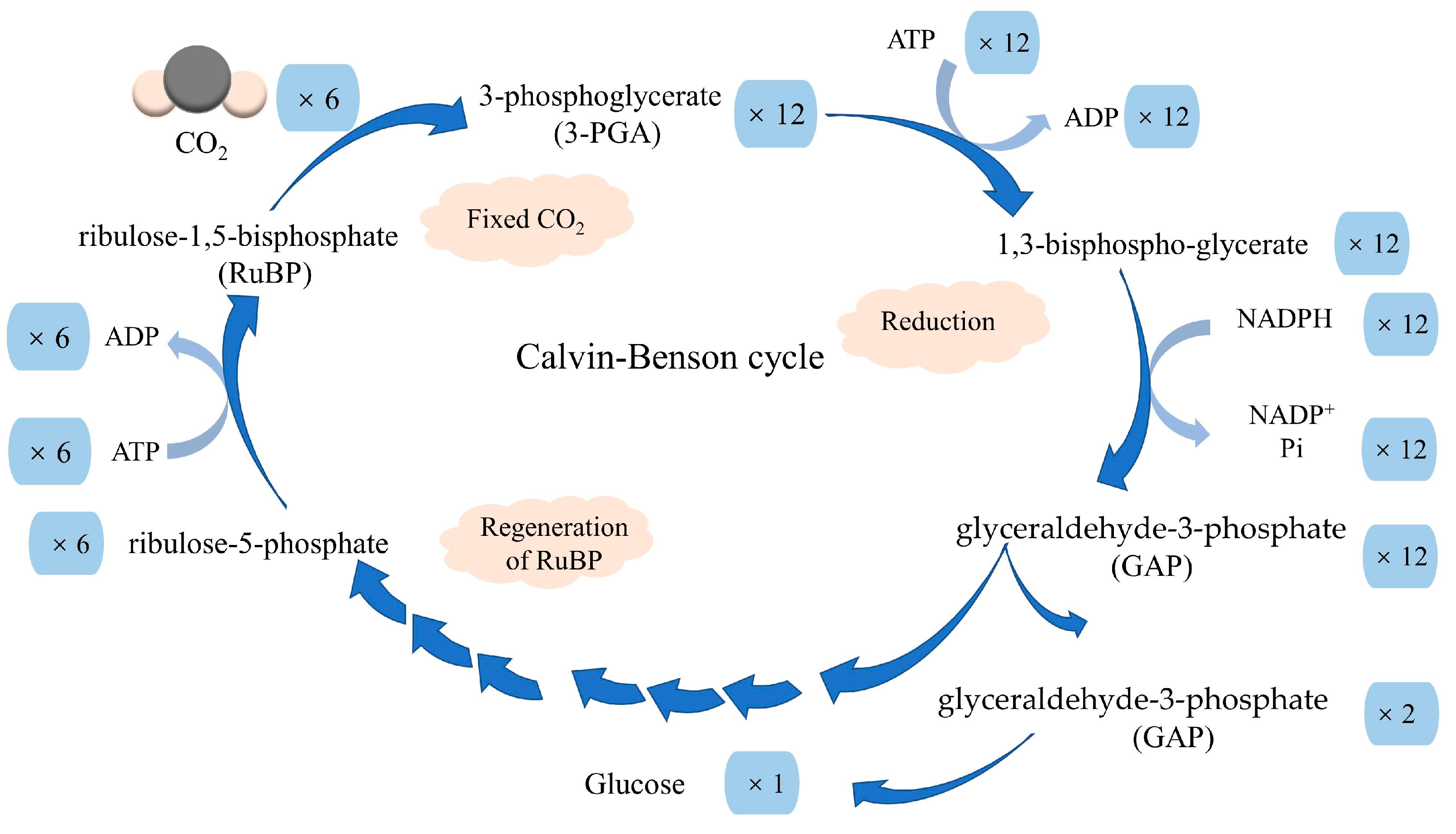
2.1. The Photosynthetic Carbon Metabolism Pathway
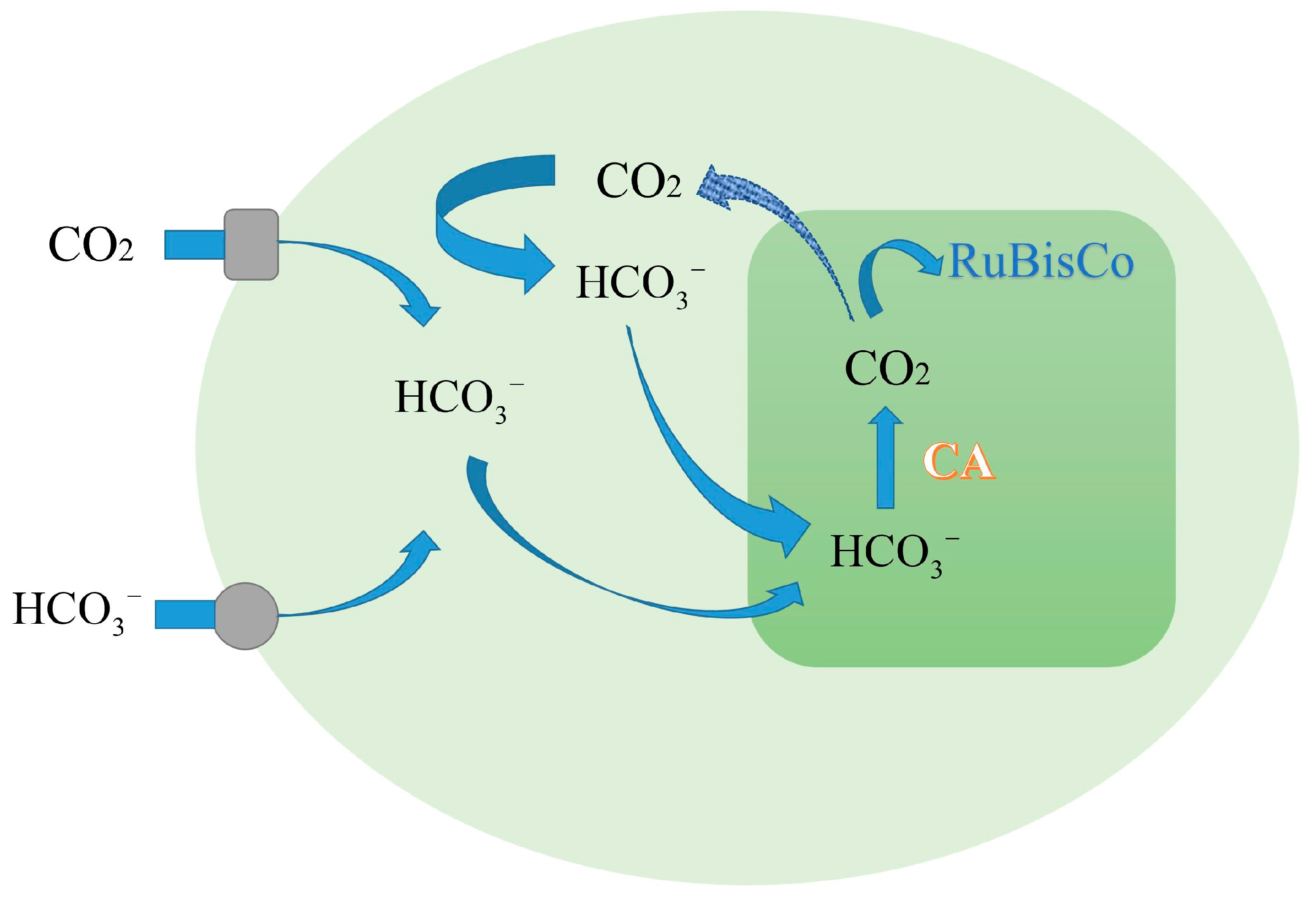
2.2. Carbon Dioxide Concentrating-Mechanism
3. Environmental Conditions for the Growth of the Microalgae
3.1. Light
3.2. Temperature
3.3. pH
3.4. Nutrient Element
3.5. Carbon Dioxide Concentration
4. Strategies for Improving Photosynthetic Efficiency in Microalgae
4.1. Random Mutagenesis
4.1.1. Chemical Mutagenesis
4.1.2. Physical Mutagenesis
4.2. Adaptive Laboratory Evolution
4.3. Genetic Engineering
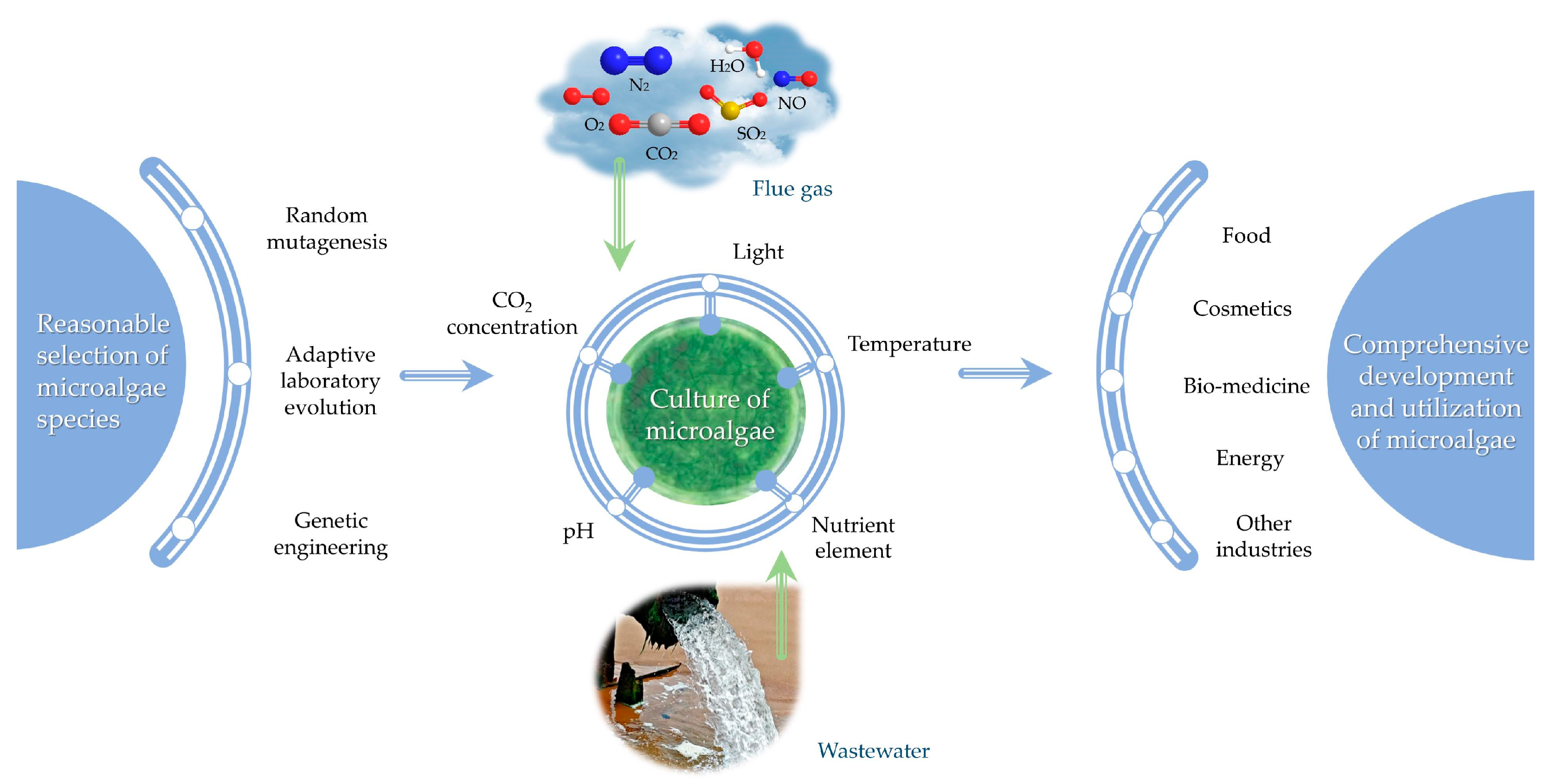
5. Application of Carbon Dioxide Fixation Technology in Microalgae
5.1. In the Atmosphere
5.2. Flue Gas
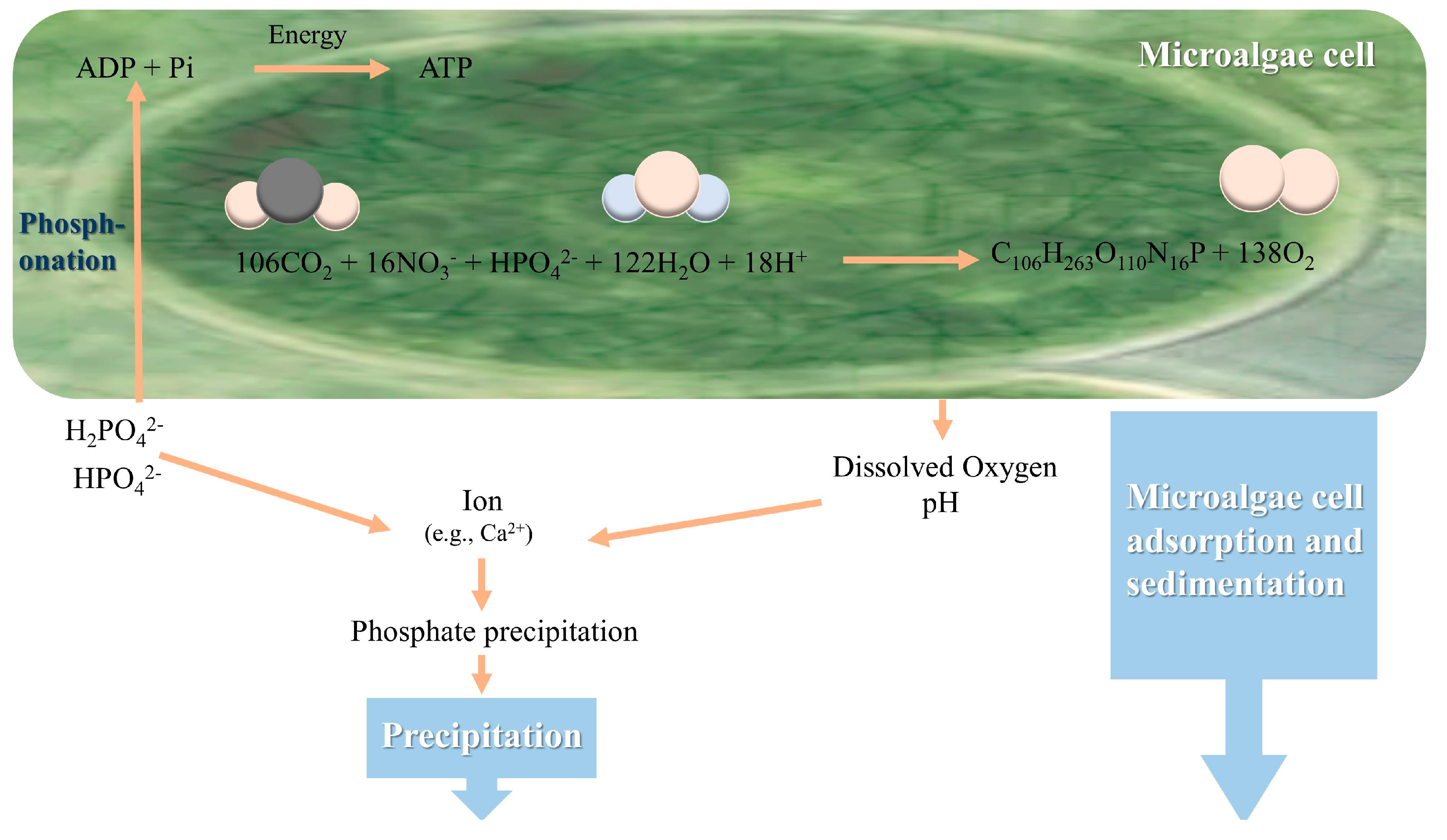
5.3. In Wastewater
5.4. Other Applications
6. A Future Prospect for Microalgae Carbon Dioxide Fixation Technology
6.1. Selection of Seeds Rationally
6.2. To Optimize the Culture Conditions of Microalgae
6.3. To Construct of Microalgae Carbon Dioxide Fixation Technology as the Core of the Industrial Model
6.4. Comprehensive Development and Utilization of High-Value-Added Products of Microalgae
6.5. Potential for Carbon Fixation in the Microalgae
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Strains | Medium | Initial Cell Density | Cycles | Stresses | Outcome | Ref. |
|---|---|---|---|---|---|---|
| Chlorella sp. | BG11 | OD750 of 0.1 | 31 cycles (97 days) | 10% CO2 | The maximal biomass concentration of AE10 was 3.68 ± 0.08 g/L in 30% CO2, which was 2.94 fold compared to the original strain. | [70] |
| Chlamydomonas reinhardtii | TAP and TAP-N | OD680 of 0.1 | 28 cycles (84 days) | Nitrogen starvation | ALE combined with nitrogen starvation substantially increased total lipid production, particularly for low-starch mutants. The endpoint strain of cc4334 under nitrogen starvation stress had the highest lipid productivity. | [129] |
| Schizochytrium sp. HX-308 | Main culture medium contained 40 g/L glucose and 0.4 g/L yeast extract | - | 40 cycles (40 days) | High oxygen | The adapted strain generated higher cell dry weight and lower lipid accumulation. | [90,130] |
| Schizochytrium sp. HX-308 | Medium with 30 g/L NaCl at a concentration of 1% v/v | - | 150 days | High salinity | The ALE150 showed a maximal cell dry weight of 134.5 g/L and a lipid yield of 80.14 g/L, representing a 32.7 and 53.31 increase over the starting strain, respectively. | [131] |
| Chlorella sp. L5 | TAP medium with 500 mg/L phenol | 0.6 g/L | 31 cycles (95 days) | High concentration phenol | The upregulations of the genes according to antioxidant enzymes and carotenoids synthesis were tolerated high phenol. | [132] |
| Chlorella sp. | TAP medium | 0.6 g/L | 31 cycles (95 days) | High concentration phenol | The strain had higher phenol biodegradation rates. | [133] |
| Phaeodactylum tricornutum (CCMP-2561) | Artificial seawater added with f/2 medium without silica | 1 × 106 cells/mL | 35 cycles (nearly 252 days) | Reducing salinity | 70% salinity potentiated the algae to enhance PUFAs. | [134] |
| Picochlorum sp. BPE23 | Liquid growth medium | OD750 of 0.2 | 322 days | Supra-optimal temperature | At the optimal growth temperature of 38 °C, the biomass yield on light was 22.3% higher, and the maximal growth rate was 70.5% higher than the wild type. | [135] |
| Synechocystis sp. PCC 6803 | BG11 medium supplemented with 1.5% agar | OD730 of 0.2 | 43 cycles (303 days) | 3% NaCl | All ALE-generated strains except S3 and S7 had a significantly higher growth rate than the control strain | [136] |
| K. marxianus | YPD medium | OD600 of 0.8 | 65 days | Various temperature | The adapted K. marxianus strain accumulates glycerol and trehalose in response to lactose stress and ameliorate osmotolerance in K. marxianus cells. | [137] |
| Scenedesmus sp. SPP | Modified Chu13 medium | - | 10 days | Salinity stress, light stress, temperature stress | The triple stress-adapted strain showed the highest lipid content. | [138] |
| Schizochytrium sp. CCTCC M209059 | Main culture medium contained 40 g/L glucose and 0.4 g/L yeast extract | - | 80 days | High temperature | The adaptive strain showed a higher growth rate and lower temperature sensitivity. | [130,139] |
| Nannochloroposis oculata CCMP525 | The f/2 agar medium | 0.32 g/L | 24 cycles | High temperature | In a 2-L photobioreactor at 35 °C, biomass and lipid productivity were 1.43-fold and 2.24-fold higher, respectively, than wild type at 25 °C. | [140] |
| Crypthecodinium cohnii ATCC 30556 | Regular fermentation medium (27 g/L glucose, 25 g/L sea salt, and 6 g/L yeast extract) | OD490 of 0.1 | 280 cycles (840 days) | Varying contents of the fermentation (30–90%) supernatant | The cell growth and DHA productivity of the evolved strain (FS280) were increased by 161.87 and 311.23%. | [77] |
| Chlorella sp. AE10 | BG11 medium | 0.04 g/L | 46 cycles (138 days) | High salinity | Chlorella sp. S30, has the potential for CO2 capture under 30 g/L salt and 10% CO2 conditions. | [141] |
| Dunaliella salina CCAP 19/18 | The f/2 medium | 1 × 105 cells/ml | 5 cycles (25 days) | Blue LED | The beta-carotene concentration (33.94 ± 0.52 μM) was enhanced by 19.7% compared to that observed for the non-ALE-treated wild type of D. salina under the B-R system (28.34 ± 0.24 μM). | [142] |
| Waste Stream | Wastewater Source | Species | Reactor | Light Intensity | Ambient Temperature (°C) | Sampling Time (day) | Water Quality Index (mg/L) | Nutrient Removal Efficiency (%) | Maximum Biomass Concentration (g/L) | CO2 Bio-Fixation (mg/L/d) | Lipid Productivity (mg/L/d) | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COD * | TN * | TP * | COD | TN | TP | |||||||||||
| Agricultural wastewater | Soybean processing wastewater | Chlorella sp. L166 | Erlenmeyer flask | 6000 lux | 25 ± 1 | 18 | 5320 | 106.99 | 23.28 | 78.20 | 96.07 | 95.55 | 1.52 | - | - | [109] |
| Starch processing wastewater | Chlorella pyrenoidosa | Conical flask | 127 μmol m−2 s−1 | 25 ± 1 | 6–7 | 702.4–1026.2 | 240.3–382.7 | 22.7–40.2 | 65.99 | 83.06 | 96.97 | 1.90 | - | 30.15 | [143] | |
| Swine wastewater | MBFJNU-1 | Flask | - | - | 12 | 824.53 | 547.78 | 81.72 | - | 90.51 | 91.54 | 0.63 | - | - | [144] | |
| Agricultural wastewater and municipal wastewater | Swine wastewater 2:2 secondary treated municipal wastewater | Chlorella sorokiniana | Erlenmeyer flask | 126 μmol m−2 s−1 | 28 ± 2 | 10 | - | 337.32 | 30.88 | - | 63.90 | 93.02 | 1.31 | - | 23 | [145] |
| Swine wastewater 1:3 secondary treated municipal wastewater | Desmodesmus communis | Erlenmeyer flask | 126 μmol m−2 s−1 | 28 ± 2 | 10 | - | 188.61 | 15.77 | - | 88.02 | 99.73 | 1.02 | - | - | ||
| Municipal wastewater | Wastewater influent after primary settling tank | Chlorella sorokiniana pa.91 | Erlenmeyer flask | 4000 lux | 30 | 16 | 211.4 | 2.01 (NO3−) 0.06 (NO2−) 34.1 (NH4+) | 6.1 (PO43−) | 76 | 73 (NH4+) 93 (NO3−) | 83 (PO43−) | 3.21 | - | - | [146] |
| The primary sedimentation tank | Chlorella vulgaris ATCC 13482 | Cylindrical glass bottles | 90 ± 5 μmol m−2 s−1 | 25 | 10 | 293 | 46.67 | 19.50 | - | 93.40 | 94.10 | 0.94 | 140.91 | - | [147] | |
| Scenedesmus obliquus FACHB 417 | Cylindrical glass bottles | 90 ± 5 μmol m−2 s−1 | 25 | 10 | 293 | 46.67 | 19.50 | - | 91.50 | 91.30 | 0.87 | 129.82 | - | |||
| Effluent of anaerobically digested food wastewater | Scenedesmus bijuga | Erlenmeyer flask | 80 μmol m−2 s−1 | 30 | 28 | 5923 | 2370 | 47.80 | - | 86.60 | 90.50 | 1.49 | - | 15.59 | [148] | |
| The sewage from the sewer | Tetradesmus obliquus PF3 | Conical flask | 6000 lux | 25 ± 1 | 5 | 267 | 43 | 4.9 | 90 | 93.20 | 99 | 1.8 | 551 | - | [149] | |
| Sterilized sewage | Tetradesmus obliquus PF3 | Conical flask | 6000 lux | 25 ± 1 | 5 | 210 | 40 | 4.70 | 42 | 94.70 | 99 | 1.8 | 558 | - | ||
| Targeting the tertiary treatment of wastewater | Neochloris oleoabundans | Erlenmeyer flask | 60 μmol·m−2 s−1 | 25 | 14 | - | 42 | 18.50 | - | 100 | 31.30 | 1.17 | 145 | - | [150] | |
| Domestic wastewater from secondary settling tanks | Scenedesmus obliquus | Conical flask | 14,500 lux | 23 ± 2 | 10 | 72.16 | 12.44 | 1.08 | - | 98.90 | 97.60 | - | - | - | [151] | |
| Manure wastewater | Scenedesmus dimorphus (FACHB-496) | Erlenmeyer flask | 60–80 μmol·m−2 s−1 | 26 ± 2 | 7 | - | 306.15 | 115.08 | - | 88.16 | 73.98 | - | 638.13 | - | [152] | |
| Industrial wastewater | Artificial brewery wastewater | Scenedesmus sp. 336 | Erlenmeyer flask | 6000 lux | 25 ± 1 | 10 | 2100 | 45 | 7 | 73.66 | 75.96 | 95.71 | - | - | 38 | [153] |
| Chlorella sp. UTEX1602 | Erlenmeyer flask | 6000 lux | 25 ± 1 | 10 | 2100 | 45 | 7 | 44.97 | 81.43 | 97.54 | - | - | - | |||
| Palm oil mill effluent from an anaerobic treatment pond | Chlorella sp. (UKM2) | Transparent glass bottle | 266 μmol m−2 s−1 | 25 ± 2 | 15 | 2900 | 330 | - | - | 80.90 | - | - | 120.80 | - | [154] | |
| Waste molasses | Scenedesmus sp. Z-4 | Erlenmeyer flask | 3000 lux | 10 | 7 | 514,000 | 458 | 67 | 87.2 | 90.50 | 88.60 | 2.5 | - | 78 | [155] | |
| Membrane-treated distillery wastewater | Chlorella vulgaris | Erlenmeyer flask | 2000 lux | 25 | 9 | - | - | - | 72.24 | 80 | 94 | 0.65 | - | - | [156] | |
| Textile wastewater | Mixed microalgae (Chlorella Species and Scenedesmus sp.) | Conical flask | 212.77 mol m−2 s−1 | - | 13 | 1900 | 480.50 | 31 | 78.78 | 93.30 | 100 | 0.4 | - | - | [157] | |
| The simulated brewery effluent | Scenedesmus obliquus | Erlenmeyer flask | 12,000 lux | 30 ± 3 | 9 | 3635 | 54 | - | 57.5 | 20.80 | - | 0.9 | - | - | [158] | |
References
- Li, S.; Li, X.; Ho, S.H. How to Enhance Carbon Capture by Evolution of Microalgal Photosynthesis? Sep. Purif. Technol. 2022, 291, 120951. [Google Scholar] [CrossRef]
- Yang, S.; Yang, D.; Shi, W.; Deng, C.; Chen, C.; Feng, S. Global Evaluation of Carbon Neutrality and Peak Carbon Dioxide Emissions: Current Challenges and Future Outlook. Environ. Sci. Pollut. Res. 2022, 43, 101371. [Google Scholar] [CrossRef]
- Li, G.; Hu, R.; Hao, Y.; Yang, T.; Li, L.; Luo, Z.; Xie, L.; Zhao, N.; Liu, C.; Sun, C.; et al. CO2 and Air Pollutant Emissions from Bio-coal Briquettes. Environ. Technol. Inno. 2023, 29, 102975. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Liu, Y.; Li, N.; Zhang, Z.; Huang, C.; Chen, X.; Wang, F. The Central Trend in Crop Yields under Climate Change in China: A Systematic Review. Sci. Total Environ. 2020, 704, 135355. [Google Scholar] [CrossRef] [PubMed]
- Krūmiņš, J.; Kļaviņš, M.; Ozola-Davidāne, R.; Ansone-Bērtiņa, L. The Prospects of Clay Minerals from the Baltic States for Industrial-Scale Carbon Capture: A Review. Minerals 2022, 12, 349. [Google Scholar] [CrossRef]
- Kamyab, H.; Din, M.F.M.; Hosseini, S.E.; Ghoshal, S.K.; Ashokkumar, V.; Keyvanfar, A.; Shafaghat, A.; Lee, C.T.; asghar Bavafa, A.; Majid, M.Z.A. Optimum Lipid Production Using Agro-Industrial Wastewater Treated Microalgae as Biofuel Substrate. Clean. Technol. Environ. Policy 2016, 18, 2513–2523. [Google Scholar] [CrossRef]
- Ochedi, F.O.; Liu, D.; Yu, J.; Hussain, A.; Liu, Y. Photocatalytic, Electrocatalytic and Photoelectrocatalytic Conversion of Carbon Dioxide: A Review. Environ. Chem. Lett. 2021, 19, 941–967. [Google Scholar] [CrossRef]
- Smit, B. Carbon Capture and Storage: Introductory Lecture. Faraday Discuss. 2016, 192, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.F.A.; Dinis, M.A.P.; Lemos de Sousa, M.J. Review of European Energy Policies Regarding the Recent “Carbon Capture, Utilization and Storage” Technologies Scenario and the Role of Coal Seams. Environ. Earth Sci. 2015, 74, 2553–2561. [Google Scholar] [CrossRef]
- da Rosa, G.M.; de Morais, M.G.; Costa, J.A.V. Green Alga Cultivation with Monoethanolamine: Evaluation of CO2 Fixation and Macromolecule Production. Bioresour. Technol. 2018, 261, 206–212. [Google Scholar] [CrossRef]
- Detheridge, A.; Hosking, L.J.; Thomas, H.R.; Sarhosis, V.; Gwynn-Jones, D.; Scullion, J. Deep Seam and Minesoil Carbon Sequestration Potential of the South Wales Coalfield, UK. J. Environ. Manag. 2019, 248, 109325. [Google Scholar] [CrossRef] [PubMed]
- Salek, S.S.; Kleerebezem, R.; Jonkers, H.M.; jan Witkamp, G.; Van Loosdrecht, M.C.M. Mineral CO2 Sequestration by Environmental Biotechnological Processes. Trends Biotechnol. 2013, 31, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Yeh, J.T.; Resnik, K.P.; Rygle, K.; Pennline, H.W. Semi-Batch Absorption and Regeneration Studies for CO2 Capture by Aqueous Ammonia. Fuel Process. Technol. 2005, 86, 1533–1546. [Google Scholar] [CrossRef]
- Rossi, F.; Olguín, E.J.; Diels, L.; De Philippis, R. Microbial Fixation of CO2 in Water Bodies and in Drylands to Combat Climate Change, Soil Loss and Desertification. N. Biotechnol. 2015, 32, 109–120. [Google Scholar] [CrossRef]
- Yahya, L.; Harun, R.; Abdullah, L.C. Screening of Native Microalgae Species for Carbon Fixation at the Vicinity of Malaysian Coal-Fired Power Plant. Sci. Rep. 2020, 10, 22355. [Google Scholar] [CrossRef]
- Dasan, Y.K.; Lam, M.K.; Yusup, S.; Lim, J.W.; Show, P.L.; Tan, I.S.; Lee, K.T. Cultivation of Chlorella Vulgaris Using Sequential-Flow Bubble Column Photobioreactor: A Stress-Inducing Strategy for Lipid Accumulation and Carbon Dioxide Fixation. J. CO2 Util. 2020, 41, 101226. [Google Scholar] [CrossRef]
- Premaratne, M.; Liyanaarachchi, V.C.; Nishshanka, G.K.S.H.; Nimarshana, P.H.V.; Ariyadasa, T.U. Nitrogen-Limited Cultivation of Locally Isolated Desmodesmus sp. For Sequestration of CO2 from Simulated Cement Flue Gas and Generation of Feedstock for Biofuel Production. J. Environ. Chem. Eng. 2021, 9, 105765. [Google Scholar] [CrossRef]
- Ding, G.T.; Mohd Yasin, N.H.; Takriff, M.S.; Kamarudin, K.F.; Salihon, J.; Yaakob, Z.; Mohd Hakimi, N.I.N. Phycoremediation of Palm Oil Mill Effluent (POME) and CO2 Fixation by Locally Isolated Microalgae: Chlorella Sorokiniana UKM2, Coelastrella sp. UKM4 and Chlorella Pyrenoidosa UKM7. J. Water Process Eng. 2020, 35, 101202. [Google Scholar] [CrossRef]
- Li, G.; Hao, Y.; Yang, T.; Xiao, W.; Pan, M.; Huo, S.; Lyu, T. Enhancing Bioenergy Production from the Raw and Defatted Microalgal Biomass Using Wastewater as the Cultivation Medium. Bioengineering 2022, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.K.; Lee, K.T.; Mohamed, A.R. Current Status and Challenges on Microalgae-Based Carbon Capture. Int. J. Greenh. Gas Control 2012, 10, 456–469. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of CO2 Concentration on Algal Growth: A Review. Renew. Sustain. Energy Rev. 2014, 38, 172–179. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, J.; Pan, M.; Hao, Y.; Hu, R.; Xiao, W.; Li, G.; Lyu, T. Valorisation of Microalgae Residues after Lipid Extraction: Pyrolysis Characteristics for Biofuel Production. Biochem. Eng. J. 2022, 179, 108330. [Google Scholar] [CrossRef]
- Raeesossadati, M.J.; Ahmadzadeh, H.; McHenry, M.P.; Moheimani, N.R. CO2 Bioremediation by Microalgae in Photobioreactors: Impacts of Biomass and CO2 Concentrations, Light, and Temperature. Algal Res. 2014, 6, 78–85. [Google Scholar] [CrossRef]
- You, X.; Yang, L.; Zhou, X.; Zhang, Y. Sustainability and Carbon Neutrality Trends for Microalgae-Based Wastewater Treatment: A Review. Environ. Res. 2022, 209, 112860. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Ling, T.C.; Juan, J.C.; Lee, D.J.; Chang, J.S.; Show, P.L. Biorefineries of Carbon Dioxide: From Carbon Capture and Storage (CCS) to Bioenergies Production. Bioresour. Technol. 2016, 215, 346–356. [Google Scholar] [CrossRef]
- Burlacot, A.; Dao, O.; Auroy, P.; Cuiné, S.; Li-Beisson, Y.; Peltier, G. Alternative Photosynthesis Pathways Drive the Algal CO2-Concentrating Mechanism. Nature 2022, 605, 366–371. [Google Scholar] [CrossRef]
- Singh, S.K.; Sundaram, S.; Sinha, S.; Rahman, M.A.; Kapur, S. Recent Advances in CO2 Uptake and Fixation Mechanism of Cyanobacteria and Microalgae. Crit. Rev. Environ. Sci. Technol. 2016, 46, 1297–1323. [Google Scholar] [CrossRef]
- Prasad, R.; Gupta, S.K.; Shabnam, N.; Oliveira, C.Y.B.; Nema, A.K.; Ansari, F.A.; Bux, F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability 2021, 13, 13061. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, J.; Chen, P.; Ji, C.; Kang, Q.; Lu, B.; Li, K.; Liu, J.; Ruan, R. Bio-Mitigation of Carbon Dioxide Using Microalgal Systems: Advances and Perspectives. Renew. Sustain. Energy Rev. 2017, 76, 1163–1175. [Google Scholar] [CrossRef]
- Kong, W.; Shen, B.; Lyu, H.; Kong, J.; Ma, J.; Wang, Z.; Feng, S. Review on Carbon Dioxide Fixation Coupled with Nutrients Removal from Wastewater by Microalgae. J. Clean. Prod. 2021, 292, 125975. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Z. Advances in the Biological Fixation of Carbon Dioxide by Microalgae. J. Chem. Technol. Biotechnol. 2021, 96, 1475–1495. [Google Scholar] [CrossRef]
- Wang, Y.; Stessman, D.J.; Spalding, M.H. The CO2 Concentrating Mechanism and Photosynthetic Carbon Assimilation in Limiting CO2: How Chlamydomonas Works against the Gradient. Plant J. 2015, 82, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, D.; Goyal, A. Carbon Concentration Mechanism(s) in Unicellular Green Algae and Cyanobacteria. J. Plant Biochem. Biotechnol. 2001, 10, 83–90. [Google Scholar] [CrossRef]
- Price, G.D.; Badger, M.R.; Woodger, F.J.; Long, B.M. Advances in Understanding the Cyanobacterial CO2-Concentrating- Mechanism (CCM): Functional Components, Ci Transporters, Diversity, Genetic Regulation and Prospects for Engineering into Plants. J. Exp. Bot. 2008, 59, 1441–1461. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Qiao, L.; Zhang, H.; Zhao, D.; Su, X. The Effects of Illumination Factors on the Growth and HCO3− Fixation of Microalgae in an Experiment Culture System. Energy 2014, 78, 40–47. [Google Scholar] [CrossRef]
- Kumar, V.; Nanda, M.; Kumar, S.; Chauhan, P.K. The Effects of Ultraviolet Radiation on Growth, Biomass, Lipid Accumulation and Biodiesel Properties of Microalgae. Energy Sources Part A Recover. Util. Environ. Eff. 2018, 40, 787–793. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Sforza, E.; Gris, B.; De Farias Silva, C.E.; Morosinotto, T.; Bertucco, A. Effects of Light on Cultivation of Scenedesmus Obliquus in Batch and Continuous Flat Plate Photobioreactor. Chem. Eng. Trans. 2014, 38, 211–216. [Google Scholar]
- Kim, T.H.; Lee, Y.; Han, S.H.; Hwang, S.J. The Effects of Wavelength and Wavelength Mixing Ratios on Microalgae Growth and Nitrogen, Phosphorus Removal Using Scenedesmus sp. for Wastewater Treatment. Bioresour. Technol. 2013, 130, 75–80. [Google Scholar] [CrossRef]
- Lau, C.C.; Teh, K.Y.; Afifudeen, C.L.W.; Yee, W.; Aziz, A.; Cha, T.S. Bright as Day and Dark as Night: Light-Dependant Energy for Lipid Biosynthesis and Production in Microalgae. World J. Microbiol. Biotechnol. 2022, 38, 70. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Su, Y. Process Effect of Microalgal-Carbon Dioxide Fixation and Biomass Production: A Review. Renew. Sustain. Energy Rev. 2014, 31, 121–132. [Google Scholar] [CrossRef]
- Sachdeva, N.; Gupta, R.P.; Mathur, A.S.; Tuli, D.K. Enhanced Lipid Production in Thermo-Tolerant Mutants of Chlorella Pyrenoidosa NCIM 2738. Bioresour. Technol. 2016, 221, 576–587. [Google Scholar] [CrossRef]
- Kumar, A.; Ergas, S.; Yuan, X.; Sahu, A.; Zhang, Q.; Dewulf, J.; Malcata, F.X.; van Langenhove, H. Enhanced CO2 Fixation and Biofuel Production via Microalgae: Recent Developments and Future Directions. Trends Biotechnol. 2010, 28, 371–380. [Google Scholar] [CrossRef]
- Liang, Y.; Tang, J.; Luo, Y.; Kaczmarek, M.B.; Li, X.; Daroch, M. Thermosynechococcus as a Thermophilic Photosynthetic Microbial Cell Factory for CO2 Utilisation. Bioresour. Technol. 2019, 278, 255–265. [Google Scholar] [CrossRef]
- Ono, E.; Cuello, J.L. Carbon Dioxide Mitigation Using Thermophilic Cyanobacteria. Biosyst. Eng. 2007, 96, 129–134. [Google Scholar] [CrossRef]
- Ong, S.C.; Kao, C.Y.; Chiu, S.Y.; Tsai, M.T.; Lin, C.S. Characterization of the Thermal-Tolerant Mutants of Chlorella sp. with High Growth Rate and Application in Outdoor Photobioreactor Cultivation. Bioresour. Technol. 2010, 101, 2880–2883. [Google Scholar] [CrossRef]
- Hsueh, H.T.; Li, W.J.; Chen, H.H.; Chu, H. Carbon Bio-Fixation by Photosynthesis of Thermosynechococcus sp. CL-1 and Nannochloropsis oculta. J. Photochem. Photobiol. B Biol. 2009, 95, 33–39. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Ho, S.-H. Microalgae as a Solution of Third World Energy Crisis for Biofuels Production from Wastewater toward Carbon Neutrality: An Updated Review. Chemosphere 2022, 291, 132863. [Google Scholar] [CrossRef] [PubMed]
- Seyed Hosseini, N.; Shang, H.; Scott, J.A. Biosequestration of Industrial Off-Gas CO2 for Enhanced Lipid Productivity in Open Microalgae Cultivation Systems. Renew. Sustain. Energy Rev. 2018, 92, 458–469. [Google Scholar] [CrossRef]
- Razzak, S.A.; Ilyas, M.; Ali, S.A.M.; Hossain, M.M. Effects of CO2 Concentration and pH on Mixotrophic Growth of Nannochloropsis oculata. Appl. Biochem. Biotechnol. 2015, 176, 1290–1302. [Google Scholar] [CrossRef]
- Wang, Z.; Wen, X.; Xu, Y.; Ding, Y.; Geng, Y.; Li, Y. Maximizing CO2 Biofixation and Lipid Productivity of Oleaginous Microalga Graesiella sp. WBG-1 via CO2-Regulated pH in Indoor and Outdoor Open Reactors. Sci. Total Environ. 2018, 619–620, 827–833. [Google Scholar] [CrossRef]
- Li, S.; Song, C.; Li, M.; Chen, Y.; Lei, Z.; Zhang, Z. Effect of Different Nitrogen Ratio on the Performance of CO2 Absorption and Microalgae Conversion (CAMC) Hybrid System. Bioresour. Technol. 2020, 306, 123126. [Google Scholar] [CrossRef]
- Li, Q.; Fu, L.; Wang, Y.; Zhou, D.; Rittmann, B.E. Excessive Phosphorus Caused Inhibition and Cell Damage during Heterotrophic Growth of Chlorella regularis. Bioresour. Technol. 2018, 268, 266–270. [Google Scholar] [CrossRef]
- Razzak, S.A.; Ali, S.A.M.; Hossain, M.M.; deLasa, H. Biological CO2 Fixation with Production of Microalgae in Wastewater—A Review. Renew. Sustain. Energy Rev. 2017, 76, 379–390. [Google Scholar] [CrossRef]
- Anjos, M.; Fernandes, B.D.; Vicente, A.A.; Teixeira, J.A.; Dragone, G. Optimization of CO2 Bio-Mitigation by Chlorella vulgaris. Bioresour. Technol. 2013, 139, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Han, W.; Li, P.; Miao, X.; Zhong, J. CO2 Biofixation and Fatty Acid Composition of Scenedesmus Obliquus and Chlorella Pyrenoidosa in Response to Different CO2 Levels. Bioresour. Technol. 2011, 102, 3071–3076. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Phooi, W.B.A.; Lim, Z.J.; Tong, Y.W. Control of CO2 Input Conditions during Outdoor Culture of Chlorella Vulgaris in Bubble Column Photobioreactors. Bioresour. Technol. 2015, 186, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Moghimifam, R.; Niknam, V.; Ebrahimzadeh, H.; Hejazi, M.A. CO2 Biofixation and Fatty Acid Composition of Two Indigenous Dunaliella sp. Isolates (ABRIINW-CH2 and ABRIINW-SH33) in Response to Extremely High CO2 Levels. Bioprocess Biosyst. Eng. 2020, 43, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Hu, Z.; Liao, Q.; Huang, Y.; Zhu, X.; Ye, W.; Sun, Y. Enhancement of CO2 Transfer and Microalgae Growth by Perforated Inverted Arc Trough Internals in a Flat-Plate Photobioreactor. Bioresour. Technol. 2018, 269, 292–299. [Google Scholar] [CrossRef]
- Rodas-Zuluaga, L.I.; Castañeda-Hernández, L.; Castillo-Vacas, E.I.; Gradiz-Menjivar, A.; López-Pacheco, I.Y.; Castillo-Zacarías, C.; Boully, L.; Iqbal, H.M.N.; Parra-Saldívar, R. Bio-Capture and Influence of CO2 on the Growth Rate and Biomass Composition of the Microalgae Botryococcus Braunii and Scenedesmus sp. J. CO2 Util. 2021, 43, 101371. [Google Scholar] [CrossRef]
- Chiang, C.L.; Lee, C.M.; Chen, P.C. Utilization of the Cyanobacteria Anabaena sp. CH1 in Biological Carbon Dioxide Mitigation Processes. Bioresour. Technol. 2011, 102, 5400–5405. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.H.; Jiang, J.W.; Chen, L.P.; Cheng, L.H.; Xu, X.H.; Chen, H.L. Effect of Carbon Source on Biomass Growth and Nutrients Removal of Scenedesmus Obliquus for Wastewater Advanced Treatment and Lipid Production. Bioresour. Technol. 2015, 190, 257–263. [Google Scholar] [CrossRef]
- Liu, S.; Elvira, P.; Wang, Y.; Wang, W. Growth and Nutrient Utilization of Green Algae in Batch and Semicontinuous Autotrophic Cultivation Under High CO2 Concentration. Appl. Biochem. Biotechnol. 2019, 188, 836–853. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Qiu, Y.; Xie, M.; Liu, J.; Liu, Q.; Li, S.; Sun, L.; Wang, K.; Kansha, Y. Novel Regeneration and Utilization Concept Using Rich Chemical Absorption Solvent As a Carbon Source for Microalgae Biomass Production. Ind. Eng. Chem. Res. 2019, 58, 11720–11727. [Google Scholar] [CrossRef]
- de Morais, E.G.; Cassuriaga, A.P.A.; Callejas, N.; Martinez, N.; Vieitez, I.; Jachmanián, I.; Santos, L.O.; de Morais, M.G.; Costa, J.A.V. Evaluation of CO2 Biofixation and Biodiesel Production by Spirulina (Arthospira) Cultivated In Air-Lift Photobioreactor. Brazilian Arch. Biol. Technol. 2018, 61, 1–11. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, X.; Hu, X.; Yang, Y.; Chen, X.; Tian, J.; Pan, T.; Ding, B. Effects of Different Concentrations of CO2 on Scenedesmus Obliquus to Overcome Sludge Extract Toxicity and Accumulate Biomass. Chemosphere 2022, 305, 135514. [Google Scholar] [CrossRef]
- Sheng, Y.; Mathimani, T.; Brindhadevi, K.; Basha, S.; Elfasakhany, A.; Xia, C.; Pugazhendhi, A. Combined Effect of CO2 Concentration and Low-Cost Urea Repletion/Starvation in Chlorella Vulgaris for Ameliorating Growth Metrics, Total and Non-Polar Lipid Accumulation and Fatty Acid Composition. Sci. Total Environ. 2022, 808, 151969. [Google Scholar] [CrossRef]
- Singh, H.; Rout, S.; Das, D. Dark Fermentative Biohydrogen Production Using Pretreated Scenedesmus Obliquus Biomass under an Integrated Paradigm of Biorefinery. Int. J. Hydrogen Energy 2022, 47, 102–116. [Google Scholar] [CrossRef]
- Li, D.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving High Carbon Dioxide Tolerance and Carbon Dioxide Fixation Capability of Chlorella sp. by Adaptive Laboratory Evolution. Bioresour. Technol. 2015, 185, 269–275. [Google Scholar] [CrossRef]
- Barahoei, M.; Hatamipour, M.S.; Afsharzadeh, S. CO2 Capturing by Chlorella Vulgaris in a Bubble Column Photo-Bioreactor; Effect of Bubble Size on CO2 Removal and Growth Rate. J. CO2 Util. 2020, 37, 9–19. [Google Scholar] [CrossRef]
- Kselíková, V.; Singh, A.; Bialevich, V.; Čížková, M.; Bišová, K. Improving Microalgae for Biotechnology—From Genetics to Synthetic Biology—Moving Forward but Not There Yet. Biotechnol. Adv. 2022, 58, 107885. [Google Scholar] [CrossRef] [PubMed]
- Nojima, D.; Ishizuka, Y.; Muto, M.; Ujiro, A.; Kodama, F.; Yoshino, T.; Maeda, Y.; Matsunaga, T.; Tanaka, T. Enhancement of Biomass and Lipid Productivities of Water Surface-Floating Microalgae by Chemical Mutagenesis. Mar. Drugs 2017, 15, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanadul, O.u.m.; Noochanong, W.; Jirakranwong, P.; Chanprame, S. EMS-Induced Mutation Followed by Quizalofop-Screening Increased Lipid Productivity in Chlorella sp. Bioprocess Biosyst. Eng. 2018, 41, 613–619. [Google Scholar] [CrossRef]
- Cheng, J.; Li, K.; Yang, Z.; Zhou, J.; Cen, K. Enhancing the Growth Rate and Astaxanthin Yield of Haematococcus Pluvialis by Nuclear Irradiation and High Concentration of Carbon Dioxide Stress. Bioresour. Technol. 2016, 204, 49–54. [Google Scholar] [CrossRef]
- Smalley, T.; Fields, F.J.; Berndt, A.J.E.; Ostrand, J.T.; Heredia, V.; Mayfield, S.P. Improving Biomass and Lipid Yields of Desmodesmus Armatus and Chlorella Vulgaris through Mutagenesis and High-Throughput Screening. Biomass Bioenergy 2020, 142, 105755. [Google Scholar] [CrossRef]
- Liu, L.; Diao, J.; Bi, Y.; Zeng, L.; Wang, F.; Chen, L.; Zhang, W. Rewiring the Metabolic Network to Increase Docosahexaenoic Acid Productivity in Crypthecodinium Cohnii by Fermentation Supernatant-Based Adaptive Laboratory Evolution. Front. Microbiol. 2022, 13, 824189. [Google Scholar] [CrossRef]
- Wei, L.; Wang, Q.; Xin, Y.; Lu, Y.; Xu, J. Enhancing Photosynthetic Biomass Productivity of Industrial Oleaginous Microalgae by Overexpression of RuBisCO Activase. Algal Res. 2017, 27, 366–375. [Google Scholar] [CrossRef]
- Baek, K.; Kim, D.H.; Jeong, J.; Sim, S.J.; Melis, A.; Kim, J.S.; Jin, E.; Bae, S. DNA-Free Two-Gene Knockout in Chlamydomonas Reinhardtii via CRISPR-Cas9 Ribonucleoproteins. Sci. Rep. 2016, 6, 30620. [Google Scholar] [CrossRef]
- Cheng, J.; Zhu, Y.; Zhang, Z.; Yang, W. Modification and Improvement of Microalgae Strains for Strengthening CO2 Fixation from Coal-Fired Flue Gas in Power Plants. Bioresour. Technol. 2019, 291, 121850. [Google Scholar] [CrossRef] [PubMed]
- Tharek, A.; Yahya, A.; Salleh, M.M.; Jamaluddin, H.; Yoshizaki, S.; Hara, H.; Iwamoto, K.; Suzuki, I.; Mohamad, S.E. Improvement and Screening of Astaxanthin Producing Mutants of Newly Isolated Coelastrum sp. Using Ethyl Methane Sulfonate Induced Mutagenesis Technique. Biotechnol. Rep. 2021, 32, e00673. [Google Scholar] [CrossRef] [PubMed]
- Kuo, E.Y.; Yang, R.; Chin, Y.Y.; Chien, Y.; Chen, Y.C.; Wei, C.; Kao, L.; Chang, Y.; Li, Y.; Chen, T.; et al. Multiomics Approaches and Genetic Engineering of Metabolism for Improved Biorefinery and Wastewater Treatment in Microalgae. Biotechnol. J. 2022, 17, e2100603. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.G.; Alcantar-Rivera, B.; Meléndez-Sánchez, E.R.; Martínez-Prado, M.A.; Sosa-Hernández, J.E.; Iqbal, H.M.N.; Parra-Saldivar, R.; Martínez-Ruiz, M. Effects of UV and UV-Vis Irradiation on the Production of Microalgae and Macroalgae: New Alternatives to Produce Photobioprotectors and Biomedical Compounds. Molecules 2022, 27, 5334. [Google Scholar] [CrossRef] [PubMed]
- Moha-León, J.D.; Pérez-Legaspi, I.A.; Ortega-Clemente, L.A.; Rubio-Franchini, I.; Ríos-Leal, E. Improving the Lipid Content of Nannochloropsis Oculata by a Mutation-Selection Program Using UV Radiation and Quizalofop. J. Appl. Phycol. 2019, 31, 191–199. [Google Scholar] [CrossRef]
- Cheng, J.; Huang, Y.; Feng, J.; Sun, J.; Zhou, J.; Cen, K. Mutate Chlorella sp. by Nuclear Irradiation to Fix High Concentrations of CO2. Bioresour. Technol. 2013, 136, 496–501. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, J.; Meng, F. Adaptive Laboratory Evolution of Microalgae: A Review of the Regulation of Growth, Stress Resistance, Metabolic Processes, and Biodegradation of Pollutants. Front. Microbiol. 2021, 12, 737248. [Google Scholar] [CrossRef]
- Arora, N.; Philippidis, G.P. Microalgae Strain Improvement Strategies: Random Mutagenesis and Adaptive Laboratory Evolution. Trends Plant Sci. 2021, 26, 1199–1200. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Wu, Y.; Fan, Y.; Zhu, C.; Fu, X.; Chu, Y.; Chen, F.; Sun, H.; Mou, H. Application of Microalgal Stress Responses in Industrial Microalgal Production Systems. Mar. Drugs 2022, 20, 30. [Google Scholar] [CrossRef]
- Zhao, Q.; Huang, H. Adaptive Evolution Improves Algal Strains for Environmental Remediation. Trends Biotechnol. 2021, 39, 112–115. [Google Scholar] [CrossRef]
- Sun, X.M.; Ren, L.J.; Ji, X.J.; Chen, S.L.; Guo, D.S.; Huang, H. Adaptive Evolution of Schizochytrium sp. by Continuous High Oxygen Stimulations to Enhance Docosahexaenoic Acid Synthesis. Bioresour. Technol. 2016, 211, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Åhlén, M.; Cheung, O.; Xu, C. Low-Concentration CO2 Capture Using Metal–Organic Frameworks—Current Status and Future Perspectives. Dalt. Trans. 2023, 52, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.M.; Mechery, J.; Paulose, S.V. Carbon Dioxide Capture Strategies from Flue Gas Using Microalgae: A Review. Environ. Sci. Pollut. Res. 2016, 23, 16926–16940. [Google Scholar] [CrossRef] [PubMed]
- Tsai, D.D.W.; Chen, P.H.; jung Chou, C.M.; Hsu, C.F.; Ramaraj, R. Carbon Sequestration by Alga Ecosystems. Ecol. Eng. 2015, 84, 386–389. [Google Scholar] [CrossRef]
- Mondal, M.; Goswami, S.; Ghosh, A.; Oinam, G.; Tiwari, O.N.; Das, P.; Gayen, K.; Mandal, M.K.; Halder, G.N. Production of Biodiesel from Microalgae through Biological Carbon Capture: A Review. 3 Biotech 2017, 7, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 Bio-Mitigation Using Microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef]
- Suparmaniam, U.; Lam, M.K.; Uemura, Y.; Lim, J.W.; Lee, K.T.; Shuit, S.H. Insights into the Microalgae Cultivation Technology and Harvesting Process for Biofuel Production: A Review. Renew. Sustain. Energy Rev. 2019, 115, 109361. [Google Scholar] [CrossRef]
- Sen Tan, J.; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.-H.; Show, P.L. A Review on Microalgae Cultivation and Harvesting, and Their Biomass Extraction Processing Using Ionic Liquids. Bioengineered 2020, 11, 116–129. [Google Scholar] [CrossRef] [Green Version]
- McGinn, P.J.; Dickinson, K.E.; Bhatti, S.; Frigon, J.C.; Guiot, S.R.; O’Leary, S.J.B. Integration of Microalgae Cultivation with Industrial Waste Remediation for Biofuel and Bioenergy Production: Opportunities and Limitations. Photosynth. Res. 2011, 109, 231–247. [Google Scholar] [CrossRef]
- Kumar, S.; Aswar, D. Recent Advances in Thin Films; Materials Horizons: From Nature to Nanomaterials; Springer: Singapore, 2020; ISBN 978-981-15-6116-0. [Google Scholar]
- Van Den Hende, S.; Vervaeren, H.; Boon, N. Flue Gas Compounds and Microalgae: (Bio-)Chemical Interactions Leading to Biotechnological Opportunities. Biotechnol. Adv. 2012, 30, 1405–1424. [Google Scholar] [CrossRef] [PubMed]
- Mallapragada, D.S.; Singh, N.R.; Curteanu, V.; Agrawal, R. Sun-to-Fuel Assessment of Routes for Fixing CO2 as Liquid Fuel. Ind. Eng. Chem. Res. 2013, 52, 5136–5144. [Google Scholar] [CrossRef]
- Vuppaladadiyam, A.K.; Yao, J.G.; Florin, N.; George, A.; Wang, X.; Labeeuw, L.; Jiang, Y.; Davis, R.W.; Abbas, A.; Ralph, P.; et al. Impact of Flue Gas Compounds on Microalgae and Mechanisms for Carbon Assimilation and Utilization. ChemSusChem 2018, 11, 334–355. [Google Scholar] [CrossRef]
- Yen, H.W.; Ho, S.H.; Chen, C.Y.; Chang, J.S. CO2, NOx and SOx Removal from Flue Gas via Microalgae Cultivation: A Critical Review. Biotechnol. J. 2015, 10, 829–839. [Google Scholar] [CrossRef]
- Duarte, J.H.; de Morais, E.G.; Radmann, E.M.; Costa, J.A.V. Biological CO2 Mitigation from Coal Power Plant by Chlorella Fusca and Spirulina sp. Bioresour. Technol. 2017, 234, 472–475. [Google Scholar] [CrossRef]
- Molazadeh, M.; Ahmadzadeh, H.; Pourianfar, H.R.; Lyon, S.; Rampelotto, P.H. The Use of Microalgae for Coupling Wastewater Treatment With CO2 Biofixation. Front. Bioeng. Biotechnol. 2019, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Yang, T.; Xiao, W.; Wu, J.; Xu, F.; Li, L.; Gao, F.; Huang, Z. Sustainable Environmental Assessment of Waste-to-Energy Practices: Co-Pyrolysis of Food Waste and Discarded Meal Boxes. Foods 2022, 11, 3840. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Han, X.; Qiu, Y.; Liu, Z.; Li, S.; Kitamura, Y. Microalgae Carbon Fixation Integrated with Organic Matters Recycling from Soybean Wastewater: Effect of pH on the Performance of Hybrid System. Chemosphere 2020, 248, 126094. [Google Scholar] [CrossRef] [PubMed]
- Omar Faruque, M.; Ilyas, M.; Mozahar Hossain, M.; Abdur Razzak, S. Influence of Nitrogen to Phosphorus Ratio and CO2 Concentration on Lipids Accumulation of Scenedesmus Dimorphus for Bioenergy Production and CO2 Biofixation. Chem. Asian J. 2020, 15, 4307–4320. [Google Scholar] [CrossRef]
- Hu, X.; Song, C.; Mu, H.; Liu, Z.; Kitamura, Y. Optimization of Simultaneous Soybean Processing Wastewater Treatment and Flue Gas CO2 Fixation via Chlorella sp. L166 Cultivation. J. Environ. Chem. Eng. 2020, 8, 103960. [Google Scholar] [CrossRef]
- Li, G.; Hu, R.; Wang, N.; Yang, T.; Xu, F.; Li, J.; Wu, J.; Huang, Z.; Pan, M.; Lyu, T. Cultivation of Microalgae in Adjusted Wastewater to Enhance Biofuel Production and Reduce Environmental Impact: Pyrolysis Performances and Life Cycle Assessment. J. Clean. Prod. 2022, 355, 131768. [Google Scholar] [CrossRef]
- Ahmad, A.L.; Yasin, N.H.M.; Derek, C.J.C.; Lim, J.K. Microalgae as a Sustainable Energy Source for Biodiesel Production: A Review. Renew. Sustain. Energy Rev. 2011, 15, 584–593. [Google Scholar] [CrossRef]
- Villaro, S.; Ciardi, M.; Morillas-españa, A.; Sánchez-Zurano, A.; Acién-Fernández, G.; Lafarga, T. Trends and Industrial Use as Food. Foods 2021, 10, 2303. [Google Scholar] [CrossRef] [PubMed]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.C.; Liang, M.H.; Jiang, J.G. Transgenic Microalgae as Bioreactors. Crit. Rev. Food Sci. Nutr. 2020, 60, 3195–3213. [Google Scholar] [CrossRef]
- Muñoz, C.F.; Südfeld, C.; Naduthodi, M.I.S.; Weusthuis, R.A.; Barbosa, M.J.; Wijffels, R.H.; D’Adamo, S. Genetic Engineering of Microalgae for Enhanced Lipid Production. Biotechnol. Adv. 2021, 52, 107836. [Google Scholar] [CrossRef]
- Mavrommati, M.; Daskalaki, A.; Papanikolaou, S.; Aggelis, G. Adaptive Laboratory Evolution Principles and Applications in Industrial Biotechnology. Biotechnol. Adv. 2022, 54, 107795. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, R.; Takriff, M.S.; Veeramuthu, A. Technical Insights into Carbon Dioxide Sequestration by Microalgae: A Biorefinery Approach towards Sustainable Environment. Biomass Convers. Bioref. 2022. [Google Scholar] [CrossRef]
- Marchão, L.; Fernandes, J.R.; Sampaio, A.; Peres, J.A.; Tavares, P.B.; Lucas, M.S. Microalgae and Immobilized TiO2/UV-A LEDs as a Sustainable Alternative for Winery Wastewater Treatment. Water Res. 2021, 203, 117464. [Google Scholar] [CrossRef]
- Huang, G.; Chen, F.; Kuang, Y.; He, H.; Qin, A. Current Techniques of Growing Algae Using Flue Gas from Exhaust Gas Industry: A Review. Appl. Biochem. Biotechnol. 2016, 178, 1220–1238. [Google Scholar] [CrossRef]
- Chia, S.R.; Chew, K.W.; Leong, H.Y.; Ho, S.H.; Munawaroh, H.S.H.; Show, P.L. CO2 Mitigation and Phycoremediation of Industrial Flue Gas and Wastewater via Microalgae-Bacteria Consortium: Possibilities and Challenges. Chem. Eng. J. 2021, 425, 131436. [Google Scholar] [CrossRef]
- Ma, Z.; Cheah, W.Y.; Ng, I.S.; Chang, J.S.; Zhao, M.; Show, P.L. Microalgae-Based Biotechnological Sequestration of Carbon Dioxide for Net Zero Emissions. Trends Biotechnol. 2022, 40, 1439–1453. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from Microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Liang, M.H.; Wang, L.; Wang, Q.; Zhu, J.; Jiang, J.G. High-Value Bioproducts from Microalgae: Strategies and Progress. Crit. Rev. Food Sci. Nutr. 2019, 59, 2423–2441. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Nguyen, T.H.T.; Jin, E.S. Improving Lipid Production by Strain Development in Microalgae: Strategies, Challenges and Perspectives. Bioresour. Technol. 2019, 292, 121953. [Google Scholar] [CrossRef] [PubMed]
- Mohsenpour, S.F.; Hennige, S.; Willoughby, N.; Adeloye, A.; Gutierrez, T. Integrating Micro-Algae into Wastewater Treatment: A Review. Sci. Total Environ. 2021, 752, 142168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; An, S.; Yao, R.; Fu, W.; Han, Y.; Du, M.; Chen, Z.; Lei, A.; Wang, J. Life Cycle Assessment of Auto-Tropically Cultivated Economic Microalgae for Final Products Such as Food, Total Fatty Acids, and Bio-Oil. Front. Mar. Sci. 2022, 9, 990635. [Google Scholar] [CrossRef]
- Last, G.V.; Schmick, M.T. A Review of Major Non-Power-Related Carbon Dioxide Stream Compositions. Environ. Earth Sci. 2015, 74, 1189–1198. [Google Scholar] [CrossRef]
- De Coninck, H.; Benson, S.M. Carbon Dioxide Capture and Storage: Issues and Prospects. Annu. Rev. Environ. Resour. 2014, 39, 243–270. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Q.; Miao, X.; Shi, J. Enhancement of Lipid Production in Low-Starch Mutants Chlamydomonas Reinhardtii by Adaptive Laboratory Evolution. Bioresour. Technol. 2013, 147, 499–507. [Google Scholar] [CrossRef]
- Ren, L.J.; Huang, H.; Xiao, A.H.; Lian, M.; Jin, L.J.; Ji, X.J. Enhanced Docosahexaenoic Acid Production by Reinforcing Acetyl-CoA and NADPH Supply in Schizochytrium sp. HX-308. Bioprocess Biosyst. Eng. 2009, 32, 837–843. [Google Scholar] [CrossRef]
- Sun, X.M.; Ren, L.J.; Bi, Z.Q.; Ji, X.J.; Zhao, Q.Y.; Huang, H. Adaptive Evolution of Microalgae Schizochytrium sp. under High Salinity Stress to Alleviate Oxidative Damage and Improve Lipid Biosynthesis. Bioresour. Technol. 2018, 267, 438–444. [Google Scholar] [CrossRef]
- Zhou, L.; Cheng, D.; Wang, L.; Gao, J.; Zhao, Q.; Wei, W.; Sun, Y. Comparative Transcriptomic Analysis Reveals Phenol Tolerance Mechanism of Evolved Chlorella Strain. Bioresour. Technol. 2017, 227, 266–272. [Google Scholar] [CrossRef]
- Wang, L.; Xue, C.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Strain Improvement of Chlorella sp. for Phenol Biodegradation by Adaptive Laboratory Evolution. Bioresour. Technol. 2016, 205, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Luo, S.W.; Luo, W.; Yang, W.D.; Liu, J.S.; Li, H.Y. Adaptive Evolution of Microalgal Strains Empowered by Fulvic Acid for Enhanced Polyunsaturated Fatty Acid Production. Bioresour. Technol. 2019, 277, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Barten, R.; van Workum, D.J.M.; de Bakker, E.; Risse, J.; Kleisman, M.; Navalho, S.; Smit, S.; Wijffels, R.H.; Nijveen, H.; Barbosa, M.J. Genetic Mechanisms Underlying Increased Microalgal Thermotolerance, Maximal Growth Rate, and Yield on Light Following Adaptive Laboratory Evolution. BMC Biol. 2022, 20, 242. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; He, J.; Dong, M.; Tang, X.; Jiang, P.; Lei, A.; Wang, J. Divergent Metabolic and Transcriptomic Responses of Synechocystis sp. PCC 6803 to Salt Stress after Adaptive Laboratory Evolution. Algal Res. 2020, 47, 101856. [Google Scholar] [CrossRef]
- Saini, P.; Beniwal, A.; Kokkiligadda, A.; Vij, S. Evolutionary Adaptation of Kluyveromyces Marxianus Strain for Efficient Conversion of Whey Lactose to Bioethanol. Process Biochem. 2017, 62, 69–79. [Google Scholar] [CrossRef]
- Maneechote, W.; Cheirsilp, B. Stepwise-Incremental Physicochemical Factors Induced Acclimation and Tolerance in Oleaginous Microalgae to Crucial Outdoor Stresses and Improved Properties as Biodiesel Feedstocks. Bioresour. Technol. 2021, 328, 124850. [Google Scholar] [CrossRef]
- Hu, X.; Tang, X.; Bi, Z.; Zhao, Q.; Ren, L. Adaptive Evolution of Microalgae Schizochytrium sp. under High Temperature for Efficient Production of Docosahexaeonic Acid. Algal Res. 2021, 54, 102212. [Google Scholar] [CrossRef]
- Arora, N.; Lo, E.; Philippidis, G.P. A Two-Prong Mutagenesis and Adaptive Evolution Strategy to Enhance the Temperature Tolerance and Productivity of Nannochloropsis oculata. Bioresour. Technol. 2022, 364, 128101. [Google Scholar] [CrossRef]
- Li, X.; Yuan, Y.; Cheng, D.; Gao, J.; Kong, L.; Zhao, Q.; Wei, W.; Sun, Y. Exploring Stress Tolerance Mechanism of Evolved Freshwater Strain Chlorella sp. S30 under 30 g/L Salt. Bioresour. Technol. 2018, 250, 495–504. [Google Scholar] [CrossRef]
- Han, S.I.; Kim, S.; Lee, C.; Choi, Y.E. Blue-Red LED Wavelength Shifting Strategy for Enhancing Beta-Carotene Production from Halotolerant Microalga, Dunaliella salina. J. Microbiol. 2019, 57, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Chu, H.; Zhang, Y.; Yang, L.; Zhao, F.; Zhou, X. Chlorella Pyrenoidosa Cultivation Using Anaerobic Digested Starch Processing Wastewater in an Airlift Circulation Photobioreactor. Bioresour. Technol. 2014, 170, 538–548. [Google Scholar] [CrossRef]
- Wen, Y.; He, Y.; Ji, X.; Li, S.; Chen, L.; Zhou, Y.; Wang, M.; Chen, B. Isolation of an Indigenous Chlorella Vulgaris from Swine Wastewater and Characterization of Its Nutrient Removal Ability in Undiluted Sewage. Bioresour. Technol. 2017, 243, 247–253. [Google Scholar] [CrossRef]
- Yao, L.; Shi, J.; Miao, X. Mixed Wastewater Coupled with CO2 for Microalgae Culturing and Nutrient Removal. PLoS ONE 2015, 10, e0139117. [Google Scholar] [CrossRef] [Green Version]
- Taghavijeloudar, M.; Yaqoubnejad, P.; Amini-Rad, H.; Park, J. Optimization of Cultivation Condition of Newly Isolated Strain Chlorella Sorokiniana Pa.91 for CO2 Bio-Fixation and Nutrients Removal from Wastewater: Impact of Temperature and Light Intensity. Clean Technol. Environ. Policy 2021, 25, 589–601. [Google Scholar] [CrossRef]
- Chaudhary, R.; Dikshit, A.K.; Tong, Y.W. Carbon-Dioxide Biofixation and Phycoremediation of Municipal Wastewater Using Chlorella Vulgaris and Scenedesmus obliquus. Environ. Sci. Pollut. Res. 2018, 25, 20399–20406. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.Y.; Cho, H.U.; Utomo, J.C.; Choi, Y.N.; Xu, X.; Park, J.M. Biodiesel Production from Scenedesmus Bijuga Grown in Anaerobically Digested Food Wastewater Effluent. Bioresour. Technol. 2015, 184, 215–221. [Google Scholar] [CrossRef]
- Ma, S.; Yu, Y.; Cui, H.; Yadav, R.S.; Li, J.; Feng, Y. Unsterilized Sewage Treatment and Carbohydrate Accumulation in Tetradesmus obliquus PF3 with CO2 Supplementation. Algal Res. 2020, 45, 101741. [Google Scholar] [CrossRef]
- Razzak, S.A. In Situ Biological CO2 Fixation and Wastewater Nutrient Removal with Neochloris Oleoabundans in Batch Photobioreactor. Bioprocess Biosyst. Eng. 2019, 42, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Jin, W.; Li, Z.; Wei, Y.; He, Z.; Chen, C.; Qin, C.; Chen, Y.; Tu, R.; Zhou, X. Cultivation of Microalgae for Lipid Production Using Municipal Wastewater. Process Saf. Environ. Prot. 2021, 155, 155–165. [Google Scholar] [CrossRef]
- Xu, X.; Shen, Y.; Chen, J. Cultivation of Scenedesmus Dimorphus for C/N/P Removal and Lipid Production. Electron. J. Biotechnol. 2015, 18, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Hu, X.; Liu, Z.; Li, S.; Kitamura, Y. Combination of Brewery Wastewater Purification and CO2 Fixation with Potential Value-Added Ingredients Production via Different Microalgae Strains Cultivation. J. Clean. Prod. 2020, 268, 122332. [Google Scholar] [CrossRef]
- Hariz, H.B.; Takriff, M.S.; Ba-Abbad, M.M.; Mohd Yasin, N.H.; Mohd Hakim, N.I.N. CO2 Fixation Capability of Chlorella sp. and Its Use in Treating Agricultural Wastewater. J. Appl. Phycol. 2018, 30, 3017–3027. [Google Scholar] [CrossRef]
- Ma, C.; Wen, H.; Xing, D.; Pei, X.; Zhu, J.; Ren, N.; Liu, B. Molasses Wastewater Treatment and Lipid Production at Low Temperature Conditions by a Microalgal Mutant Scenedesmus sp. Z-4. Biotechnol. Biofuels 2017, 10, 111. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Amenorfenyo, D.K.; Zhang, Y.; Zhang, N.; Li, C.; Huang, X. Cultivation of Chlorella Vulgaris in Membrane-Treated Industrial Distillery Wastewater: Growth and Wastewater Treatment. Front. Environ. Sci. 2021, 9, 770633. [Google Scholar] [CrossRef]
- Huy, M.; Kumar, G.; Kim, H.W.; Kim, S.H. Photoautotrophic Cultivation of Mixed Microalgae Consortia Using Various Organic Waste Streams towards Remediation and Resource Recovery. Bioresour. Technol. 2018, 247, 576–581. [Google Scholar] [CrossRef]
- Mata, T.M.; Melo, A.C.; Simões, M.; Caetano, N.S. Parametric Study of a Brewery Effluent Treatment by Microalgae Scenedesmus obliquus. Bioresour. Technol. 2012, 107, 151–158. [Google Scholar] [CrossRef]







| Species | Culture Medium | Optimal Growth Temperature (°C) | Average Specific Growth (d−1) | Ref. |
|---|---|---|---|---|
| Chlorella pyrenoidosa M18 | BG11 | 37 | 0.70 | [43] |
| Thermosynechococcus elongatus PKUAC-SCTE542 | BG11 | 55 | 0.22 | [45] |
| Chlorogleopsis sp. | BG11 | 50 | 0.14 | [46] |
| Chlorella sp. MT-15 | Artificial sea water | 30 | Approximately 0.80 | [47] |
| Chlorella sp. MT-7 | Artificial sea water | 30 | Approximately 0.60 | |
| Thermosynechococcus sp. CL-1 | Modified Fitzgerald | 50 | 2.70 | [48] |
| Nannochloropsis sp. Oculta | Modified Fitzgerald | 30 | 1.60 |
| Species | CO2 (%) | Culture Medium | Aeration Rate (vvm) | CO2 Fixation Rate (mg/L/d) | Biomass Yield (g/L) | Biomass Productivity (mg/L/d) | Ref. |
|---|---|---|---|---|---|---|---|
| Chlorella vulgaris | 2% constant | Liquid medium of 3N-BBM+V | 0.4 | 4110 | 2.59 | 3530 | [58] |
| 4% provided intermittently | 4500 | 2.62 | 3410 | ||||
| Spirulina platensis | 5 | Zarrouk medium | 0.1 | 178.46 | 1.75 | - | [43] |
| Dunaliella sp. ABRIINW-SH33 | 10 | Modified Johnson medium | 3 | 455.74 | 2.98 | 248.60 | [59] |
| 20 | 317.32 | 2.08 | 173.08 | ||||
| 30 | 279.74 | 1.83 | 152.60 | ||||
| Dunaliella sp. ABRIINW-CH2 | 10 | 423.19 | 2.77 | 230.83 | |||
| 20 | 317.32 | 2.08 | 173.08 | ||||
| 30 | 272.56 | 1.78 | 148.67 | ||||
| Chlorella vulgaris FACHB-31 | 15 | Modified BG11 | 0.02 | 878.40 | 3.35 | - | [60] |
| Botryococcus braunii | 0.03 | BG11 | 0.1 | - | 0.64 | - | [61] |
| 10 | 0.41 | ||||||
| 20 | 0.26 | ||||||
| Scenedesmus sp. | 0.03 | BG11 | 0.1 | - | 0.72 | - | |
| 10 | 0.90 | ||||||
| 20 | 1.90 | ||||||
| Anabaena sp. CH1 | 10 | Arnon medium | 0.4 | 1010 | 1.16 | - | [62] |
| Scenedesmus obliquus | 5 | Selenite enrichment medium | - | 577.60 | - | - | [63] |
| Chlorella protothecoides | 20 | BG11 | - | 370 | 1.55 | 190 | [64] |
| Chlorella vulgaris P12 | 6.50 | - | 0.5 | 2290 | 9.97 | 1330 | [56] |
| Chlorella sp. L38 | 5 | BG11 | - | - | 0.60 | - | [65] |
| Spirulina sp. LEB 18 | 10 | Zarrouk medium | - | 160 | 1.07 | 20 | [66] |
| Scenedesmus obliquus (FACHB-13) | 15 | BG11 | 1 | - | 1.61 | - | [67] |
| Chlorella vulgaris | 15 | BG11 | - | 120 | 1.83 | 144 | [68] |
| Scenedesmus obliquus UTEX 393 | 5 | Airlift photobioreactor | 0.43 | 727.70 | - | 405.70 | [69] |
| Chlorella sp. AE 10 | 30% | BG11 | - | - | 3.68 | - | [70] |
| Strategies | Organism | Phenotype | Ref. | |
|---|---|---|---|---|
| Random mutagenesis | Chemical mutagenesis | Chlorococcum sp. FFG039 | The FFG039 PM exhibited 1.7-fold and 1.9-fold higher biomass and lipid productivities than those of the wild type. | [73] |
| Chlorella sp. | The E100-30-60 showed that the highest biomass yield and biomass productivity were 111 and 110% higher than the wild type, respectively. | [74] | ||
| Physical mutagenesis | Haematococcus pluvialis | The average specific growth rate of Haematococcus pluvialis mutated with 4000 Gy γ-ray irradiation was increased by 15% compared with the original strain with air aeration. | [75] | |
| Chlorella vulgaris | The resulting mutant resulted in a 33% increase in lipid yield. | [76] | ||
| ALE | Chlorella sp. | The maximal biomass concentration of AE10 was 3.68 ± 0.08 g/L in 30% CO2, which was 2.94 fold compared to the original strain. | [70] | |
| Crypthecodinium cohnii ATCC 30556 | The cell growth of the evolved strain (FS280) was increased by 161.87%. | [77] | ||
| Genetic engineering | Nannochloropsis oceanica | The growth rate of mutants was enhanced by 32%, and biomass accumulation by 46%. | [78] | |
| Chlamydomonas reinhardtii | The strain improved photosynthetic productivity. | [79] | ||
| Category | Method | Description | Advantages | Limitations | Ref. |
|---|---|---|---|---|---|
| Physical | Geologic injection | Separate and capture CO2, transport it to a storage location, and inject it deep underground for long-term isolation from the atmosphere |
|
| [127] |
| Oceanic injection | Injection of CO2 into deep ocean |
|
| [128] | |
| Chemical | Chemical absorption | Chemical absorption and desorption concept, determined by solubility of CO2 |
|
| [26] |
| Mineral carbonation | CO2 reacts with calcium- or magnesium-bearing rocks to form magnesite or calcite |
|
| [128] | |
| Biological | Forest planting | Absorption of CO2 through the photosynthesis of the trees |
|
| [30] |
| Microalgae carbon fixation | Carbon sequestration by microalgal photosynthesis |
|
| [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Xiao, W.; Yang, T.; Lyu, T. Optimization and Process Effect for Microalgae Carbon Dioxide Fixation Technology Applications Based on Carbon Capture: A Comprehensive Review. C 2023, 9, 35. https://doi.org/10.3390/c9010035
Li G, Xiao W, Yang T, Lyu T. Optimization and Process Effect for Microalgae Carbon Dioxide Fixation Technology Applications Based on Carbon Capture: A Comprehensive Review. C. 2023; 9(1):35. https://doi.org/10.3390/c9010035
Chicago/Turabian StyleLi, Gang, Wenbo Xiao, Tenglun Yang, and Tao Lyu. 2023. "Optimization and Process Effect for Microalgae Carbon Dioxide Fixation Technology Applications Based on Carbon Capture: A Comprehensive Review" C 9, no. 1: 35. https://doi.org/10.3390/c9010035
APA StyleLi, G., Xiao, W., Yang, T., & Lyu, T. (2023). Optimization and Process Effect for Microalgae Carbon Dioxide Fixation Technology Applications Based on Carbon Capture: A Comprehensive Review. C, 9(1), 35. https://doi.org/10.3390/c9010035