

Dynamic Optimization of Lactic Acid Production from Grape Stalk Solid-State Fermentation with Rhizopus oryzae Applying a Variable Temperature Profile
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solid-State Fermentation
- (a)
- When using a small-scale experimental growth chamber (membrane filter culture system), heat transfer phenomena are not limiting. Therefore, it can be assumed that the solid substrate is at the temperature and ambient relative humidity of the incubator chamber air.
- (b)
- Petri dishes were incubated at different constant temperatures (maintaining constant relative humidity). The ranges of temperature were chosen to encompass optimal parameters with the minimum number of trials. These conditions remained constant in each culture throughout the growth cycle.
- (c)
- All Petri dishes were inoculated with the same spore suspension at the same concentration.
- (d)
- In SSF, pH is a very difficult parameter to measure and control. Additionally, R. oryzae is a strain capable of growing in a range of pH from 4 to 9 [21]. So, the pH was neither measured nor controlled in the experiments.
2.2. Mathematical Bioprocess Model
2.2.1. Parametric Identification
2.2.2. Effect of Temperature Polynomial Relationship on the Kinetic Parameters of R. oryzae Growth and Lactic Acid Production
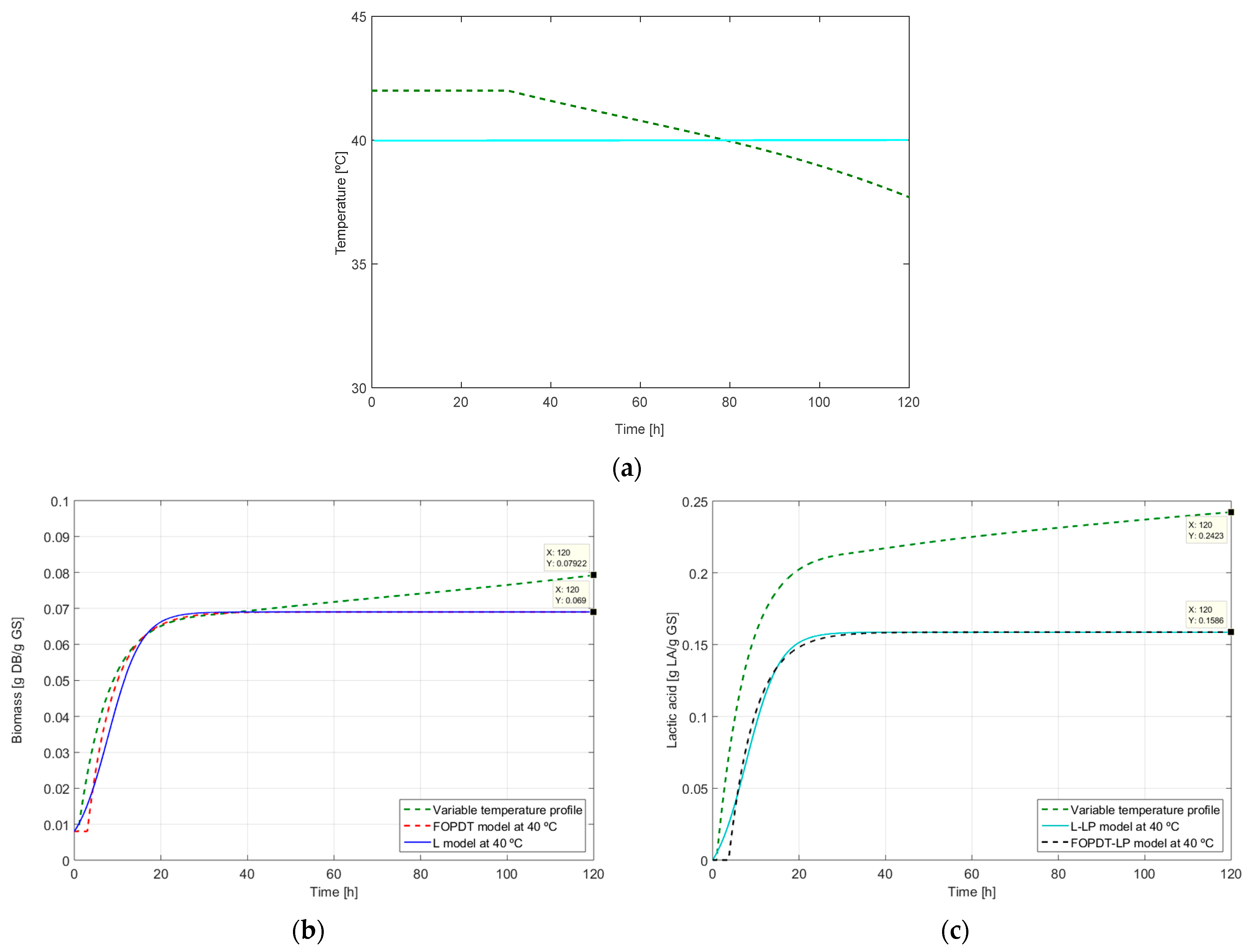
2.3. Dynamic Optimization of LA Production
3. Results and Discussion
3.1. SSF and Mathematical Bioprocess Model
3.2. Effect of Temperature Polynomial Relationship on the Kinetic Parameters of R. oryzae Growth and Lactic Acid Production
3.3. Effect of Temperature on the Growth Morphology of R. oryzae NCIM 1299
3.4. Optimal Control Problem Statement
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawoof, S.A.A.; Kumar, P.S.; Vo, D.V.N.; Devaraj, K.; Mani, Y.; Devaraj, T.; Subramanian, S. Production of Optically Pure Lactic Acid by Microbial Fermentation: A Review. Environ. Chem. Lett. 2021, 19, 539–556. [Google Scholar] [CrossRef]
- Krishna, B.S.; Sai Nikhilesh, G.S.; Tarun, B.; Narayana Saibaba, K.V.; Gopinadh, R. Industrial Production of Lactic Acid and Its Applications. Int. J. Biotech Res. 2018, 1, 42–54. [Google Scholar] [CrossRef]
- Ameen, S.M.; Caruso, G. Lactic Acid in the Food Industry; Springer: Cham, Switzerland, 2017; ISBN 9783319581460. [Google Scholar]
- Pohanka, M. D-Lactic Acid as a Metabolite: Toxicology, Diagnosis, and Detection. Biomed Res. Int. 2020, 2020, 3419034. [Google Scholar] [CrossRef] [PubMed]
- Alves De Oliveira, R.; Komesu, A.; Vaz Rossell, C.E.; Filho, R.M. Challenges and Opportunities in Lactic Acid Bioprocess Design—From Economic to Production Aspects. Biochem. Eng. J. 2018, 133, 219–239. [Google Scholar] [CrossRef]
- L(+)-Lactic Acid. Available online: https://www.jungbunzlauer.com/en/products/lactics/l-lactic-acid#:~:text=The%20US%20Food%20and%20Drug,flavor%20enhancer%2C%20fla-voring%20agent%20or (accessed on 16 November 2023).
- Thongchul, N. Production of Lactic Acid and Polylactic Acid for Industrial Applications. In Bioprocessing Technologies in Biorefinery for Sustainable Production of Fuels, Chemicals, and Polymers; Yang, S.-T., El-Enshasy, H.A., Thongchul, N., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 293–316. ISBN 9780470541951. [Google Scholar]
- Chung, M.R.W.Y.; Tan, I.S.; Foo, H.C.Y.; Lam, M.K.; Lim, S. Potential of Macroalgae-Based Biorefinery for Lactic Acid Production from Exergy Aspect. Biomass Convers. Biorefinery 2023, 13, 2623–2653. [Google Scholar] [CrossRef]
- LinkedIn. ClimateTech Food Waste Upcycling Process Goes Commercial. Available online: https://www.linkedin.com/pulse/climatetech-food-waste-upcycling-process-goes-commercial-%3FtrackingId=AmaRJwjwR0q2e1xeCkITYA%253D%253D/?trackingId=AmaRJwjwR0q2e1xeCkITYA%3D%3D (accessed on 30 January 2024).
- Groff, M.C.; Scaglia, G.; Gaido, M.; Kassuha, D.; Ortiz, O.A.; Noriega, S.E. Kinetic Modeling of Fungal Biomass Growth and Lactic Acid Production in Rhizopus oryzae Fermentation by Using Grape Stalk as a Solid Substrate. Biocatal. Agric. Biotechnol. 2022, 39, 102255. [Google Scholar] [CrossRef]
- Möller, J.; Hernández Rodríguez, T.; Müller, J.; Arndt, L.; Kuchemüller, K.B.; Frahm, B.; Eibl, R.; Eibl, D.; Pörtner, R. Model Uncertainty-Based Evaluation of Process Strategies during Scale-up of Biopharmaceutical Processes. Comput. Chem. Eng. 2020, 134, 106693. [Google Scholar] [CrossRef]
- Mazaheri, D.; Shojaosadati, S.A. Mathematical Models for Microbial Kinetics in Solid-State Fermentation: A Review. Iran. J. Biotechnol. 2013, 11, 156–167. [Google Scholar] [CrossRef]
- Groff, M.C.; Scaglia, G.; Ortiz, O.A.; Noriega, S.E. Modification of the Luedeking and Piret Model with a Delay Time Parameter for Biotechnological Lactic Acid Production. Biotechnol. Lett. 2022, 44, 415–427. [Google Scholar] [CrossRef]
- Groff, C.; Kuchen, B.; Gil, R.; Fernández, C. Scaglia Application of the Luedeking and Piret with Delay Time Model in Bioproductions with Non-Zero Kinetic Parameters. IEEE Lat. Am. Trans. 2023, 21, 882–888. [Google Scholar] [CrossRef]
- Leon, G.; Cujilema, M.; Barloyo, L.; Rosero, E.; Córdova, J.; Beltrán, L. Determination of the Optimum Temperature for the Cellulase Production by Aspergillus niger in Solid Fermentation. Rev. Cent. Azúcar 2018, 45, 1–13. [Google Scholar]
- Narayanan, H.; Luna, M.F.; von Stosch, M.; Cruz Bournazou, M.N.; Polotti, G.; Morbidelli, M.; Butté, A.; Sokolov, M. Bioprocessing in the Digital Age: The Role of Process Models. Biotechnol. J. 2020, 15, e1900172. [Google Scholar] [CrossRef] [PubMed]
- Noll, P.; Lilge, L.; Hausmann, R.; Henkel, M. Modeling and Exploiting Microbial Temperature Response. Processes 2020, 8, 121. [Google Scholar] [CrossRef]
- Pantano, M.N.; Fernández, M.C.; Amicarelli, A.; Scaglia, G.J.E. Evolutionary Algorithms and Orthogonal Basis for Dynamic Optimization in L2 Space for Batch Biodiesel Production. Chem. Eng. Res. Des. 2022, 177, 354–364. [Google Scholar] [CrossRef]
- Pantano, M.N.; Fernández, M.C.; Ortiz, O.A.; Scaglia, G.J.E.; Vega, J.R. A Fourier-Based Control Vector Parameterization for the Optimization of Nonlinear Dynamic Processes with a Finite Terminal Time. Comput. Chem. Eng. 2020, 134, 106721. [Google Scholar] [CrossRef]
- Borshchevskaya, L.N.; Gordeeva, T.L.; Kalinina, A.N.; Sineokii, S.P. Spectrophotometric Determination of Lactic Acid. J. Anal. Chem. 2016, 71, 755–758. [Google Scholar] [CrossRef]
- Londoño-Hernández, L.; Ramírez-Toro, C.; Ruiz, H.A.; Ascacio-Valdés, J.A.; Aguilar-Gonzalez, M.A.; Rodríguez-Herrera, R.; Aguilar, C.N. Rhizopus oryzae—Ancient Microbial Resource with Importance in Modern Food Industry. Int. J. Food Microbiol. 2017, 257, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Fernández, C.; Pantano, N.; Godoy, S.; Serrano, E.; Scaglia, G. Optimización de Parámetros Utilizando Los Métodos de Monte Carlo y Algoritmos Evolutivos. Aplicación a Un Controlador de Seguimiento de Trayectoria En Sistemas No Lineales. Rev. Iberoam. Autom. Inform. Ind. 2018, 16, 89–99. [Google Scholar] [CrossRef]
- Feng, J.; Feng, N.; Zhang, J.S.; Yang, Y.; Jia, W.; Lin, C.C. A New Temperature Control Shifting Strategy for Enhanced Triterpene Production by Ganoderma Lucidum G0119 Based on Submerged Liquid Fermentation. Appl. Biochem. Biotechnol. 2016, 180, 740–752. [Google Scholar] [CrossRef]
- Cheng, K.K.; Zeng, J.; Jian, J.H.; Zhu, J.F.; Zhang, G.X.; Liu, D.H. Model-Based Temperature Control for Improving Lactic Acid Production from Glycerol. RSC Adv. 2019, 9, 11614–11620. [Google Scholar] [CrossRef]
- Saithi, S.; Borg, J.; Nopharatana, M.; Tongta, A. Mathematical Modeling of Biomass and Enzyme Production Kinetics by Aspergillus Niger in Solid-State Fermentation at Various Temperatures and Moisture Contents. J. Microb. Biochem. Technol. 2016, 8, 123–130. [Google Scholar] [CrossRef]
- Ramos-Sánchez, L.B.; Leon-Revelo, G.; Baryolo, L.; Cujilema-Quitio, M.C.; Rosero Delgado, E.; Córdova, J. Modelo Cinético Para La Producción de Celulasas Por Una Cepa de Aspergillus niger En Fermentación Sólida. Rev. Cent. Azúcar 2018, 45, 1–13. [Google Scholar]
- Hamidi-Esfahani, Z.; Shojaosadati, S.A.; Rinzema, A. Modelling of Simultaneous Effect of Moisture and Temperature on A. niger Growth in Solid-State Fermentation. Biochem. Eng. J. 2004, 21, 265–272. [Google Scholar] [CrossRef]
- Huang, L.P.; Jin, B.; Lant, P.; Zhou, J. Simultaneous Saccharification and Fermentation of Potato Starch Wastewater to Lactic Acid by Rhizopus oryzae and Rhizopus arrhizus. Biochem. Eng. J. 2005, 23, 265–276. [Google Scholar] [CrossRef]
- Takano, M.; Hoshino, K. Lactic Acid Production from Paper Sludge by SSF with Thermotolerant Rhizopus sp. Bioresurces Bioprocess. 2016, 3, 29. [Google Scholar] [CrossRef]
- Wang, X.; Sun, L.; Wei, D.; Wang, R. Reducing By-Product Formation in L-Lactic Acid Fermentation by Rhizopus oryzae. J. Ind. Microbiol. Biotechnol. 2005, 32, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Longacre, A.; Reimers, J.M.; Gannon, J.E.; Wright, B.E. Flux Analysis of Glucose Metabolism in Rhizopus oryzae for the Purpose of Increasing Lactate Yields. Fungal Genet. Biol. 1997, 21, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Das, R.K.; Brar, S.K. Enhanced Fumaric Acid Production from Brewery Wastewater and Insight into the Morphology of Rhizopus oryzae 1526. Appl. Biochem. Biotechnol. 2014, 172, 2974–2988. [Google Scholar] [CrossRef]
- Aghababaie, M.; Beheshti, M.; Khanahmadi, M. Effect of Temperature and PH on Formulating the Kinetic Growth Parameters and Lactic Acid Production of Lactobacillus bulgaricus. Nutr. Food Sci. Res. 2014, 1, 49–56. [Google Scholar]
- Brandam, C.; Castro-Martínez, C.; Délia, M.L.; Ramón-Portugal, F.; Strehaiano, P. Effect of Temperature on Brettanomyces bruxellensis: Metabolic and Kinetic Aspects. Can. J. Microbiol. 2008, 54, 11–18. [Google Scholar] [CrossRef]
- Krull, R.; Wucherpfennig, T.; Esfandabadi, M.E.; Walisko, R.; Melzer, G.; Hempel, D.C.; Kampen, I.; Kwade, A.; Wittmann, C. Characterization and Control of Fungal Morphology for Improved Production Performance in Biotechnology. J. Biotechnol. 2013, 163, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Du, J.; Tsao, G. Mycelial Pellet Formation by Rhizopus oryzae ATCC 20344. Appl. Biochem. Biotechnol. 2000, 84–86, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Veiter, L.; Rajamanickam, V.; Herwig, C. The Filamentous Fungal Pellet—Relationship between Morphology and Productivity. Appl. Microbiol. Biotechnol. 2018, 102, 2997–3006. [Google Scholar] [CrossRef]
- Miyazawa, K.; Yoshimi, A.; Yoshimi, A.; Abe, K.; Abe, K.; Abe, K. The Mechanisms of Hyphal Pellet Formation Mediated by Polysaccharides, α-1,3-Glucan and Galactosaminogalactan, in Aspergillus Species. Fungal Biol. Biotechnol. 2020, 7, 10. [Google Scholar] [CrossRef]
- Liao, W.; Liu, Y.; Chen, S.L. Studying Pellet Formation of a Filamentous Fungus Rhizopus oryzae to Enhance Organic Acid Production. Appl. Biochem. Biotechnol. 2007, 137, 689–701. [Google Scholar]
- Esaú López-Jácome, L.; Hernández-Durán, M.; Colín-Castro, C.A.; Ortega-Peña, S.; Cerón-González, G.; Franco-Cendejas, R. Las Tinciones Básicas En El Laboratorio de Microbiología. Investig. Discapac. 2014, 3, 10–18. [Google Scholar]
- Karmakar, M.; Ghosh, B.; Ray, R. Effect of Extracellular Factors on Growth and Dimorphism of Rhizopus Oryzae with Multiple Enzyme Synthesizing Ability. Indian J. Microbiol. 2012, 52, 215–221. [Google Scholar] [CrossRef]
- Böl, M.; Schrinner, K.; Tesche, S.; Krull, R. Challenges of Influencing Cellular Morphology by Morphology Engineering Techniques and Mechanical Induced Stress on Filamentous Pellet Systems—A Critical Review. Eng. Life Sci. 2020, 21, 51–67. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, J.; Ou, W.; Chen, Y.; Wang, R.; Li, K.; Sun, X.; Li, Y.; Xu, Q.; Huang, H. Biotechnology for Biofuels Transcriptome Analysis of Rhizopus oryzae Seed Pellet Formation Using Triethanolamine. Biotechnol. Biofuels 2021, 14, 230. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.; Manan, M.A. Design Aspects of Solid State Fermentation as Applied to Microbial Bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 511–532. [Google Scholar] [CrossRef]
- Pandey, A. Solid-State Fermentation. Biochem. Eng. J. 2003, 13, 81–84. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
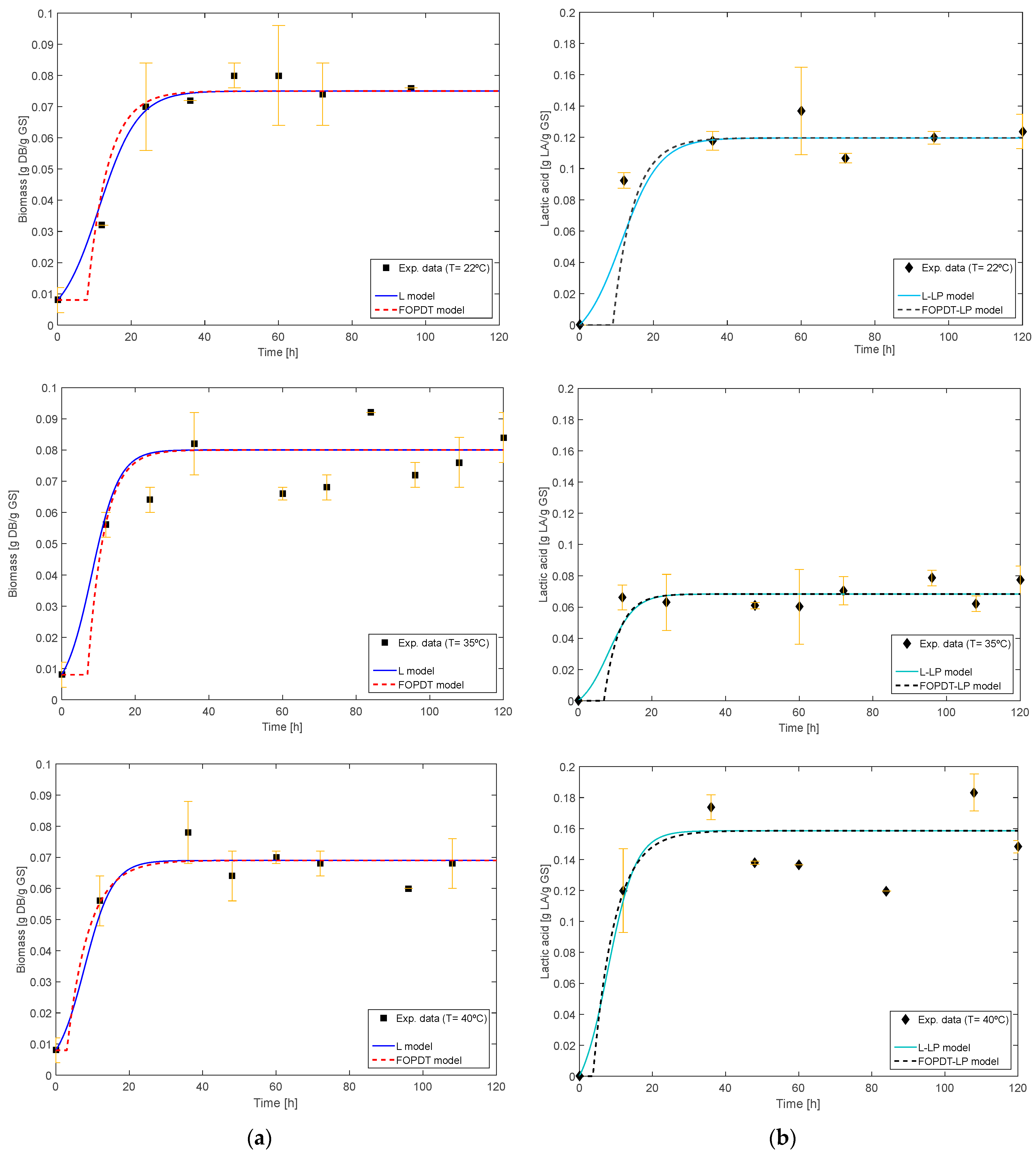
| Ambiental Conditions | R. oryzae Growth | Lactic Acid Production | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Logistic Model | FOPDT Model | L-LPwDT Model | FOPDT-LPwDT Model | ||||||||||
| RH | T | X0 | Xmax | µmax | R2 | tp | t0 | R2 | Yp/x | R2 | Yp/x | td | R2 |
| 50 | 22 | 0.008 ± 0.002 | 0.075 ± 0.002 | 0.189 ± 0.006 | 96.15 | 5.50 ± 0.14 | 9.00 ± 0.27 | 97.36 | 1.785 ± 0.053 | 82.45 | 1.785 ± 0.053 | 0 | 79.23 |
| 35 | 0.080 ± 0.002 | 0.270 ± 0.008 | 82.77 | 4.50 ± 0.12 | 7.00 ± 0.20 | 83.55 | 0.95 ± 0.03 | 83.76 | 0.95 ± 0.03 | 0 | 83.92 | ||
| 40 | 0.069 ± 0.002 | 0.260 ± 0.008 | 89.38 | 6.00 ± 0.17 | 3.00 ± 0.08 | 91.60 | 2.60 ± 0.13 | 84.33 | 2.60 ± 0.13 | 0.70 ± 0.02 | 84.40 | ||
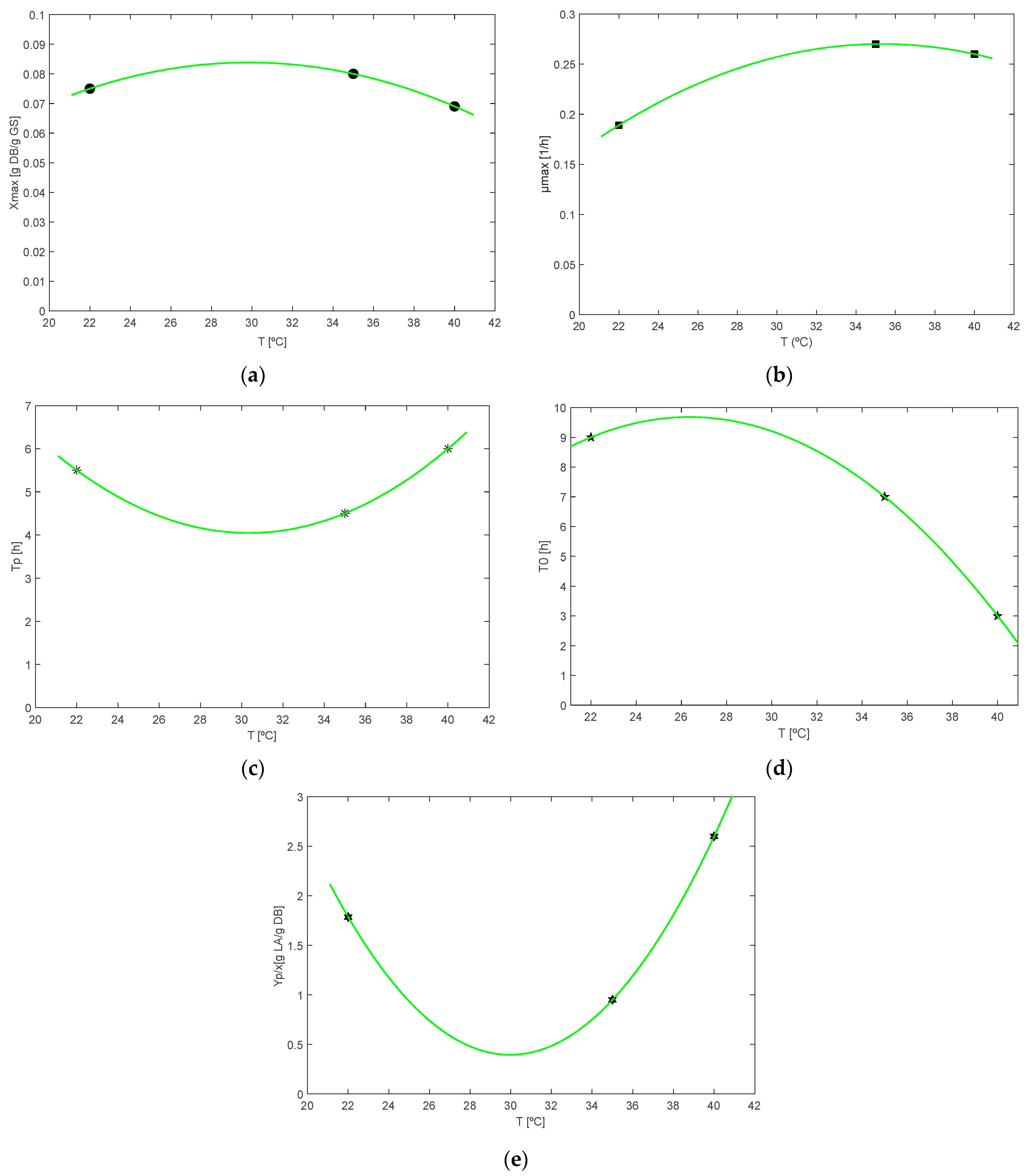
| Parameters | Fitted Polynomial | R2 |
|---|---|---|
| Xmax | Xmax (T) = −0.00014 T2 + 0.00857 T − 0.04403 | 100 |
| µmax | μmax (T) = −0.00046 T2 + 0.0323 T − 0.3002 | 100 |
| tp | tp (T) = 0.021 T2 − 1.271 T + 23.32 | 100 |
| t0 | t0 (T) = −0.0359 T2 + 1.892 T − 15.26 | 81.76 |
| Yp/x | Yp/x (T) = 0.0219 T2 − 1.313 T + 20.06 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groff, M.C.; Noriega, S.E.; Gil, R.M.; Pantano, N.; Scaglia, G. Dynamic Optimization of Lactic Acid Production from Grape Stalk Solid-State Fermentation with Rhizopus oryzae Applying a Variable Temperature Profile. Fermentation 2024, 10, 101. https://doi.org/10.3390/fermentation10020101
Groff MC, Noriega SE, Gil RM, Pantano N, Scaglia G. Dynamic Optimization of Lactic Acid Production from Grape Stalk Solid-State Fermentation with Rhizopus oryzae Applying a Variable Temperature Profile. Fermentation. 2024; 10(2):101. https://doi.org/10.3390/fermentation10020101
Chicago/Turabian StyleGroff, María Carla, Sandra Edith Noriega, Rocío Mariel Gil, Nadia Pantano, and Gustavo Scaglia. 2024. "Dynamic Optimization of Lactic Acid Production from Grape Stalk Solid-State Fermentation with Rhizopus oryzae Applying a Variable Temperature Profile" Fermentation 10, no. 2: 101. https://doi.org/10.3390/fermentation10020101
APA StyleGroff, M. C., Noriega, S. E., Gil, R. M., Pantano, N., & Scaglia, G. (2024). Dynamic Optimization of Lactic Acid Production from Grape Stalk Solid-State Fermentation with Rhizopus oryzae Applying a Variable Temperature Profile. Fermentation, 10(2), 101. https://doi.org/10.3390/fermentation10020101