
Efficient Production of Succinic Acid from Sugarcane Bagasse Hydrolysate by Actinobacillus succinogenes GXAS137

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Pretreatment
2.3. Enzymatic Hydrolysis
2.4. Orthogonal Design
2.5. Microorganism and Culture Conditions
2.6. Fermentation in Anaerobic Bottles
2.7. Fermentation in a Stirred Bioreactor
2.8. Analytical methods
2.9. Statistical Design
3. Results and Discussion
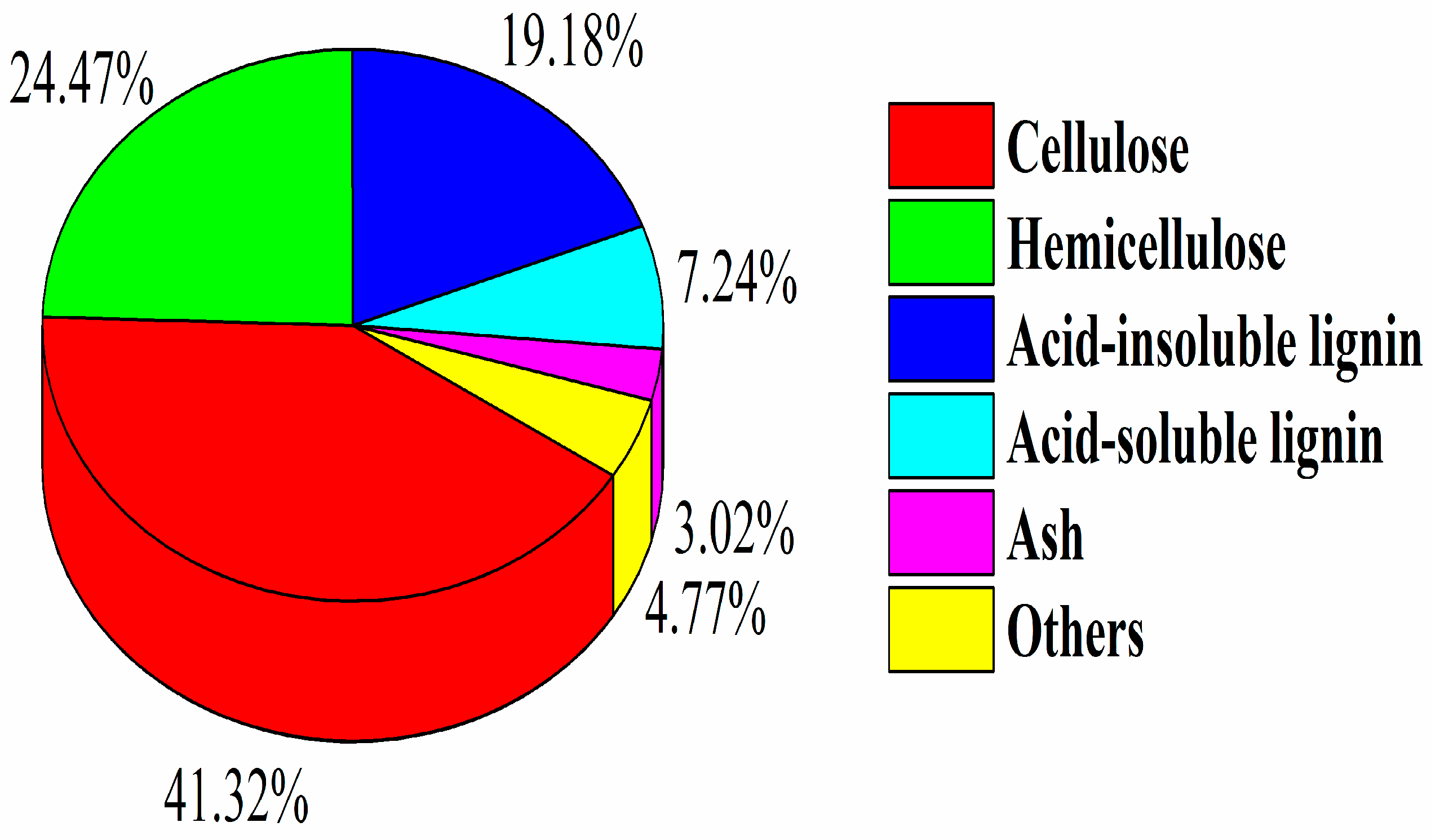
3.1. Composition Analysis of SCB
3.2. Effects of Different Pretreatment Methods on the Composition and Enzymatic Hydrolysis of SCB
3.3. Enzymatic Hydrolysis of Pretreated SCB Using Multiple Enzymes
3.4. Effect of Initial Reducing Sugar Concentration on SA Production
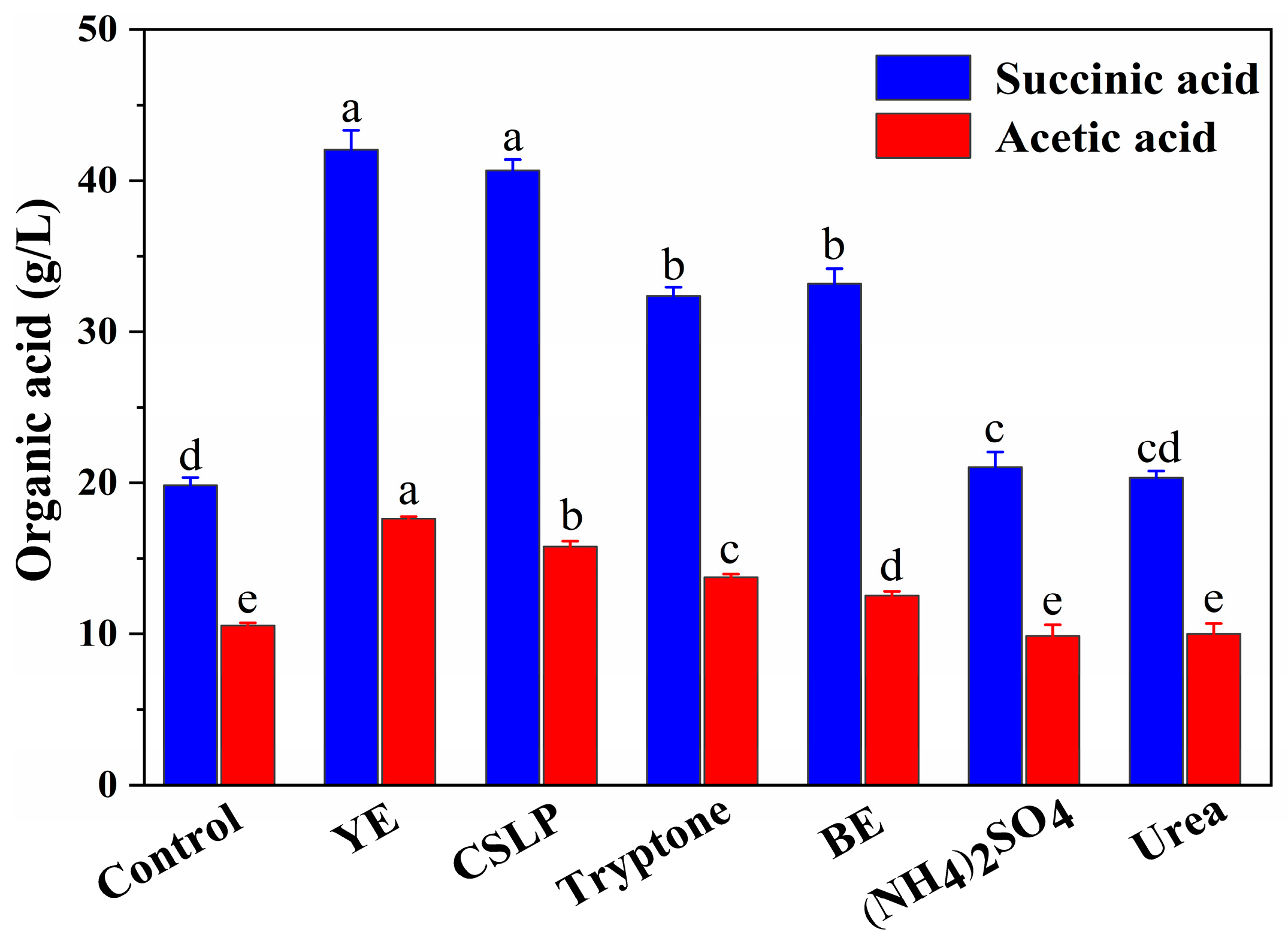
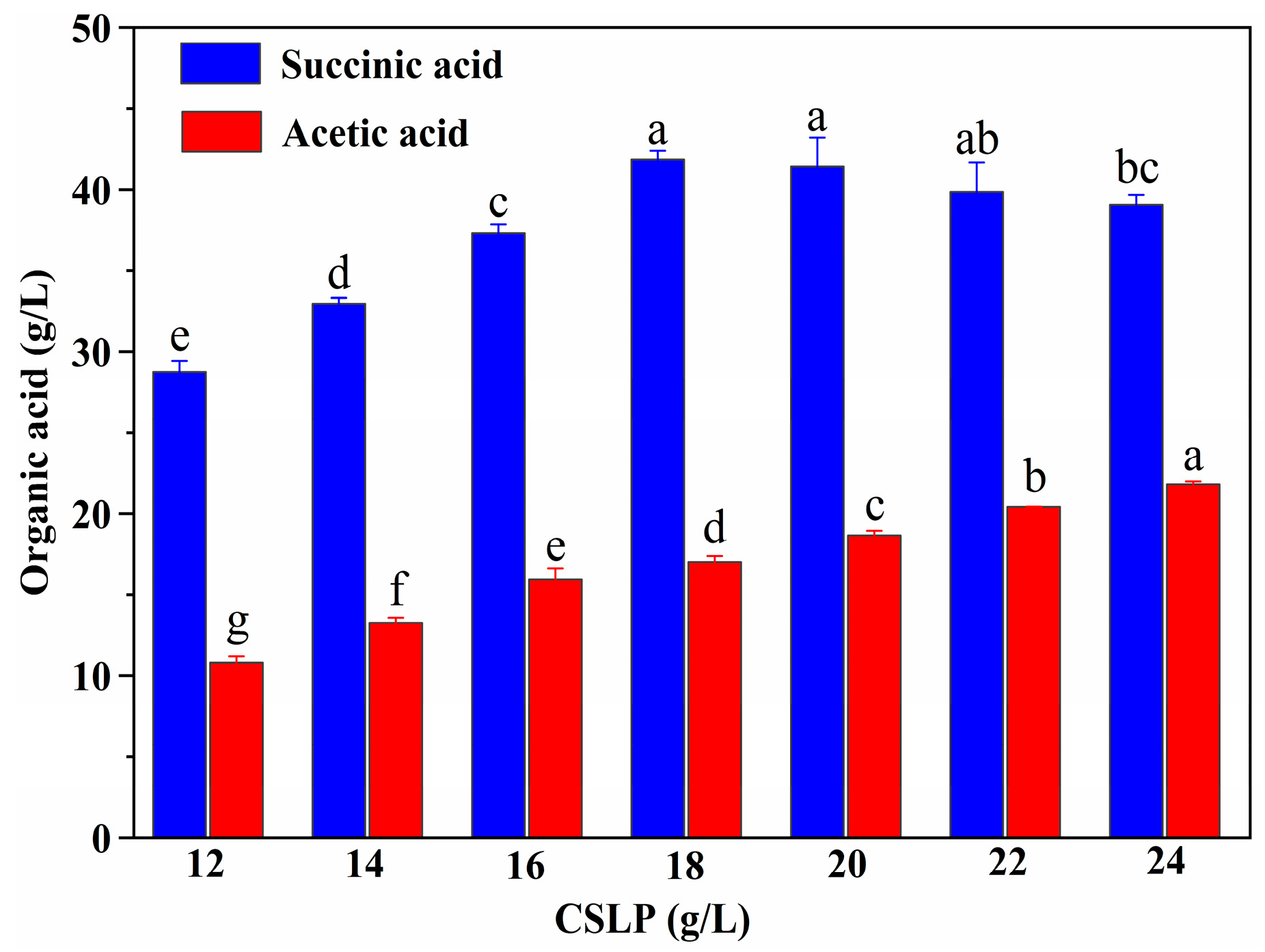
3.5. Effect of Nitrogen Source on SA Production from SCB Hydrolysate
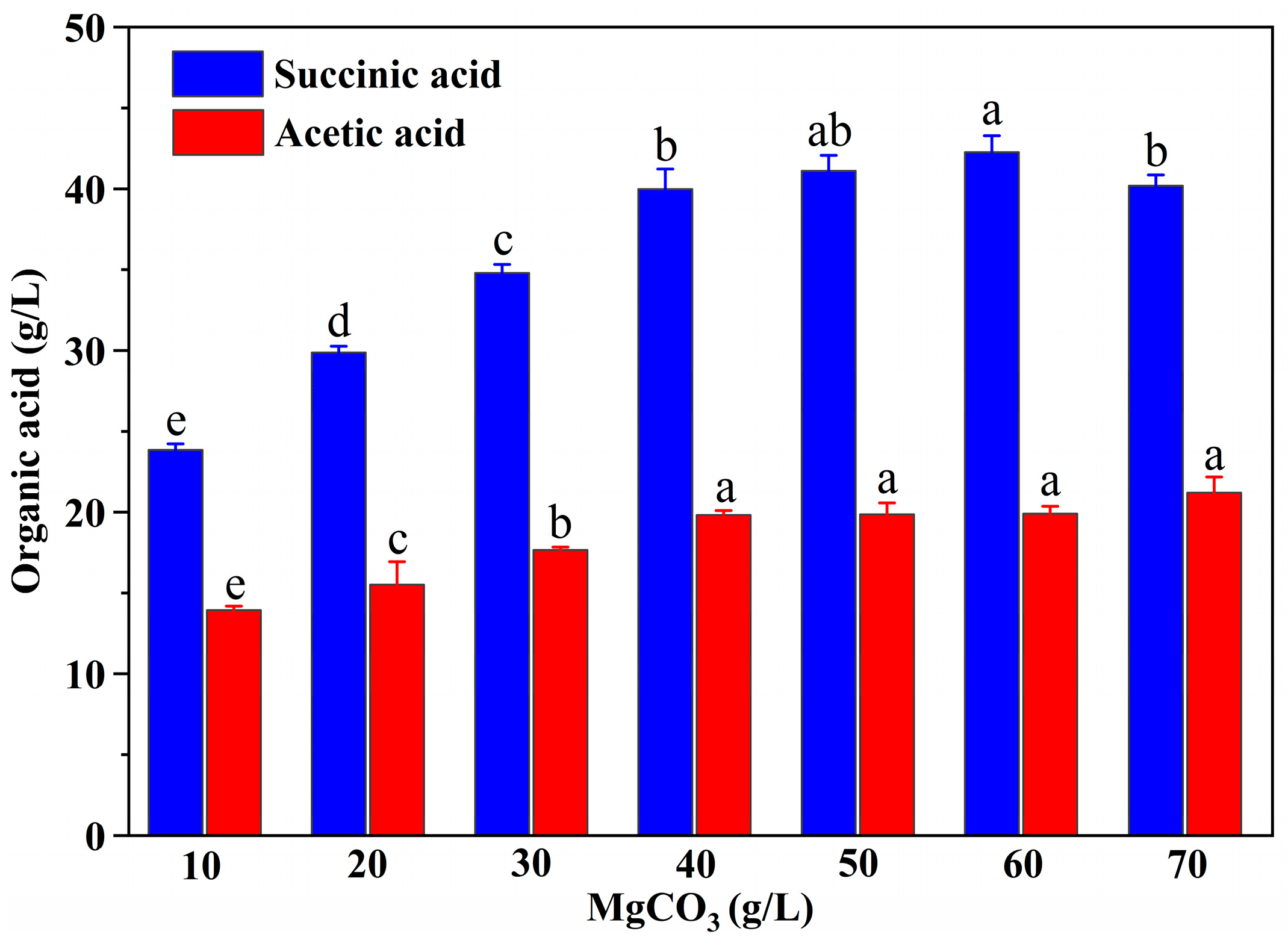
3.6. Effect of MgCO3 Supplementation on SA Production
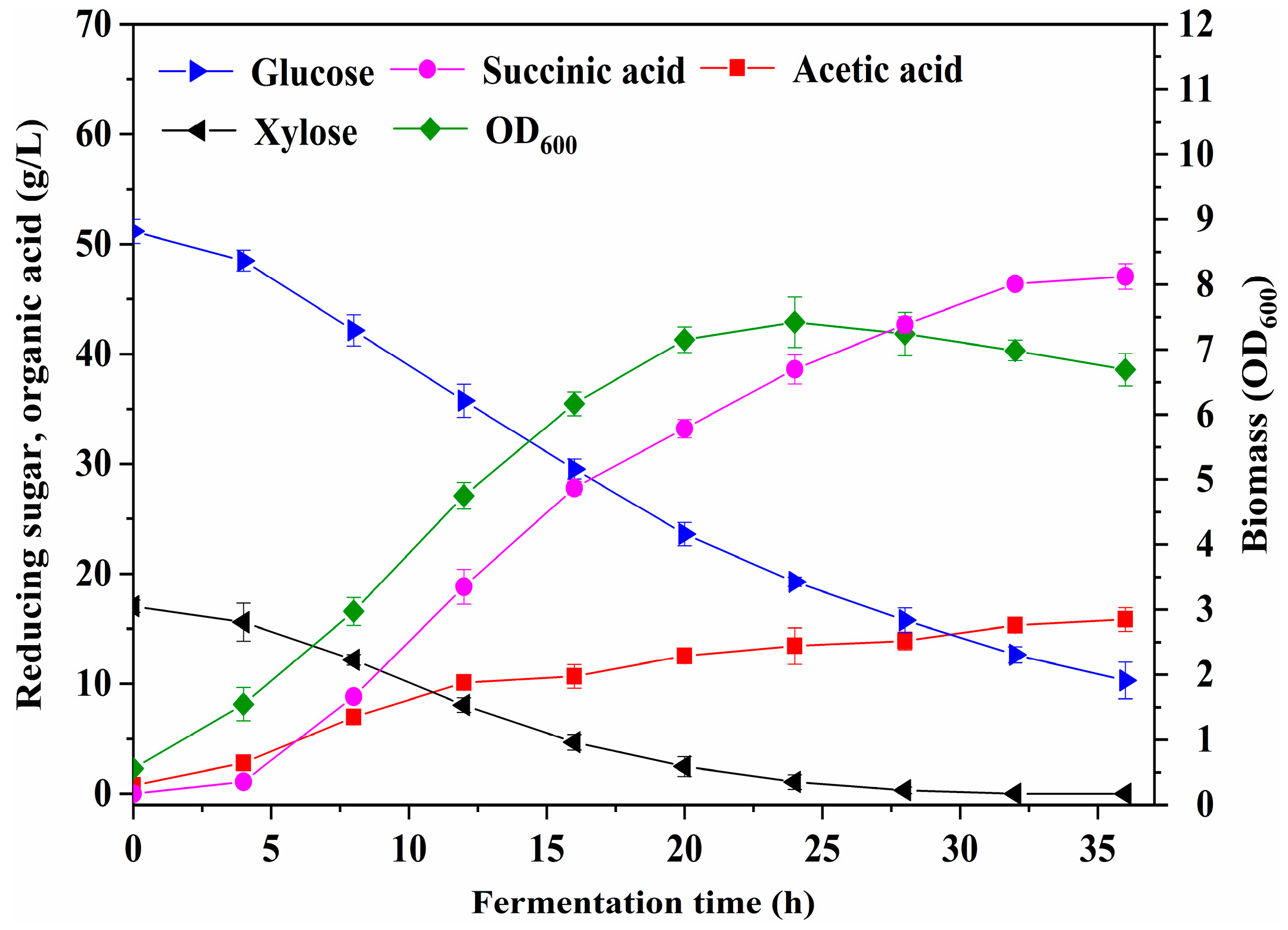
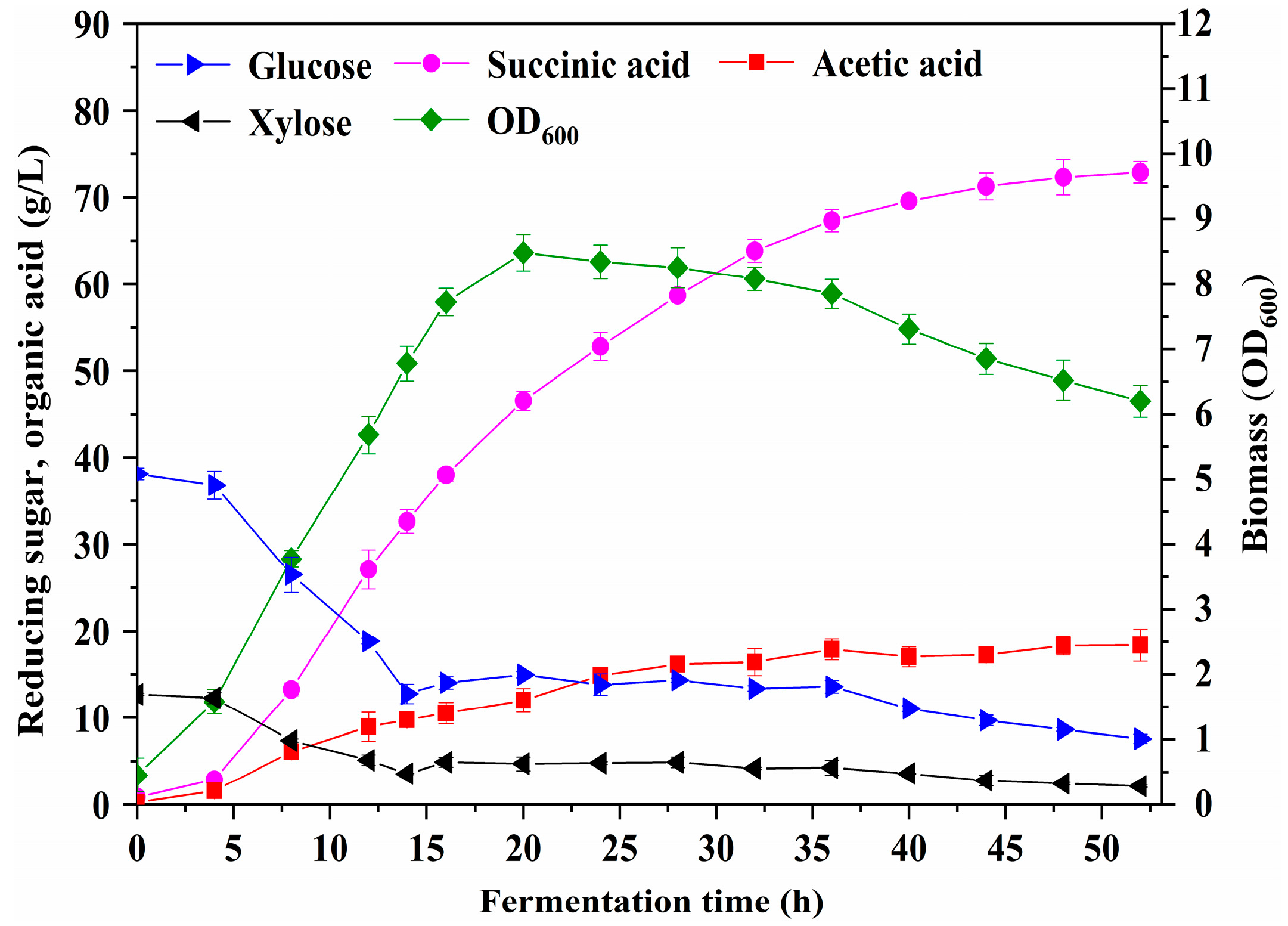
3.7. Fermentation in a 2 L Stirred Bioreactor
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Escanciano, I.A.; Ripoll, V.; Ladero, M.; Santos, V.E. Study on the Operational Modes Using Both Growing and Resting Cells for Succinic Acid Production from Xylose Kinetic Modelling. Fermentation 2023, 9, 663. [Google Scholar] [CrossRef]
- Ahn, J.H.; Jang, Y.-S.; Lee, S.Y. Production of succinic acid by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2016, 42, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.L.; Li, C.; Li, X.; Zhang, Y.; Xu, J.; Lin, C.S.K. Co-fermentation of glucose and xylose from sugarcane bagasse into succinic acid by Yarrowia lipolytica. Biochem. Eng. J. 2019, 148, 108–115. [Google Scholar] [CrossRef]
- Xu, C.; Alam, M.A.; Wang, Z.; Peng, Y.; Xie, C.; Gong, W.; Yang, Q.; Huang, S.; Zhuang, W.; Xu, J. Co-fermentation of succinic acid and ethanol from sugarcane bagasse based on full hexose and pentose utilization and carbon dioxide reduction. Bioresour. Technol. 2021, 339, 125578. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Guo, F.; Zhang, S.; Zhang, W.; Yang, Q.; Dong, W.; Jiang, M.; Ma, J.; Xin, F. Bio-based succinic acid: An overview of strain development, substrate utilization, and downstream purification. Biofuels Bioprod. Biorefining 2020, 14, 965–985. [Google Scholar] [CrossRef]
- Mancini, E.; Mansouri, S.S.; Gernaey, K.V.; Luo, J.; Pinelo, M. From second generation feed-stocks to innovative fermentation and downstream techniques for succinic acid production. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1829–1873. [Google Scholar] [CrossRef]
- Shen, N.; Li, S.; Qin, Y.; Jiang, M.; Zhang, H. Optimization of succinic acid production from xylose mother liquor (XML) by Actinobacillus succinogenes using response surface methodology. Biotechnol. Biotechnol. Equip. 2022, 36, 442–450. [Google Scholar] [CrossRef]
- Chen, J.; Yang, S.; Alam, M.A.; Wang, Z.; Zhang, J.; Huang, S.; Zhuang, W.; Xu, C.; Xu, J. Novel biorefining method for succinic acid processed from sugarcane bagasse. Bioresour. Technol. 2021, 324, 124615. [Google Scholar] [CrossRef]
- Efe, Ç.; van der Wielen, L.A.M.; Straathof, A.J.J. Techno-economic analysis of succinic acid production using adsorption from fermentation medium. Biomass Bioenergy 2013, 56, 479–492. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, V.; Singh, B. Cellulosic and hemicellulosic fractions of sugarcane bagasse: Potential, challenges and future perspective. Int. J. Biol. Macromol. 2021, 169, 564–582. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, B.; Chen, H.; Wu, W.; Wu, S.; Jin, Y.; Xiao, H. Recent advances in understanding the effects of lignin structural characteristics on enzymatic hydrolysis. Biotechnol. Biofuels 2021, 14, 205. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Tian, D.; Shen, F.; Hu, J.; Zhang, Y.; Yang, G.; Zeng, Y.; Deng, S.; Hu, Y. A comparative investigation of H2O2-involved pretreatments on lignocellulosic biomass for enzymatic hydrolysis. Biomass Convers. Biorefining 2019, 9, 321–331. [Google Scholar] [CrossRef]
- September, L.A.; Kheswa, N.; Seroka, N.S.; Khotseng, L. Green synthesis of silica and silicon from agricultural residue sugarcane bagasse ash-a mini review. RSC Adv. 2023, 13, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wen, Y.; Kapu, N.S. Ethanol production from bamboo using mild alkaline pre-extraction followed by alkaline hydrogen peroxide pretreatment. Bioresour. Technol. 2018, 247, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.M.D.; Queiroz, J.H.D.; Colodette, J.L. Assessment of alkaline pretreatment for the production of bioethanol from eucalyptus, sugarcane bagasse and sugarcane straw. Ind. Crops Prod. 2016, 94, 932–941. [Google Scholar] [CrossRef]
- Liu, T.; Williams, D.L.; Pattathil, S.; Li, M.; Hahn, M.G.; Hodge, D.B. Coupling alkaline pre-extraction with alkaline-oxidative post-treatment of corn stover to enhance enzymatic hydrolysis and fermentability. Biotechnol. Biofuels 2014, 7, 48. [Google Scholar] [CrossRef]
- Bolado-Rodríguez, S.; Toquero, C.; Martín-Juárez, J.; Travaini, R.; García-Encina, P.A. Effect of thermal, acid, alkaline and alkaline-peroxide pretreatments on the biochemical methane potential and kinetics of the anaerobic digestion of wheat straw and sugarcane bagasse. Bioresour. Technol. 2016, 201, 182–190. [Google Scholar] [CrossRef]
- Cardona, C.A.; Quintero, J.A.; Paz, I.C. Production of bioethanol from sugarcane bagasse: Status and perspectives. Bioresour. Technol. 2010, 101, 4754–4766. [Google Scholar] [CrossRef]
- Talebnia, F.; Karakashev, D.; Angelidaki, I. Production of bioethanol from wheat straw: An overview on pretreatment, hydrolysis and fermentation. Bioresour. Technol. 2010, 101, 4744–4753. [Google Scholar] [CrossRef]
- Tabka, M.G.; Herpoël-Gimbert, I.; Monod, F.; Asther, M.; Sigoillot, J.C. Enzymatic saccharification of wheat straw for bioethanol production by a combined cellulase xylanase and feruloyl esterase treatment. Enzyme Microb. Technol. 2006, 39, 897–902. [Google Scholar] [CrossRef]
- Chen, P.; Tao, S.; Zheng, P. Efficient and repeated production of succinic acid by turning sugarcane bagasse into sugar and support. Bioresour. Technol. 2016, 211, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xiang, B.; Liu, Y.; Huang, Q.; Yang, Y.; Chen, S.; Bai, G.; An, Q.; Cao, J.; Zheng, S.; et al. Synthesis of aqueous and hydroxy-terminated polyurethanes: Impacts of formulation parameters by orthogonal matrix design. Prog. Org. Coat. 2016, 90, 1–9. [Google Scholar] [CrossRef]
- Guettler, M.V.; Rumler, D.; Jain, M.K. Actinobacillus succinogenes sp. Nov., a novel succinic-acid-producing strain from the bovine rumen. Int. J. Syst. Evol. Microbiol. 1999, 49, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, Q.; Zhu, J.; Qin, Y.; Liao, S.; Li, Y.; Zhu, Q.; Jin, Y.; Du, L.; Huang, R. Succinic acid production from duckweed (Landoltia punctata) hydrolysate by batch fermentation of Actinobacillus succinogenes GXAS137. Bioresour. Technol. 2016, 211, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass: Laboratory Analytical Procedure (LAP); National Renewable Energy Laboratory: Golden, CO, USA, 2012. [Google Scholar]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.; Negro, M.J. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour. Technol. 2010, 101, 4851–4861. [Google Scholar]
- Xu, C.; Zhang, J.; Zhang, Y.; Guo, Y.; Xu, H.; Liang, C.; Wang, Z.; Xu, J. Lignin prepared from different alkaline pretreated sugarcane bagasse and its effect on enzymatic hydrolysis. Int. J. Biol. Macromol. 2019, 141, 484–492. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, J.; Zhang, Y.; Yu, Q.; Yuan, Z.; Liu, Y. Effects of different pretreatment methods on chemical composition of sugarcane bagasse and enzymatic hydrolysis. Bioresour. Technol. 2013, 144, 396–400. [Google Scholar]
- Hernández-Salas, J.M.; Villa-Ramírez, M.S.; Veloz-Rendón, J.S.; Rivera-Hernández, K.N.; González-César, R.A.; Plascencia-Espinosa, M.A.; Trejo-Estrada, S.R. Comparative hydrolysis and fermentation of sugarcane and agave bagasse. Bioresour. Technol. 2009, 100, 1238–1245. [Google Scholar] [CrossRef]
- Zheng, P.; Dong, J.-J.; Sun, Z.-H.; Ni, Y.; Fang, L. Fermentative production of succinic acid from straw hydrolysate by Actinobacillus succinogenes. Bioresour. Technol. 2009, 100, 2425–2429. [Google Scholar] [CrossRef]
- Romero-Rodríguez, A.; Maldonado-Carmona, N.; Ruiz-Villafán, B.; Koirala, N.; Rocha, D.; Sánchez, S. Interplay between carbon, nitrogen and phosphate utilization in the control of secondary metabolite production in streptomyces. Antonie Leeuwenhoek 2018, 111, 761–781. [Google Scholar] [CrossRef]
- Xi, Y.-L.; Chen, K.-Q.; Xu, R.; Zhang, J.-H.; Bai, X.-F.; Jiang, M.; Wei, P.; Chen, J.-Y. Effect of biotin and a similar compound on succinic acid fermentation by Actinobacillus succinogenes in a chemically defined medium. Biochem. Eng. J. 2012, 69, 87–92. [Google Scholar] [CrossRef]
- Xi, Y.-L.; Chen, K.-Q.; Dai, W.-Y.; Ma, J.-F.; Zhang, M.; Jiang, M.; Wei, P.; Ouyang, P.-K. Succinic acid production by Actinobacillus succinogenes NJ113 using corn steep liquor powder as nitrogen source. Bioresour. Technol. 2013, 136, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, M.; Dai, Z.; Xin, F.; Zhou, J.; Dong, W.; Ma, J.; Jiang, M.; Zhang, W. Comprehensive investigation of succinic acid production by Actinobacillus succinogenes: A promising native succinic acid producer. Biofuels, Bioprod. Biorefin. 2020, 14, 950–964. [Google Scholar] [CrossRef]
- Sheng, L.; Zhu, G.; Tong, Q. Comparative proteomic analysis of aureobasidium pullulans in the presence of high and low levels of nitrogen source. J. Agric. Food Chem. 2014, 62, 10529–10534. [Google Scholar] [CrossRef]
- Jokodola, E.O.; Narisetty, V.; Castro, E.; Durgapal, S.; Coulon, F.; Sindhu, R.; Binod, P.; Banu, J.R.; Kumar, G.; Kumar, V. Process optimisation for production and recovery of succinic acid using xylose-rich hydrolysates by Actinobacillus succinogenes. Bioresour. Technol. 2022, 344, 126224. [Google Scholar]
- Banger, G.; Kaya, K.; Omwene, P.; Shakoory, S.; Karagündüz, A.; Keskinler, B.; Nikerel, E. Delactosed whey permeate as substrate for succinic acid fermentation by Actinobacillus succinogenes. Waste Biomass Valorization 2021, 12, 5481–5489. [Google Scholar] [CrossRef]
- Saxena, R.K.; Saran, S.; Isar, J.; Kaushik, R. 27-Production and Applications of Succinic Acid. In Current Developments in Biotechnology and Bioengineering, Production, Isolation and Purification of Industrial Products; Pandey, A., Negi, S., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 601–630. [Google Scholar]
- Dorado, M.P.; Lin, S.K.C.; Koutinas, A.; Du, C.; Wang, R.; Webb, C. Cereal-based biorefinery development: Utilisation of wheat milling by-products for the production of succinic acid. J. Biotechnol. 2009, 143, 51–59. [Google Scholar] [CrossRef]
- Dessie, W.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Opportunities, challenges, and future perspectives of succinic acid production by Actinobacillus succinogenes. Appl. Microbiol. Biotechnol. 2018, 102, 9893–9910. [Google Scholar] [CrossRef]
- Shen, N.; Zhang, H.; Qin, Y.; Wang, Q.; Zhu, J.; Li, Y.; Jiang, M.-G.; Huang, R. Efficient production of succinic acid from duckweed (Landoltia punctata) hydrolysate by Actinobacillus succinogenes GXAS137. Bioresour. Technol. 2018, 250, 35–42. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, H.; Miao, Y.; Jiang, M.; Chen, J. Succinic acid production from enzymatic hydrolysate of sake lees using Actinobacillus succinogenes 130z. Enzyme Microb. Technol. 2010, 47, 236–240. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, Z.; Zheng, Y.; Zhou, J.; Xiu, Z. Efficient production of lactic acid from sugarcane molasses by a newly microbial consortium CEE-DL15. Process Biochem. 2019, 81, 132–138. [Google Scholar] [CrossRef]
- Li, Q.; Lei, J.; Zhang, R.; Li, J.; Xing, J.; Gao, F.; Gong, F.; Yan, X.; Wang, D.; Su, Z.; et al. Efficient decolorization and deproteinization using uniform polymer microspheres in the succinic acid biorefinery from bio-waste cotton (Gossypium hirsutum L.) stalks. Bioresour. Technol. 2013, 135, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, J.; Hassan, N.; Idris, A.; Ngadiman, N.H.A. Optimization of simultaneous saccharification and fermentation process conditions for the production of succinic acid from oil palm empty fruit bunches. J. Wood Chem. Technol. 2020, 40, 136–145. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, H.; Miao, Y.; Wei, P.; Chen, J. Simultaneous saccharification and fermentation of acid-pretreated rapeseed meal for succinic acid production using Actinobacillus succinogenes. Enzyme Microb. Technol. 2011, 48, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.; Roca, C.; Reis, M.A.M. Improving succinic acid production by Actinobacillus succinogenes from raw industrial carob pods. Bioresour. Technol. 2016, 218, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Bradfield, M.F.A.; Mohagheghi, A.; Salvachúa, D.; Smith, H.; Black, B.A.; Dowe, N.; Beckham, G.T.; Nicol, W. Continuous succinic acid production by Actinobacillus succinogenes on xylose-enriched hydrolysate. Biotechnol. Biofuels 2015, 8, 181. [Google Scholar] [CrossRef]
- Filippi, K.; Georgaka, N.; Alexandri, M.; Papapostolou, H.; Koutinas, A. Valorisation of grape stalks and pomace for the production of bio-based succinic acid by Actinobacillus succinogenes. Ind. Crops Prod. 2021, 168, 113578. [Google Scholar] [CrossRef]
- Dessie, W.; Zhu, J.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Bio-succinic acid production from coffee husk treated with thermochemical and fungal hydrolysis. Bioprocess Biosyst. Eng. 2018, 41, 1461–1470. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pretreatment | |||||
|---|---|---|---|---|---|
| Raw | NaOH | AHP | NaOH-AHP | ||
| Solid remain (%) | — | 56.03 ± 0.21 b | 63.67 ± 0.33 a | 46.97 ± 0.18 c | |
| Cellulose | Content (%) | 41.32 ± 0.15 d | 57.44 ± 0.63 b | 54.23 ± 0.09 c | 64.45 ± 0.22 a |
| Recovery (%) | — | 77.89 ± 0.41 b | 83.56 ± 0.23 a | 73.26 ± 0.26 c | |
| Hemicellulose | Content (%) | 24.47 ± 0.11 b | 26.06 ± 0.03 a | 23.53 ± 0.43 b | 22.53 ± 0.22 c |
| Recovery (%) | — | 59.68 ± 0.53 b | 61.22 ± 0.62 a | 43.24 ± 0.18 c | |
| Lignin | Content (%) | 26.42 ± 0.08 a | 11.76 ± 0.12 c | 15.36 ± 0.46 b | 9.89 ± 0.37 d |
| Removal (%) | — | 75.08 ± 0.76 b | 62.99 ± 0.59 c | 82.42 ± 0.24 a | |
| FU and HMF (mg/L) | None | None | None | None | |
| Glucose (g/L) | 0.51 ± 0.08 d | 41.65 ± 0.32 b | 37.63 ± 0.78 c | 51.31 ± 0.32 a | |
| Cellulose hydrolysis rate (%) | 1.16 ± 0.02 d | 67.87 ± 0.48 b | 64.95 ± 0.61 c | 74.52 ± 0.38 a | |
| Xylose (g/L) | 0.13 ± 0.01 d | 20.14 ± 0.06 a | 13.66 ± 0.62 c | 17.12 ± 0.15 b | |
| Hemicellulose hydrolysis rate (%) | 0.49 ± 0.08 c | 70.73 ± 0.27 a | 53.13 ± 0.75 b | 69.54 ± 0.26 a | |
| Total reducing sugar (g/L) | 0.64 ± 0.06 d | 61.79 ± 0.35 b | 51.29 ± 0.82 c | 68.43 ± 1.02 a | |
| Total reducing sugar yield (%) | 0.9 ± 0.00 d | 68.75 ± 0.28 b | 61.29 ± 0.75 c | 73.20 ± 0.22 a | |
| Test No. | Factors (Levels) | Reducing Sugar Concentration (g/L) | |||
|---|---|---|---|---|---|
| (A) Cellulase Concentration (% v/w-Biomass) | (B) Xylanase Concentration (% v/w-biomass) | (C) Pectinase Concentration (% v/w-biomass) | Null Column | ||
| 1 | 10.00 (1) | 2.00 (1) | 10.00 (1) | (1) | 60.96 ± 0.15 |
| 2 | 10.00 (1) | 3.00 (2) | 15.00 (2) | (2) | 59.75 ± 0.06 |
| 3 | 10.00 (1) | 4.00 (3) | 20.00 (3) | (3) | 58.05 ± 0.77 |
| 4 | 15.00 (2) | 2.00 (1) | 15.00 (2) | (3) | 61.77 ± 0.93 |
| 5 | 15.00 (2) | 3.00 (2) | 20.00 (3) | (1) | 67.87 ± 0.77 |
| 6 | 15.00 (2) | 4.00 (3) | 10.00 (1) | (2) | 76.90 ± 0.75 |
| 7 | 20.00 (3) | 2.00 (1) | 20.00 (3) | (2) | 70.47 ± 0.52 |
| 8 | 20.00 (3) | 3.00 (2) | 10.00 (1) | (3) | 77.13 ± 0.31 |
| 9 | 20.00 (3) | 4.00 (3) | 15.00 (2) | (1) | 75.10 ± 0.86 |
| k1 | 59.59 | 64.40 | 71.66 | 67.98 | |
| k2 | 68.85 | 68.25 | 65.54 | 69.04 | |
| k3 | 74.23 | 70.02 | 65.46 | 65.65 | |
| R | 14.65 | 5.62 | 6.20 | 3.39 | |
| Best level | A3 | B3 | C1 | 78.34 ± 0.87 | |
| Initial Reducing Sugar (g/L) | SA (g/L) | AA (g/L) | Residual Reducing Sugar (g/L) | SA Yield (%) |
|---|---|---|---|---|
| 40 | 25.28 ± 1.93 d | 13.78 ± 1.16 c | 6.17 ± 0.12 f | 74.72 ± 0.32 a |
| 50 | 30.90 ± 2.74 c | 16.85 ± 1.68 b | 8.96 ± 0.11 e | 75.29 ± 0.55 a |
| 60 | 35.58 ± 1.91 b | 18.41 ± 0.33 ab | 11.94 ± 0.74 d | 74.03 ± 0.46 a |
| 70 | 40.56 ± 0.36 a | 19.09 ± 0.57 ab | 17.11 ± 0.54 c | 76.70 ± 0.27 a |
| 80 | 38.70 ± 0.95 ab | 19.54 ± 0.31 a | 28.37 ± 0.42 b | 74.95 ± 0.13 a |
| 90 | 38.39 ± 0.61 ab | 18.98 ± 0.19 ab | 36.15 ± 1.31 a | 71.29 ± 0.26 a |
| Substrate | Nitrogen Resource (g/L) | Fermentation Type | SA Titer (g/L) | SA Yield (%) | SA Productivity (g/L/h) | Reference |
|---|---|---|---|---|---|---|
| Cotton stalk | YE (30) | Batch | 63.0 | 64.0 | 1.17 | 2013 Li [44] |
| Oil palm empty fruit bunches | YE (20) and CSL (20) | Batch | 42.9 | 61.3 | 0.89 | 2019 Akhtar [45] |
| Rapeseed meal | YE (15) | Fed-batch | 23.4 | 11.5 | 0.33 | 2011 Chen [46] |
| Carob Pods | YE (10) | Batch | 9.4 | 54.0 | 1.32 | 2016 Carvalho [47] |
| Fed-batch | 19.0 | 94.0 | 1.43 | |||
| Corn stover | YE (6) and CSL (10) | Continuous | 39.6 | 78.0 | 1.77 | 2015 Bradfield [48] |
| Grape pomace and stalks | YE (5) | Batch | 24.6 | 47.0 | 0.75 | 2021 Filippi [49] |
| Fed-batch | 40.2 | 67.0 | 0.79 | |||
| Sake lees | YE (2.5) | Batch | 48.0 | 75.5 | 0.94 | 2010 Chen [42] |
| Coffee husk | CHH | Batch | 19.3 | 95.0 | 0.54 | 2018 Dessie [50] |
| Sugarcane bagasse | CSLP (20) | Batch | 39.9 | 82.0 | 1.37 | 2016 Chen [21] |
| Fed-batch | 70.8 | 81.5 | 1.42 | |||
| Sugarcane bagasse | SBH | Fed-batch | 41.0 | 32.0 | 0.30 | 2021 Chen [8] |
| Sugarcane bagasse | CSLP (18) | Batch | 47.1 | 81.3 | 1.31 | This study |
| Fed-batch | 72.9 | 83.2 | 1.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Li, Y.; Liang, G.; Shen, N.; Xian, L.; Wang, Q. Efficient Production of Succinic Acid from Sugarcane Bagasse Hydrolysate by Actinobacillus succinogenes GXAS137. Fermentation 2024, 10, 22. https://doi.org/10.3390/fermentation10010022
Qin Y, Li Y, Liang G, Shen N, Xian L, Wang Q. Efficient Production of Succinic Acid from Sugarcane Bagasse Hydrolysate by Actinobacillus succinogenes GXAS137. Fermentation. 2024; 10(1):22. https://doi.org/10.3390/fermentation10010022
Chicago/Turabian StyleQin, Yan, Yi Li, Ge Liang, Naikun Shen, Liang Xian, and Qingyan Wang. 2024. "Efficient Production of Succinic Acid from Sugarcane Bagasse Hydrolysate by Actinobacillus succinogenes GXAS137" Fermentation 10, no. 1: 22. https://doi.org/10.3390/fermentation10010022
APA StyleQin, Y., Li, Y., Liang, G., Shen, N., Xian, L., & Wang, Q. (2024). Efficient Production of Succinic Acid from Sugarcane Bagasse Hydrolysate by Actinobacillus succinogenes GXAS137. Fermentation, 10(1), 22. https://doi.org/10.3390/fermentation10010022