

Source-Separated Industrial Wastewater Is a Candidate for Biogas Production through Anaerobic Digestion

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Selection and Collection
2.2. Biomethane Potential Tests
2.3. Semi-Continuous Digestion Tests
2.4. Sequencing of the Microbial Community
2.5. Analysis
2.6. Statistics
3. Results
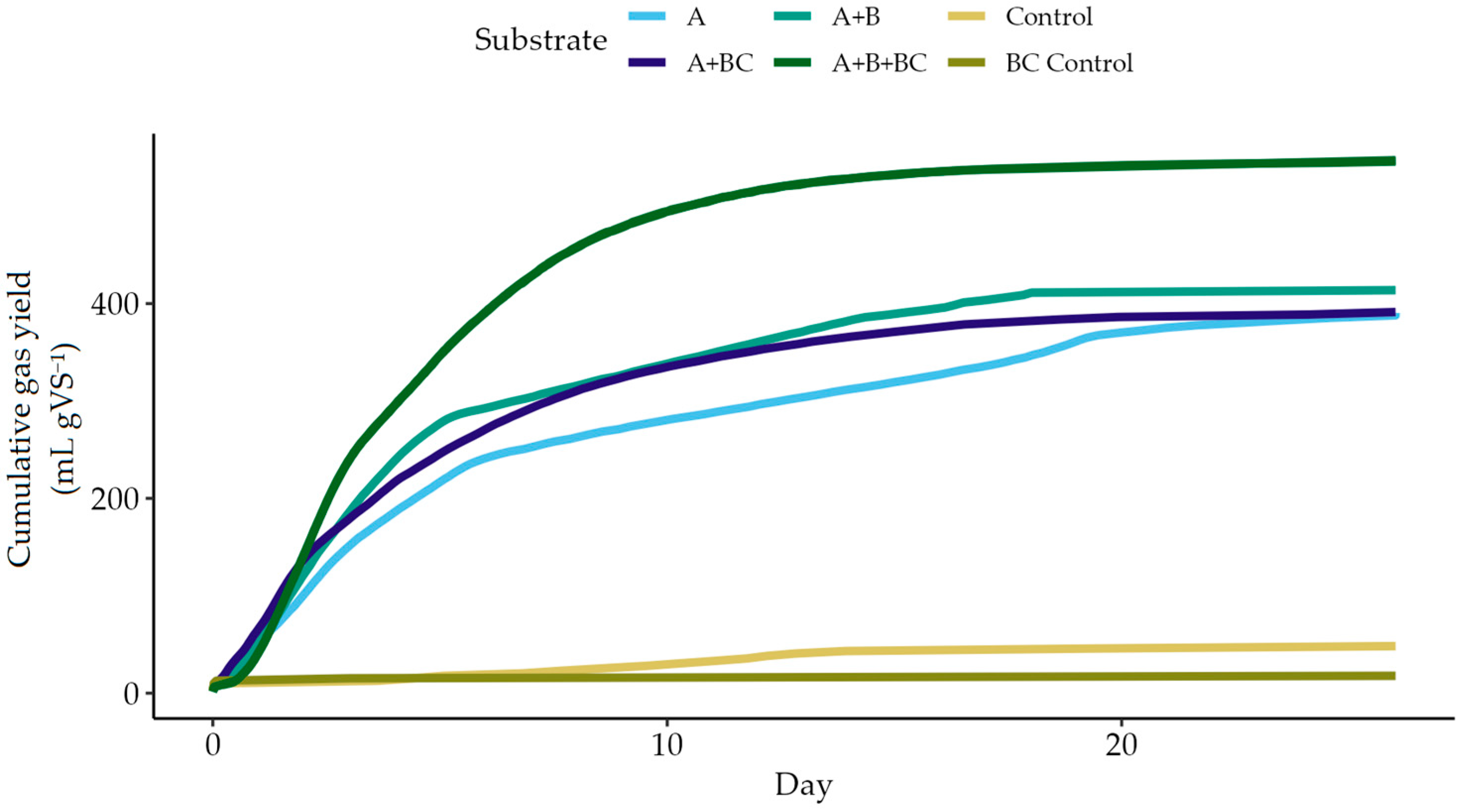
3.1. Biomethane Production
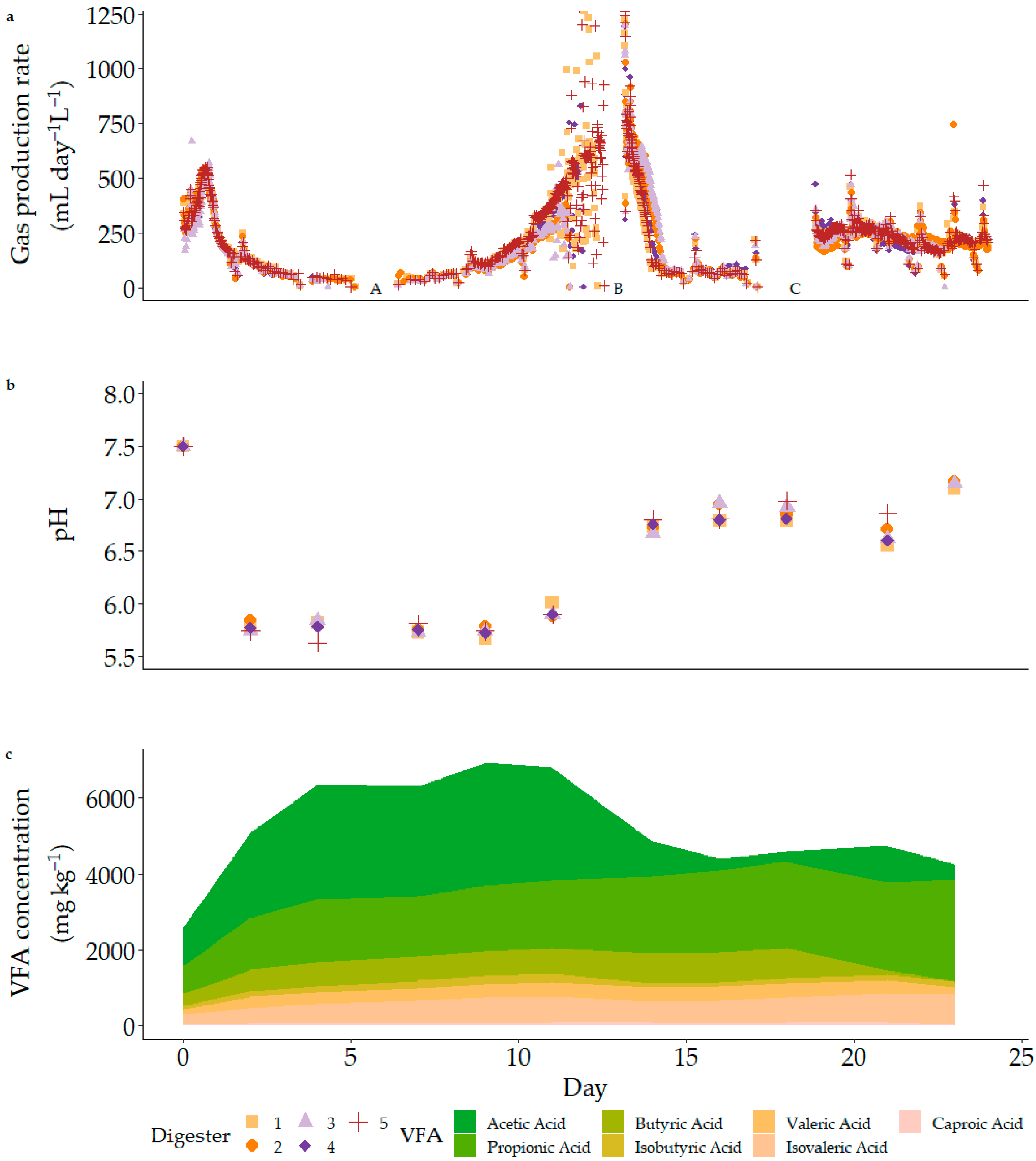
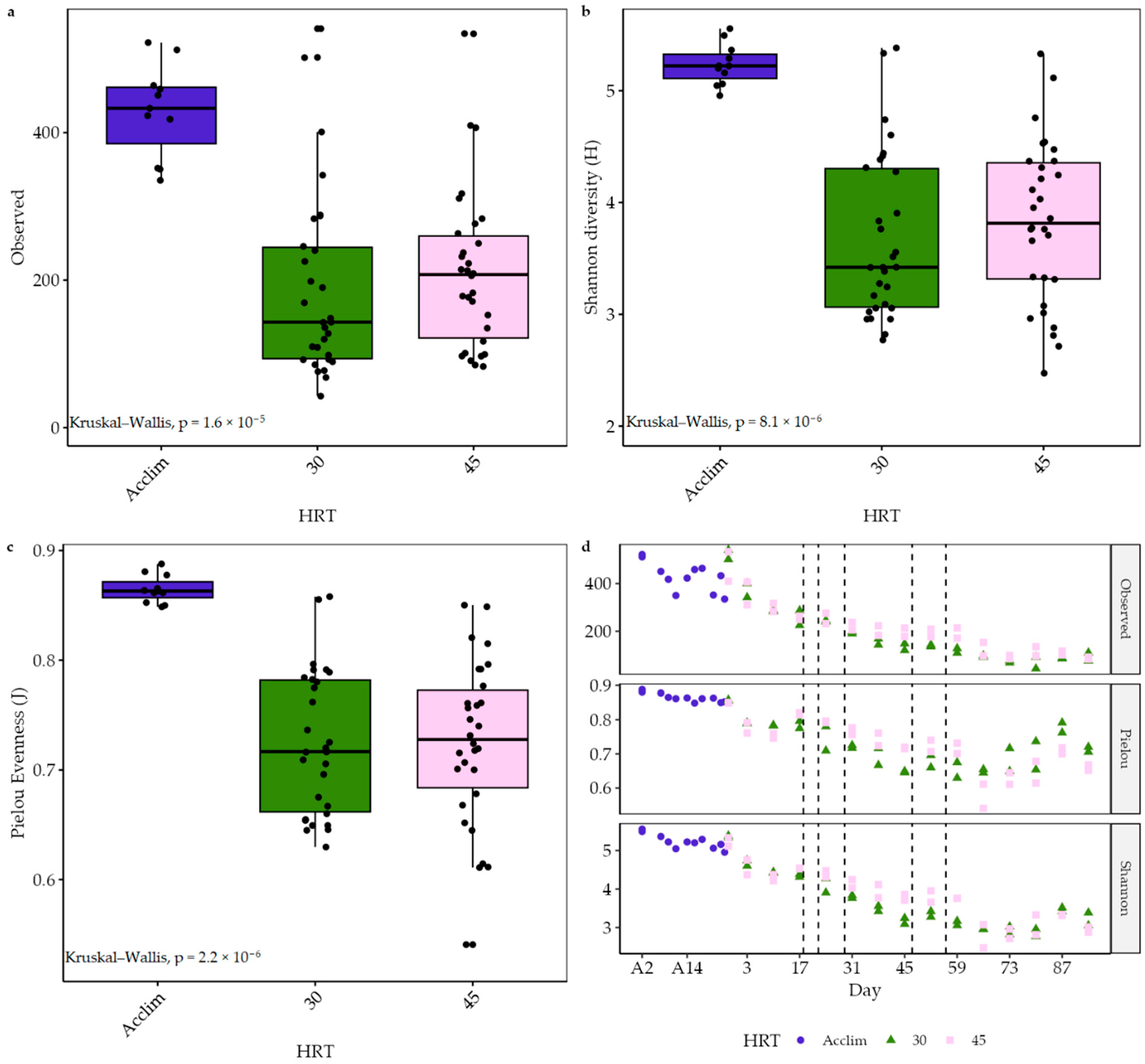
3.2. Acclimatisation
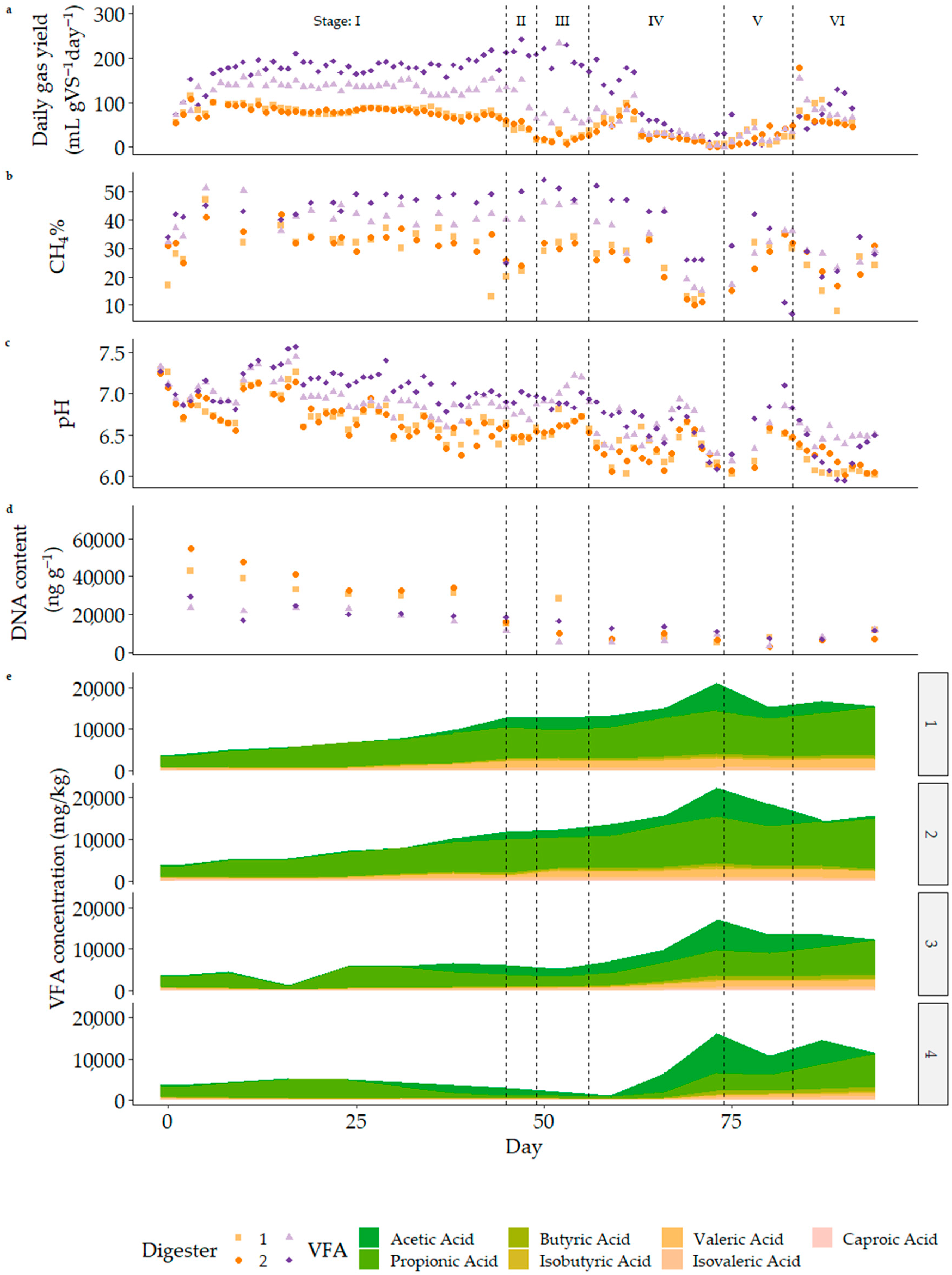
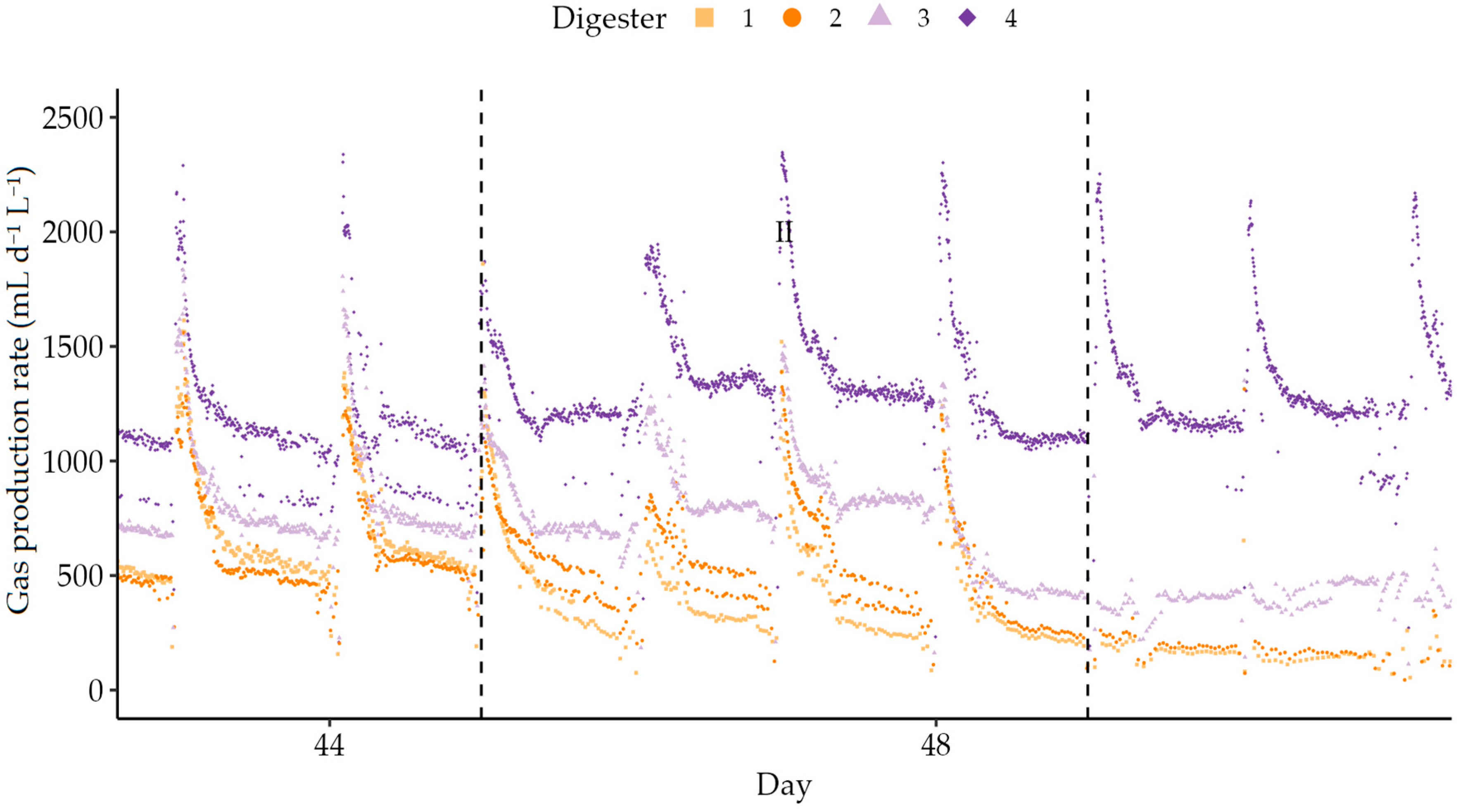
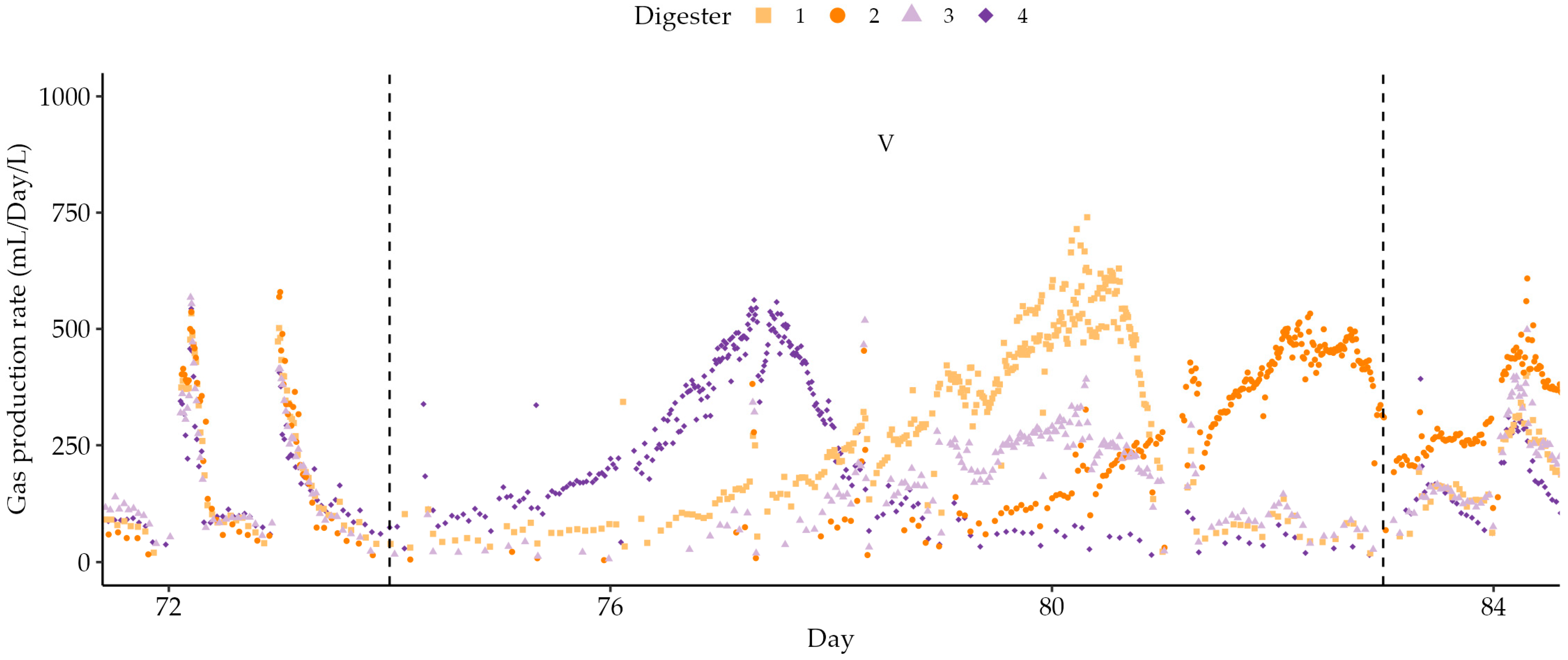
3.3. Semi-Continuous Operation
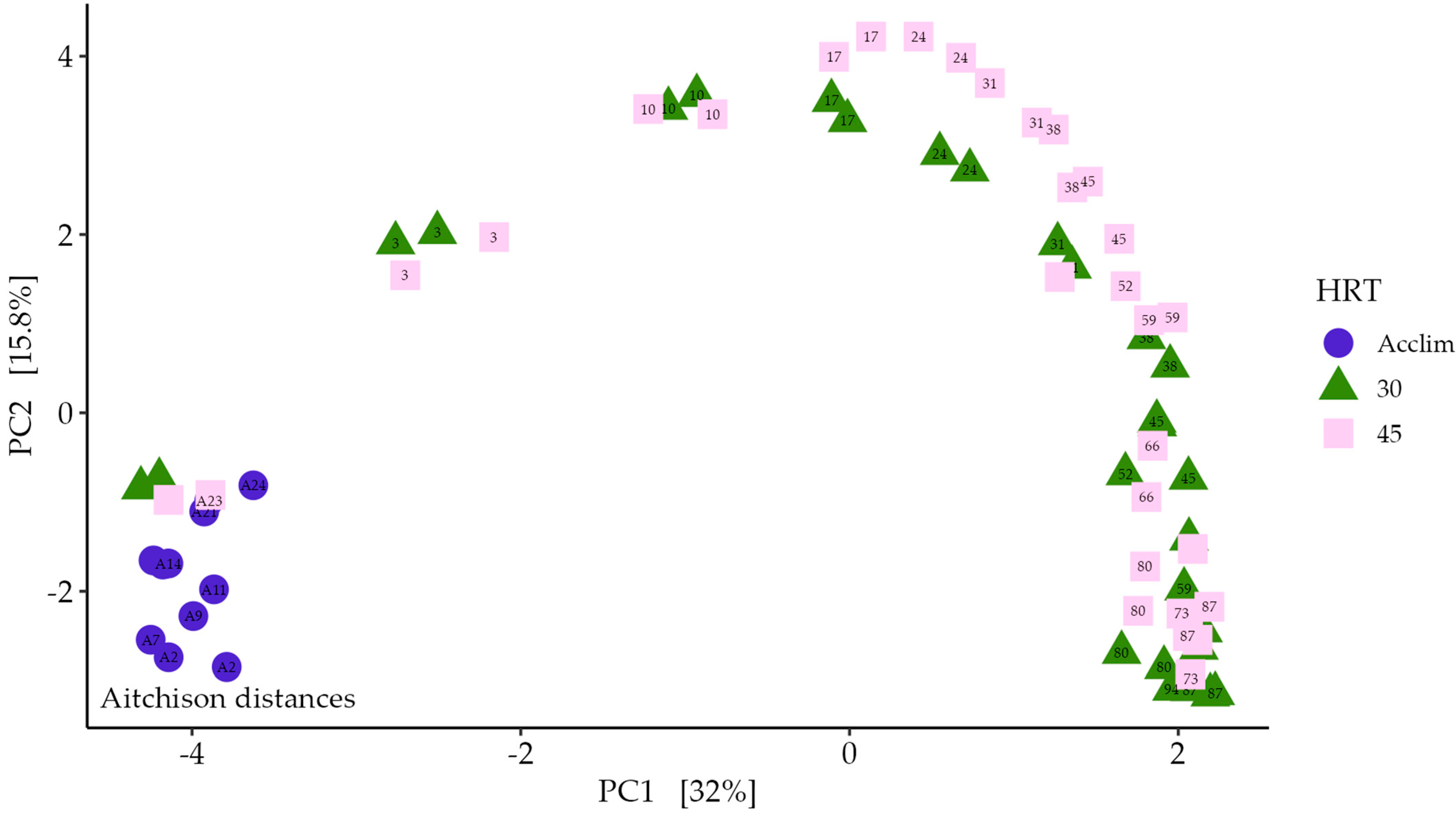
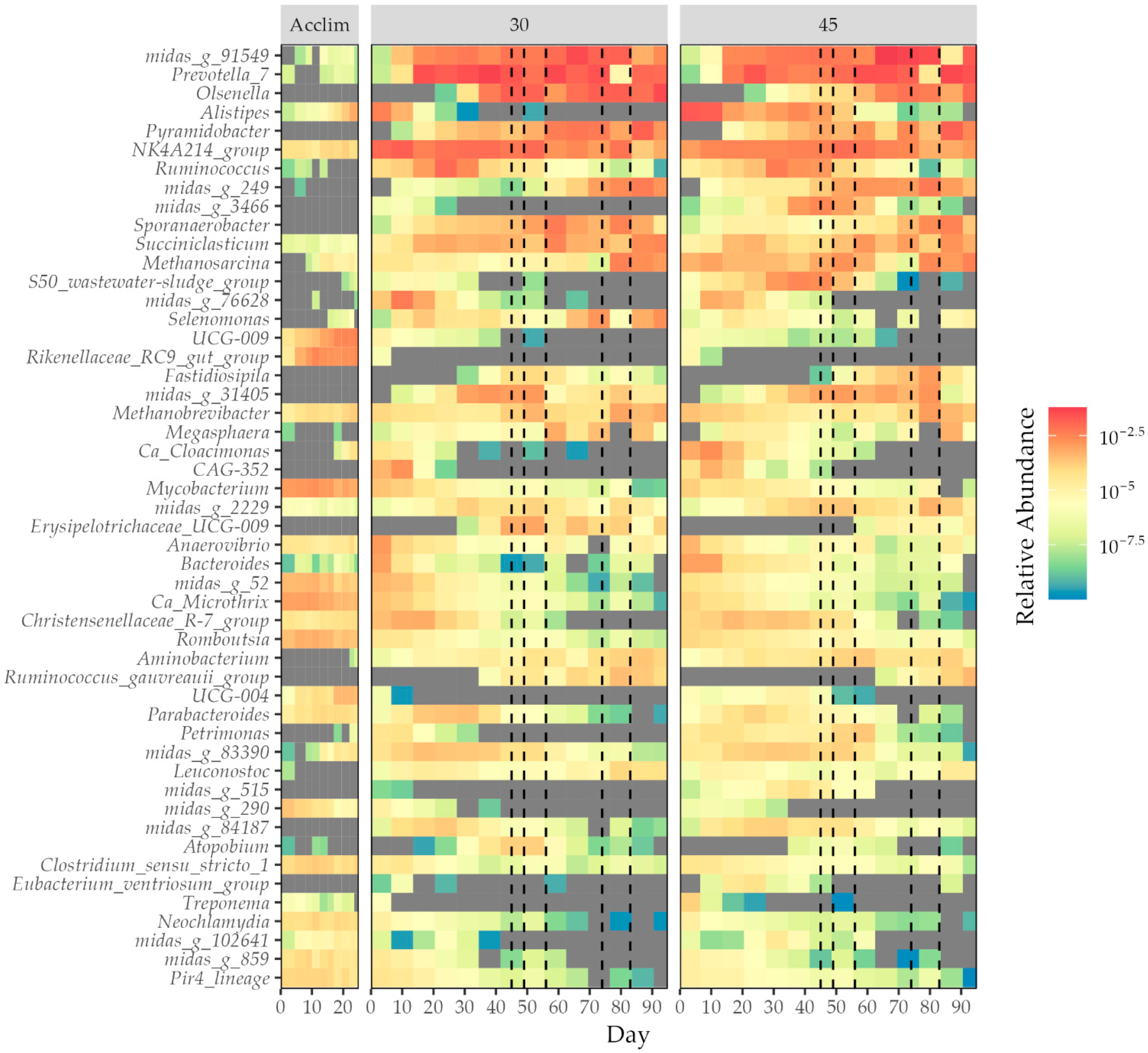
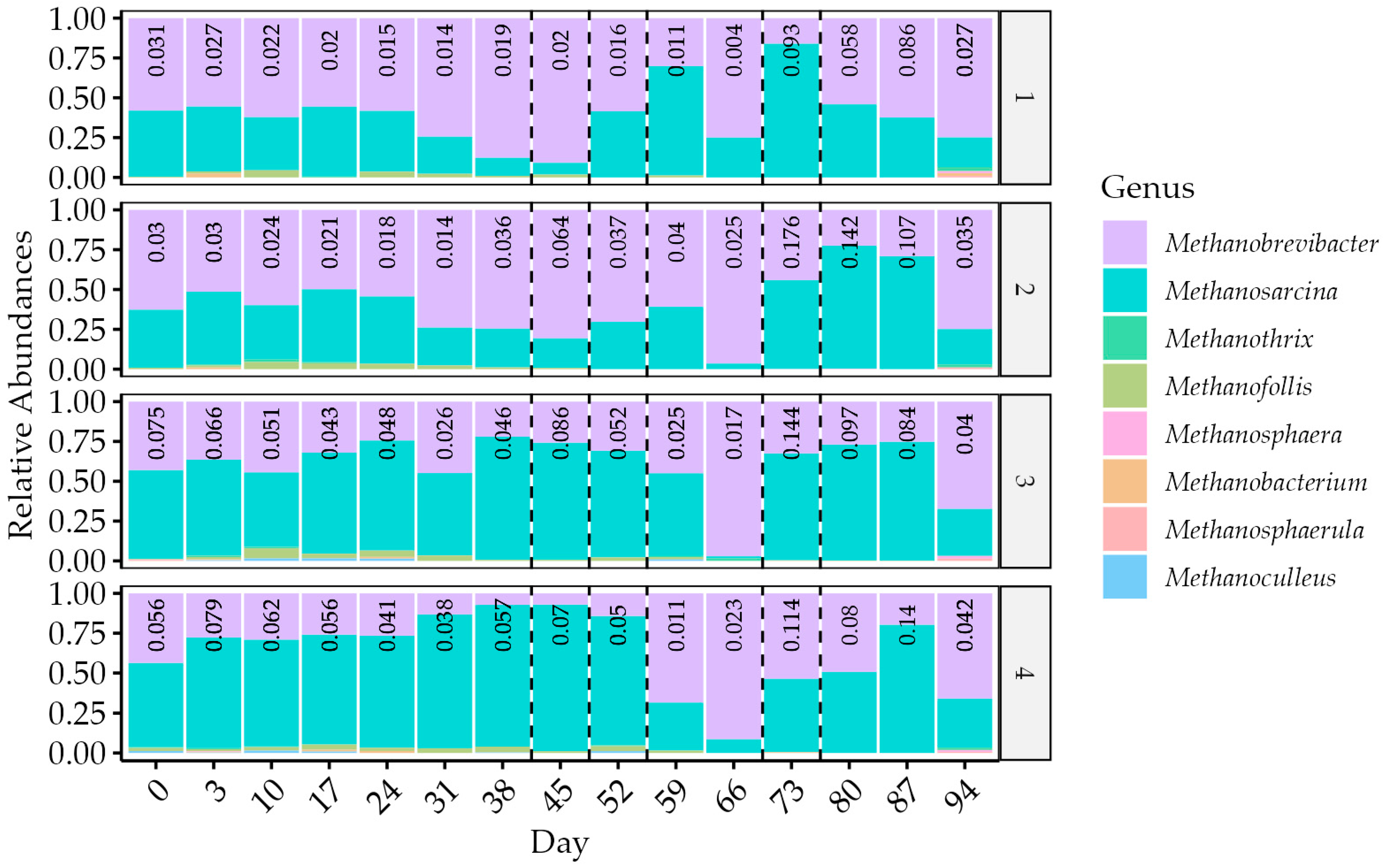
3.4. Changes in the Microbial Community
4. Discussion
4.1. Source-Separated Wastewater Potential
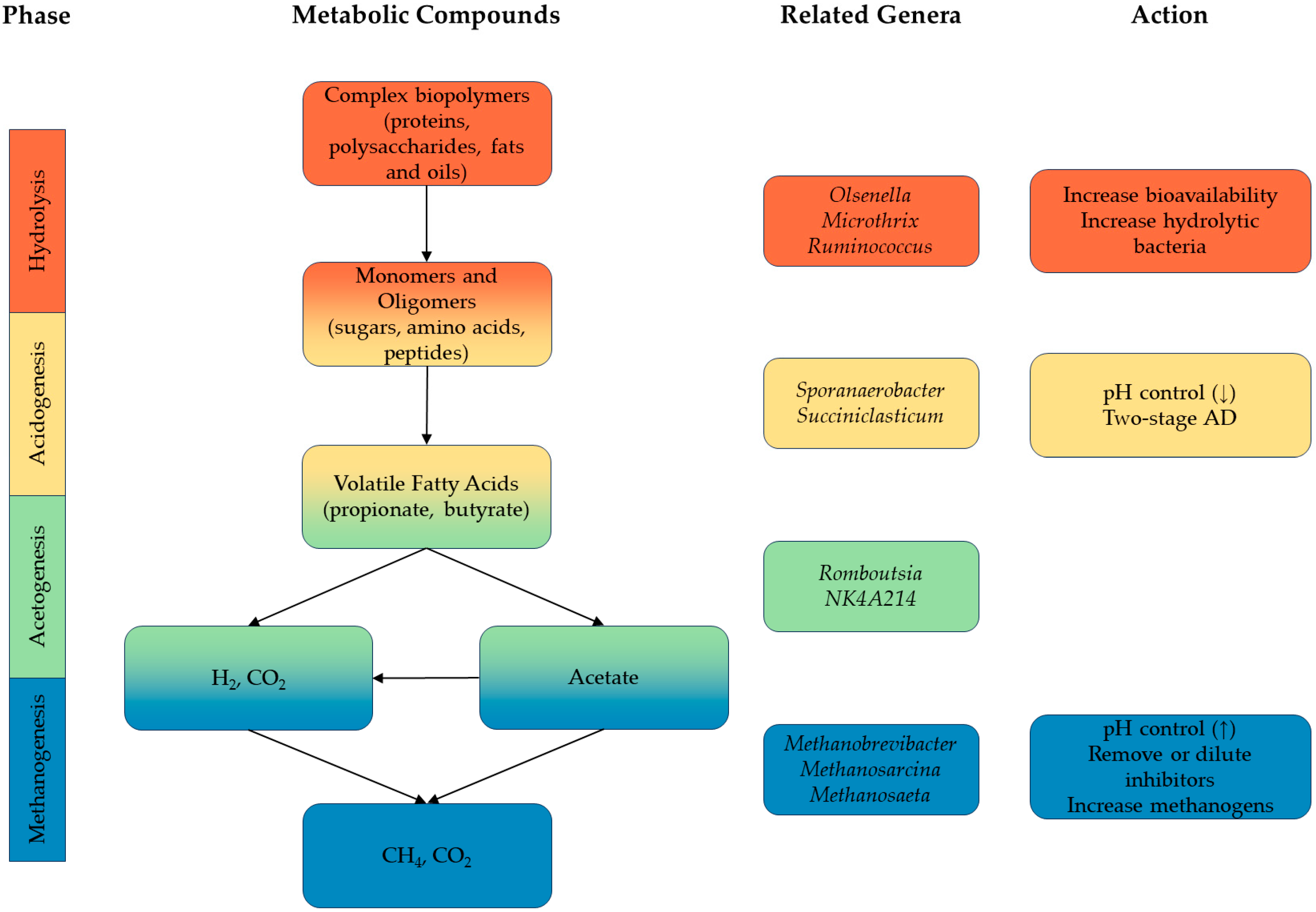
4.2. Microbial Community
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elliott, J.A.K.; Ball, A.S. Selection of industrial trade waste resource recovery technologies—A systematic review. Resources 2021, 10, 29. [Google Scholar] [CrossRef]
- Bywater, A.; Kusch-Brandt, S. Exploring farm anaerobic digester economic viability in a time of policy change in the UK. Processes 2022, 10, 212. [Google Scholar] [CrossRef]
- Hu, Y.; Cai, X.; Du, R.; Yang, Y.; Rong, C.; Qin, Y.; Li, Y.Y. A review on anaerobic membrane bioreactors for enhanced valorization of urban organic wastes: Achievements, limitations, energy balance and future perspectives. Sci. Total Environ. 2022, 820, 153284. [Google Scholar] [CrossRef]
- Economou, E.A.; Dimitropoulou, G.; Prokopidou, N.; Dalla, I.; Sfetsas, T. Anaerobic digestion remediation in three full-scale biogas plants through supplement additions. Methane 2023, 2, 265–278. [Google Scholar] [CrossRef]
- Tsigkou, K.; Zagklis, D.; Parasoglou, M.; Zafiri, C.; Kornaros, M. Proposed protocol for rate-limiting step determination during anaerobic digestion of complex substrates. Bioresour. Technol. 2022, 361, 127660. [Google Scholar] [CrossRef]
- Zhang, C.; Xiao, G.; Peng, L.; Su, H.; Tan, T. The anaerobic co-digestion of food waste and cattle manure. Bioresour. Technol. 2013, 129, 170–176. [Google Scholar] [CrossRef]
- Cavinato, C.; Bolzonella, D.; Pavan, P.; Fatone, F.; Cecchi, F. Mesophilic and thermophilic anaerobic co-digestion of waste activated sludge and source sorted biowaste in pilot- and full-scale reactors. Renew. Energy 2013, 55, 260–265. [Google Scholar] [CrossRef]
- Kim, J.K.; Oh, B.R.; Chun, Y.N.; Kim, S.W. Effects of temperature and hydraulic retention time on anaerobic digestion of food waste. J. Biosci. Bioeng. 2006, 102, 328–332. [Google Scholar] [CrossRef]
- Krohn, C.; Khudur, L.; Dias, D.A.; van den Akker, B.; Rees, C.A.; Crosbie, N.D.; Surapaneni, A.; O’Carroll, D.M.; Stuetz, R.M.; Batstone, D.J.; et al. The role of microbial ecology in improving the performance of anaerobic digestion of sewage sludge. Front. Microbiol. 2022, 13, 1079136. [Google Scholar] [CrossRef]
- Khafipour, A.; Jordaan, E.M.; Flores-Orozco, D.; Khafipour, E.; Levin, D.B.; Sparling, R.; Cicek, N. Response of microbial community to induced failure of anaerobic digesters through overloading with propionic acid followed by process recovery. Front. Bioeng. Biotechnol. 2020, 8, 604838. [Google Scholar] [CrossRef]
- Ziganshin, A.M.; Schmidt, T.; Lv, Z.; Liebetrau, J.; Richnow, H.H.; Kleinsteuber, S.; Nikolausz, M. Reduction of the hydraulic retention time at constant high organic loading rate to reach the microbial limits of anaerobic digestion in various reactor systems. Bioresour. Technol. 2016, 217, 62–71. [Google Scholar] [CrossRef]
- Stankiewicz, A.; Moulijn, J.A. Process intensification. Ind. Eng. Chem. Res. 2002, 41, 1920–1924. [Google Scholar] [CrossRef]
- Srivastava, S.; Pandey, V.K.; Fatima, A.; Sachin, V.M.; Pandey, S.; Singh, R.; Dar, A.H.; Dhillon, B. A literature review on process intensification: An innovative and sustainable food processing method. Appl. Food Res. 2024, 4, 100363. [Google Scholar] [CrossRef]
- Dareioti, M.A.; Kornaros, M. Effect of hydraulic retention time (HRT) on the anaerobic co-digestion of agro-industrial wastes in a two-stage CSTR system. Bioresour. Technol. 2014, 167, 407–415. [Google Scholar] [CrossRef]
- Fitamo, T.; Boldrin, A.; Boe, K.; Angelidaki, I.; Scheutz, C. Co-digestion of food and garden waste with mixed sludge from wastewater treatment in continuously stirred tank reactors. Bioresour. Technol. 2016, 206, 245–254. [Google Scholar] [CrossRef]
- Yu, Q.; Feng, L.; Zhen, X. Effects of organic loading rate and temperature fluctuation on the microbial community and performance of anaerobic digestion of food waste. Environ. Sci. Pollut. Res. Int. 2021, 28, 13176–13187. [Google Scholar] [CrossRef]
- Nguyen, A.Q.; Nguyen, L.N.; Phan, H.V.; Galway, B.; Bustamante, H.; Nghiem, L.D. Effects of operational disturbance and subsequent recovery process on microbial community during a pilot-scale anaerobic co-digestion. Int. Biodeterior. Biodegrad. 2019, 138, 70–77. [Google Scholar] [CrossRef]
- Kazimierowicz, J.; Dzienis, L.; Dębowski, M.; Zieliński, M. Optimisation of methane fermentation as a valorisation method for food waste products. Biomass Bioenergy 2021, 144, 105913. [Google Scholar] [CrossRef]
- Akunna, J.C. Anaerobic Waste-Wastewater Treatment and Biogas Plants: A Practical Handbook; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Nguyen, V.T.; Beyer, E.; Neumann, J.; Awe, D.; Pfeiffer, W.; Tranckner, J. Anaerobic treatment of residuals from tanks transporting food and fodder. Environ. Sci. Pollut. Res. Int. 2019, 26, 32698–32707. [Google Scholar] [CrossRef]
- Jensen, P.D.; Sullivan, T.; Carney, C.; Batstone, D.J. Analysis of the potential to recover energy and nutrient resources from cattle slaughterhouses in Australia by employing anaerobic digestion. Appl. Energy 2014, 136, 23–31. [Google Scholar] [CrossRef]
- Weber, K.; Quicker, P. Properties of biochar. Fuel 2018, 217, 240–261. [Google Scholar] [CrossRef]
- Patel, S.; Kundu, S.; Halder, P.; Ratnnayake, N.; Marzbali, M.H.; Aktar, S.; Selezneva, E.; Paz-Ferreiro, J.; Surapaneni, A.; de Figueiredo, C.C.; et al. A critical literature review on biosolids to biochar: An alternative biosolids management option. Rev. Environ. Sci. Bio/Technol. 2020, 19, 807–841. [Google Scholar] [CrossRef]
- Deena, S.R.; Vickram, A.S.; Manikandan, S.; Subbaiya, R.; Karmegam, N.; Ravindran, B.; Chang, S.W.; Awasthi, M.K. Enhanced biogas production from food waste and activated sludge using advanced techniques—A review. Bioresour. Technol. 2022, 355, 127234. [Google Scholar] [CrossRef]
- Hassan, S.; Ngo, T.; Khudur, L.S.; Krohn, C.; Dike, C.C.; Hakeem, I.G.; Shah, K.; Surapaneni, A.; Ball, A.S. Biosolids-derived biochar improves biomethane production in the anaerobic digestion of chicken manure. Resources 2023, 12, 123. [Google Scholar] [CrossRef]
- Ngo, T.; Khudur, L.S.; Hakeem, I.G.; Shah, K.; Surapaneni, A.; Ball, A.S. Wood biochar enhances the valorisation of the anaerobic digestion of chicken manure. Clean. Technol. 2022, 4, 420–439. [Google Scholar] [CrossRef]
- Ma, J.; Chen, F.; Xue, S.; Pan, J.; Khoshnevisan, B.; Yang, Y.; Liu, H.; Qiu, L. Improving anaerobic digestion of chicken manure under optimized biochar supplementation strategies. Bioresour. Technol. 2021, 325, 124697. [Google Scholar] [CrossRef]
- Wang, G.; Chu, Y.; Zhu, J.; Sheng, L.; Liu, G.; Xing, Y.; Fu, P.; Li, Q.; Chen, R. Multi-faceted influences of biochar addition on swine manure digestion under tetracycline antibiotic pressure. Bioresour. Technol. 2022, 346, 126352. [Google Scholar] [CrossRef]
- Shen, R.; Jing, Y.; Feng, J.; Zhao, L.; Yao, Z.; Yu, J.; Chen, J.; Chen, R. Simultaneous carbon dioxide reduction and enhancement of methane production in biogas via anaerobic digestion of cornstalk in continuous stirred-tank reactors: The influences of biochar, environmental parameters, and microorganisms. Bioresour. Technol. 2021, 319, 124146. [Google Scholar] [CrossRef]
- Shen, R.; Jing, Y.; Feng, J.; Luo, J.; Yu, J.; Zhao, L. Performance of enhanced anaerobic digestion with different pyrolysis biochars and microbial communities. Bioresour. Technol. 2020, 296, 122354. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Peng, H.; Adhikari, S.; Higgins, B.; Roy, P.; Dai, W.; Shi, X. Enhancement of biogas production from wastewater sludge via anaerobic digestion assisted with biochar amendment. Bioresour. Technol. 2020, 309, 123368. [Google Scholar] [CrossRef]
- Wang, G.; Zhu, J.; Xing, Y.; Yin, Y.; Li, Y.; Li, Q.; Chen, R. When dewatered swine manure-derived biochar meets swine wastewater in anaerobic digestion: A win-win scenario towards highly efficient energy recovery and antibiotic resistance genes attenuation for swine manure management. Sci. Total Environ. 2022, 803, 150126. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Nieto, A.; Mendez, A.; Askeland, M.P.J.; Gasco, G. Biochar from biosolids pyrolysis: A review. Int. J. Environ. Res. Public. Health 2018, 15, 956. [Google Scholar] [CrossRef] [PubMed]
- Chagas, J.K.M.; Figueiredo, C.C.; Silva, J.D.; Shah, K.; Paz-Ferreiro, J. Long-term effects of sewage sludge-derived biochar on the accumulation and availability of trace elements in a tropical soil. J. Environ. Qual. 2021, 50, 264–277. [Google Scholar] [CrossRef]
- de la Rosa, J.M.; Paneque, M.; Miller, A.Z.; Knicker, H. Relating physical and chemical properties of four different biochars and their application rate to biomass production of Lolium perenne on a calcic cambisol during a pot experiment of 79 days. Sci. Total Environ. 2014, 499, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, J.; Wang, Y.; Yang, C. Impacts of different biochar types on the anaerobic digestion of sewage sludge. RSC Adv. 2019, 9, 42375–42386. [Google Scholar] [CrossRef]
- Chiappero, M.; Berruti, F.; Masek, O.; Fiore, S. Semi-continuous anaerobic digestion of mixed wastewater sludge with biochar addition. Bioresour. Technol. 2021, 340, 125664. [Google Scholar] [CrossRef]
- Kraatz, M.; Wallace, R.J.; Svensson, L. Olsenella umbonata sp. nov., a microaerotolerant anaerobic lactic acid bacterium from the sheep rumen and pig jejunum, and emended descriptions of Olsenella, Olsenella uli and Olsenella profusa. Int. J. Syst. Evol. Microbiol. 2011, 61, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Ziels, R.M.; Karlsson, A.; Beck, D.A.C.; Ejlertsson, J.; Yekta, S.S.; Bjorn, A.; Stensel, H.D.; Svensson, B.H. Microbial community adaptation influences long-chain fatty acid conversion during anaerobic codigestion of fats, oils, and grease with municipal sludge. Water Res. 2016, 103, 372–382. [Google Scholar] [CrossRef]
- Rossetti, S.; Tomei, M.C.; Nielsen, P.H.; Tandoi, V. “Microthrix parvicella”, a filamentous bacterium causing bulking and foaming in activated sludge systems: A review of current knowledge. FEMS Microbiol. Rev. 2005, 29, 49–64. [Google Scholar] [CrossRef]
- Warren, R.A.J. Microbial hydrolysis of polysaccharides. Annu. Rev. Microbiol. 1996, 50, 183–212. [Google Scholar] [CrossRef]
- Hernandez-Eugenio, G. Sporanaerobacter acetigenes gen. nov., sp. nov., a novel acetogenic, facultatively sulfur-reducing bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 1217–1223. [Google Scholar] [PubMed]
- Van Gylswyk, N.O. Succiniclasticum ruminis gen. nov., sp. nov., a ruminal bacterium converting succinate to propionate as the sole energy-yielding mechanism. Int. J. Syst. Bacteriol. 1995, 45, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, J.; Fuentes, S.; Grievink, W.; van Niftrik, L.; Tindall, B.J.; Timmerman, H.M.; Rijkers, G.T.; Smidt, H. Characterization of Romboutsia ilealis gen. nov., sp. nov., isolated from the gastro-intestinal tract of a rat, and proposal for the reclassification of five closely related members of the genus Clostridium into the genera Romboutsia gen. nov., Intestinibacter gen. nov., Terrisporobacter gen. nov. and Asaccharospora gen. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 1600–1616. [Google Scholar] [PubMed]
- Liu, C.; Wu, H.; Liu, S.; Chai, S.; Meng, Q.; Zhou, Z. Dynamic alterations in yak rumen bacteria community and metabolome characteristics in response to feed type. Front. Microbiol. 2019, 10, 1116. [Google Scholar] [CrossRef]
- Garcia, J.L.; Patel, B.K.; Ollivier, B. Taxonomic, phylogenetic, and ecological diversity of methanogenic archaea. Anaerobe 2000, 6, 205–226. [Google Scholar] [CrossRef]
- Riesco, R.; Rose, J.J.A.; Batinovic, S.; Petrovski, S.; Sanchez-Juanes, F.; Seviour, R.J.; Goodfellow, M.; Trujillo, M.E. Gordonia pseudamarae sp. nov., a home for novel Actinobacteria isolated from stable foams on activated sludge wastewater treatment plants. Int. J. Syst. Evol. Microbiol. 2022, 72, 005547. [Google Scholar] [CrossRef]
- Elliott, J.A.K.; Ball, A.S.; Shah, K. Investigations into valorisation of trade wastewater for biomethane production. Heliyon 2023, 9, e13309. [Google Scholar] [CrossRef]
- Baird, R.B.; Eaton, A.D.; Rice, E.W. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Hach Company. Oxygen Demand, Chemical. 2021. Available online: https://au.hach.com/asset-get.download.jsa?id=7639983816 (accessed on 13 December 2021).
- Kundu, S.; Patel, S.; Halder, P.; Patel, T.; Hedayati Marzbali, M.; Pramanik, B.K.; Paz-Ferreiro, J.; de Figueiredo, C.C.; Bergmann, D.; Surapaneni, A.; et al. Removal of PFASs from biosolids using a semi-pilot scale pyrolysis reactor and the application of biosolids derived biochar for the removal of PFASs from contaminated water. Environ. Sci. Water Res. Technol. 2021, 7, 638–649. [Google Scholar] [CrossRef]
- Anaero Technology. Available online: www.anaerotech.com/bmp-reactors (accessed on 15 September 2022).
- Mpofu, A.B.; Kibangou, V.A.; Kaira, W.M.; Oyekola, O.O.; Welz, P.J. Anaerobic co-digestion of tannery and slaughterhouse wastewater for solids reduction and resource recovery: Effect of sulfate concentration and inoculum to substrate ratio. Energies 2021, 14, 2491. [Google Scholar] [CrossRef]
- Latifi, P.; Karrabi, M.; Danesh, S. Anaerobic co-digestion of poultry slaughterhouse wastes with sewage sludge in batch-mode bioreactors (effect of inoculum-substrate ratio and total solids). Renew. Sustain. Energy Rev. 2019, 107, 288–296. [Google Scholar] [CrossRef]
- Yang, S.; Chen, Z.; Wen, Q. Impacts of biochar on anaerobic digestion of swine manure: Methanogenesis and antibiotic resistance genes dissemination. Bioresour. Technol. 2021, 324, 124679. [Google Scholar] [CrossRef]
- Jimenez, J.; Charnier, C.; Kouas, M.; Latrille, E.; Torrijos, M.; Harmand, J.; Patureau, D.; Sperandio, M.; Morgenroth, E.; Beline, F.; et al. Modelling hydrolysis: Simultaneous versus sequential biodegradation of the hydrolysable fractions. Waste Manag. 2020, 101, 150–160. [Google Scholar] [CrossRef]
- Torrijos, M. Assessment of BMP and Kinetics in 6l-Batch Reactors with Successive Feedings. In Proceedings of the Workshop on the Conundrum of Biomethane Potential Tests, Leysin, Switzerland, 10–12 June 2015. [Google Scholar]
- BPC Instruments. Bioreactor Simulator. 2019. Available online: https://bpcinstruments.com/ (accessed on 12 July 2022).
- Thermo Fisher Scientific Inc. Dionex Onguard ii Cartridges 031688. 2017. Available online: https://assets.thermofisher.com/TFS-Assets%2FCMD%2Fmanuals%2FMan-031688-IC-OnGuard-II-Cartridges-Man031688-EN.pdf (accessed on 20 April 2023).
- Chantarasukon, C.; Tukkeeree, S.; Rohrer, J. Determination of Organic Acids in Wastewater Using Ion-Exclusion Chromatography and On-Line Carbonate Removal; Thermo Fisher Scientific Inc.: Waltham, MA, USA, 2016. [Google Scholar]
- Voelklein, M.A.; O’ Shea, R.; Jacob, A.; Murphy, J.D. Role of trace elements in single and two-stage digestion of food waste at high organic loading rates. Energy 2017, 121, 185–192. [Google Scholar] [CrossRef]
- Industrial Scientific. Available online: https://www.indsci.com/en/gas-detectors/multi/mx6-ibrid (accessed on 12 July 2022).
- Qiagen. Dneasy® Powersoil® Pro Kit Handbook. 2023. Available online: https://www.qiagen.com/us/resources/download.aspx?id=9bb59b74-e493-4aeb-b6c1-f660852e8d97&lang=en (accessed on 20 August 2023).
- Invitrogen. Comparison of Fluorescence-Based Quantitation with UV Absorbance Measurements; ThermoFisher Scientific: Waltham, MA, USA, 2018; Volume COL22609 0618. [Google Scholar]
- Invitrogen. Qubit™ 4 Fluorometer; ThermoFisher Scientific: Waltham, MA, USA, 2021; Volume MAN0017209. [Google Scholar]
- Mazzoli, L.; Munz, G.; Lotti, T.; Ramazzotti, M. A novel universal primer pair for prokaryotes with improved performances for anammox containing communities. Sci. Rep. 2020, 10, 15648. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- Illumina. 16s Metagenomic Sequencing Library Preparation; Illumina: San Diego, CA, USA, 2013. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. Dada2: High-resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using qiime 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with qiime 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Dueholm, M.K.D.; Nierychlo, M.; Andersen, K.S.; Rudkjobing, V.; Knutsson, S.; MIDAS Global Consortium; Albertsen, M.; Nielsen, P.H. Midas 4: A global catalogue of full-length 16s rRNA gene sequences and taxonomy for studies of bacterial communities in wastewater treatment plants. Nat. Commun. 2022, 13, 1908. [Google Scholar] [CrossRef] [PubMed]
- Dueholm, M.K.D.; Andersen, K.S.; Petersen, A.-K.C.; Rudkjøbing, V.; Nielsen, P.H. MiDAS 5: Global diversity of bacteria and archaea in anaerobic digesters. bioRxiv 2023. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An r package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Wilke, C. Cowplot: Streamlined Plot Theme and Plot Annotations for ‘ggplot2’, 1.1.1 ed.; 2020. Available online: https://CRAN.R-project.org/package=cowplot (accessed on 23 October 2023).
- Lahti, L.; Shetty, S. Microbiome r Package. 2012–2019. Available online: http://microbiome.github.io (accessed on 23 October 2023).
- Zeileis, A.; Fisher, J.C.; Hornik, K.; Ihaka, R.; McWhite, C.D.; Murrell, P.; Stauffer, R.; Wilke, C.O. Colorspace: A toolbox for manipulating and assessing colors and palettes. J. Stat. Softw. 2020, 96, 1–49. [Google Scholar] [CrossRef]
- Kassambara, A. Ggpubr: ‘Ggplot2’ Based Publication Ready Plots. 2023. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 23 October 2023).
- Wickham, H. Ggplot2; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Wickham, H.; Vaughan, D.; Girlich, M. Tidyr: Tidy messy data. J. Stat. Softw. 2014, 59, 1–23. [Google Scholar] [CrossRef]
- Müller, K.; Wickham, H. Tibble: Simple Data Frames. 2023. Available online: https://CRAN.R-project.org/package=tibble (accessed on 23 October 2023).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Sarkar, D. Lattice: Multivariate Data Visualization with r; Springer: New York, NY, USA, 2008. [Google Scholar]
- Simpson, G.L. Permute: Functions for Generating Restricted Permutations of Data. 2022. Available online: https://CRAN.R-project.org/package=permute (accessed on 23 October 2023).
- Wickham, H. Stringr: Simple, Consistent Wrappers for Common String Operations. 2022. Available online: https://CRAN.R-project.org/package=stringr (accessed on 23 October 2023).
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. Dplyr: A Grammar of Data Manipulation. 2023. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 23 October 2023).
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. 2023. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 23 October 2023).
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 23 October 2023).
- Hafner, S.D.; Fruteau de Laclos, H.; Koch, K.; Holliger, C. Improving inter-laboratory reproducibility in measurement of biochemical methane potential (BMP). Water 2020, 12, 1752. [Google Scholar] [CrossRef]
- Weinrich, S.; Schäfer, F.; Bochmann, G.; Liebetrau, J. Value of Batch Tests for Biogas Potential Analysis; IEA Bioenergy: Paris, France, 2018. [Google Scholar]
- Maya-Altamira, L.; Baun, A.; Angelidaki, I.; Schmidt, J.E. Influence of wastewater characteristics on methane potential in food-processing industry wastewaters. Water Res. 2008, 42, 2195–2203. [Google Scholar] [CrossRef]
- Hai, F.I.; Yamamoto, K. 4.16 membrane biological reactors. In Treatise on Water Science; Wilderer, P., Ed.; Elsevier: Oxford, UK, 2011; pp. 571–613. [Google Scholar]
- Casallas-Ojeda, M.R.; Marmolejo-Rebellón, L.F.; Torres-Lozada, P. Identification of factors and variables that influence the anaerobic digestion of municipal biowaste and food waste. Waste Biomass Valorization 2020, 12, 2889–2904. [Google Scholar] [CrossRef]
- Zhuang, H.; Amy Tan, G.-Y.; Jing, H.; Lee, P.-H.; Lee, D.-J.; Leu, S.-Y. Enhanced primary treatment for net energy production from sewage—The genetic clarification of substrate-acetate-methane pathway in anaerobic digestion. Chem. Eng. J. 2022, 431, 133416. [Google Scholar] [CrossRef]
- Bjornsson, L.; Murto, M.; Mattiasson, B. Evaluation of parameters for monitoring an anaerobic co-digestion process. Appl. Microbiol. Biotechnol. 2000, 54, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Angelidaki, I.; Zhang, Y. Microbial electrochemical monitoring of volatile fatty acids during anaerobic digestion. Environ. Sci. Technol. 2016, 50, 4422–4429. [Google Scholar] [CrossRef] [PubMed]
- Boe, K.; Batstone, D.J.; Steyer, J.P.; Angelidaki, I. State indicators for monitoring the anaerobic digestion process. Water Res. 2010, 44, 5973–5980. [Google Scholar] [CrossRef]
- García-Sandoval, J.P.; Méndez-Acosta, H.O.; González-Alvarez, V.; Schaum, A.; Alvarez, J. VFA robust control of an anaerobic digestion pilot plant: Experimental implementation. IFAC-PapersOnLine 2016, 49, 973–977. [Google Scholar] [CrossRef]
- Nielsen, H.B.; Ahring, B.K. Responses of the biogas process to pulses of oleate in reactors treating mixtures of cattle and pig manure. Biotechnol. Bioeng. 2006, 95, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Cavaleiro, A.J.; Pereira, M.A.; Alves, M. Enhancement of methane production from long chain fatty acid based effluents. Bioresour. Technol. 2008, 99, 4086–4095. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, J.; Verstraete, W.; Boon, N. Repeated pulse feeding induces functional stability in anaerobic digestion. Microb. Biotechnol. 2013, 6, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Peces, M.; Astals, S.; Mata-Alvarez, J. Response of a sewage sludge mesophilic anaerobic digester to short and long-term thermophilic temperature fluctuations. Chem. Eng. J. 2013, 233, 109–116. [Google Scholar] [CrossRef]
- Yu, L.; Li, R.; Delatolla, R.; Zhang, R.; Yang, X.; Peng, D. Natural continuous influent nitrifier immigration effects on nitrification and the microbial community of activated sludge systems. J. Environ. Sci 2018, 74, 159–167. [Google Scholar] [CrossRef]
- Peces, M.; Astals, S.; Jensen, P.D.; Clarke, W.P. Transition of microbial communities and degradation pathways in anaerobic digestion at decreasing retention time. New Biotechnol. 2021, 60, 52–61. [Google Scholar] [CrossRef]
- Zhu, L.; Wu, B.; Liu, Y.; Zhang, J.; Deng, R.; Gu, L. Strategy to enhance semi-continuous anaerobic digestion of food waste by combined use of calcium peroxide and magnetite. Water Res. 2022, 221, 118801. [Google Scholar] [CrossRef]
- García-Ruíz, M.J.; Castellano-Hinojosa, A.; Armato, C.; González-Martínez, A.; González-López, J.; Osorio, F. Biogas production and microbial community structure in a stable-stage of a two-stage anaerobic digester. AIChE J. 2019, 66, e16807. [Google Scholar] [CrossRef]
- Camarillo, R.; Rincón, J. Effect of inhibitory compounds on the anaerobic digestion performance of diluted wastewaters from the alimentary industry. J. Chem. Technol. Biotechnol. 2009, 84, 1615–1623. [Google Scholar] [CrossRef]
- Edwiges, T.; Bastos, J.A.; Lima Alino, J.H.; d’avila, L.; Frare, L.M.; Somer, J.G. Comparison of various pretreatment techniques to enhance biodegradability of lignocellulosic biomass for methane production. J. Environ. Chem. Eng. 2019, 7, 103495. [Google Scholar] [CrossRef]
- Di Berardino, S.; Costa, S.; Converti, A. Semi-continuous anaerobic digestion of a food industry wastewater in an anaerobic filter. Bioresour. Technol. 2000, 71, 261–266. [Google Scholar] [CrossRef]
- Chiu, S.L.H.; Lo, I.M.C. Reviewing the anaerobic digestion and co-digestion process of food waste from the perspectives on biogas production performance and environmental impacts. Environ. Sci. Pollut. Res. Int. 2016, 23, 24435–24450. [Google Scholar] [CrossRef] [PubMed]
- Borth, P.L.B.; Perin, J.K.H.; Torrecilhas, A.R.; Lopes, D.D.; Santos, S.C.; Kuroda, E.K.; Fernandes, F. Pilot-scale anaerobic co-digestion of food and garden waste: Methane potential, performance and microbial analysis. Biomass Bioenergy 2022, 157, 106331. [Google Scholar] [CrossRef]
- Lay, J.J.; Li, Y.Y.; Noike, T. The influence of pH and ammonia concentration on the methane production in high-solids digestion processes. Water Environ. Res. 1998, 70, 1075–1082. [Google Scholar] [CrossRef]
- Song, L.; Song, Y.; Li, D.; Liu, R.; Niu, Q. The auto fluorescence characteristics, specific activity, and microbial community structure in batch tests of mono-chicken manure digestion. Waste Manag. 2019, 83, 57–67. [Google Scholar] [CrossRef]
- Yenigün, O.; Demirel, B. Ammonia inhibition in anaerobic digestion: A review. Process Biochem. 2013, 48, 901–911. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Kong, X.; Li, L.; Yuan, Z.; Dong, R.; Sun, Y. Effects of ammonia on propionate degradation and microbial community in digesters using propionate as a sole carbon source. J. Chem. Technol. Biotechnol. 2017, 92, 2538–2545. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, P.; Zhang, Y.; Wang, X.; Peng, X.; Li, L. Consecutive batch anaerobic digestion under ammonia stress: Microbial community assembly and process performance. J. Environ. Chem. Eng. 2021, 9, 106061. [Google Scholar] [CrossRef]
- Conklin, A.; Stensel, H.D.; Ferguson, J. Growth kinetics and competition between methanosarcina and methanosaeta in mesophilic anaerobic digestion. Water Environ. Res. 2006, 78, 486–496. [Google Scholar] [CrossRef]
- Fryer, M.; O’Flaherty, E.; Gray, N.F. Evaluating the measurement of activated sludge foam potential. Water 2011, 3, 424–444. [Google Scholar] [CrossRef]
- Strom, N.; Ma, Y.; Bi, Z.; Andersen, D.; Trabue, S.; Chen, C.; Hu, B. Eubacterium coprostanoligenes and Methanoculleus identified as potential producers of metabolites that contribute to swine manure foaming. J. Appl. Microbiol. 2022, 132, 2906–2924. [Google Scholar] [CrossRef]
- Pagilla, K.R.; Craney, K.C.; Kido, W.H. Causes and effects of foaming in anaerobic sludge digesters. Wat. Sci. Technol. 1997, 36, 463–470. [Google Scholar] [CrossRef]
- He, Q.; Li, L.; Zhao, X.; Qu, L.; Wu, D.; Peng, X. Investigation of foaming causes in three mesophilic food waste digesters: Reactor performance and microbial analysis. Sci. Rep. 2017, 7, 13701. [Google Scholar] [CrossRef]
- Steinberg, L.M.; Regan, J.M. Response of lab-scale methanogenic reactors inoculated from different sources to organic loading rate shocks. Bioresour. Technol. 2011, 102, 8790–8798. [Google Scholar] [CrossRef]
- Fonoll, X.; Astals, S.; Dosta, J.; Mata-Alvarez, J. Anaerobic co-digestion of sewage sludge and fruit wastes: Evaluation of the transitory states when the co-substrate is changed. Chem. Eng. J. 2015, 262, 1268–1274. [Google Scholar] [CrossRef]
- Coggins, L.X.; Ghisalberti, M.; Ghadouani, A. Sludge accumulation and distribution impact the hydraulic performance in waste stabilisation ponds. Water Res. 2017, 110, 354–365. [Google Scholar] [CrossRef]
- Wang, L.K.; Hung, Y.-T.; Lo, H.H.; Yapijakis, C. Waste Treatment in the Food Processing Industry; Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- Liang, H.; Luo, X.; Ye, D.; Yan, J.; Luo, L. Seasonal succession of bacterial communities, metabolic potential and ecological functions in soy sauce wastewater treatment system. Int. Biodeterior. Biodegrad. 2020, 154, 105083. [Google Scholar] [CrossRef]
- Austermann-Haun, U.; Seyfried, C.F.; Rosenwinkel, K.-H. Full scale experiences with anaerobic pre-treatment of wastewater in the food and beverage industry in Germany. Wat. Sci. Technol. 1997, 36, 321–328. [Google Scholar] [CrossRef]
- Guo, J.; Peng, Y.; Ni, B.J.; Han, X.; Fan, L.; Yuan, Z. Dissecting microbial community structure and methane-producing pathways of a full-scale anaerobic reactor digesting activated sludge from wastewater treatment by metagenomic sequencing. Microb. Cell Fact. 2015, 14, 33. [Google Scholar] [CrossRef]
- Jiang, F.; Gao, H.; Qin, W.; Song, P.; Wang, H.; Zhang, J.; Liu, D.; Wang, D.; Zhang, T. Marked seasonal variation in structure and function of gut microbiota in forest and alpine musk deer. Front. Microbiol. 2021, 12, 699797. [Google Scholar] [CrossRef]
- Morotomi, M.; Nagai, F.; Watanabe, Y. Description of Christensenella minuta gen. nov., sp. nov., isolated from human faeces, which forms a distinct branch in the order Clostridiales, and proposal of Christensenellaceae fam. Nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 144–149. [Google Scholar] [CrossRef]
- Werner, J.J.; Knights, D.; Garcia, M.L.; Scalfone, N.B.; Smith, S.; Yarasheski, K.; Cummings, T.A.; Beers, A.R.; Knight, R.; Angenent, L.T. Bacterial community structures are unique and resilient in full-scale bioenergy systems. Proc. Natl. Acad. Sci. USA 2011, 108, 4158–4163. [Google Scholar] [CrossRef]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E. The Prokaryotes, 3rd ed.; Springer Science+Business Media: New York, NY, USA, 2006; Volume 4. [Google Scholar]
- Guo, F.; Zhang, T.; Li, B.; Wang, Z.; Ju, F.; Liang, Y.T. Mycobacterial species and their contribution to cholesterol degradation in wastewater treatment plants. Sci. Rep. 2019, 9, 836. [Google Scholar] [CrossRef]
- Bonk, F.; Popp, D.; Weinrich, S.; Strauber, H.; Kleinsteuber, S.; Harms, H.; Centler, F. Intermittent fasting for microbes: How discontinuous feeding increases functional stability in anaerobic digestion. Biotechnol. Biofuels 2018, 11, 274. [Google Scholar] [CrossRef]
- Kang, S.; Khan, S.; Webb, R.; Denman, S.; McSweeney, C. Characterization and survey in cattle of a rumen Pyrimadobacter sp. Which degrades the plant toxin fluoroacetate. FEMS Microbiol. Ecol. 2020, 96, fiaa077. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Gaio, J.; Lora, N.L.; Iltchenco, J.; Magrini, F.E.; Paesi, S. Seasonal characterization of the prokaryotic microbiota of full-scale anaerobic UASB reactors treating domestic sewage in southern Brazil. Bioprocess. Biosyst. Eng. 2023, 46, 69–87. [Google Scholar] [CrossRef]
- Zhang, L.; Loh, K.C.; Zhang, J.; Mao, L.; Tong, Y.W.; Wang, C.H.; Dai, Y. Three-stage anaerobic co-digestion of food waste and waste activated sludge: Identifying bacterial and methanogenic archaeal communities and their correlations with performance parameters. Bioresour. Technol. 2019, 285, 121333. [Google Scholar] [CrossRef]
- De Vrieze, J.; Hennebel, T.; Boon, N.; Verstraete, W. Methanosarcina: The rediscovered methanogen for heavy duty biomethanation. Bioresour. Technol. 2012, 112, 1–9. [Google Scholar] [CrossRef]
- Vanwonterghem, I.; Jensen, P.D.; Dennis, P.G.; Hugenholtz, P.; Rabaey, K.; Tyson, G.W. Deterministic processes guide long-term synchronised population dynamics in replicate anaerobic digesters. ISME J. 2014, 8, 2015–2028. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biosolids | Biochar | |
|---|---|---|
| Total solids (%) | 84% | 100% |
| Volatile solids (%) | 43% | 14% |
| pH | 6.7 | 9.6 |
| Total organic carbon (g/kg) | 230 | 120 |
| Total nitrogen (g/kg) | 41 | 24 |
| C:N | 5.6 | 5 |
| Total phosphorus (g/kg) | 20 | 12 |
| Total sulphur (g/kg) | 5 | 7 |
| Type | ISR | A:B | I + S:BC |
|---|---|---|---|
| Control | 1:0 | - | 1:0 |
| Sample A | 3.5:1 | 1:0 | 1:0 |
| Co-digestion | 3.5:1 | 1:1 | 1:0 |
| BC control | 1:0 | - | 2:1 |
| BC Sample A | 3.5:1 | 1:0 | 2:1 |
| BC co-digestion | 3.5:1 | 1:1 | 2:1 |
| Stage | Day | Reactor | HRT (d) | OLR (gVSsubstrate L−1 d−1) | Substrate A:Substrate B |
|---|---|---|---|---|---|
| I | 0–45 | 1 | 30 | 8.14 | 1:0 |
| 2 | 30 | 8.14 | 1:0 | ||
| 3 | 45 | 5.42 | 1:0 | ||
| 4 | 45 | 5.42 | 1:0 | ||
| II | 45–49 1 | 1 | 30 | 9.18 | 1:1 |
| 2 | 30 | 9.18 | 1:1 | ||
| 3 | 45 | 6.12 | 1:1 | ||
| 4 | 45 | 6.12 | 1:1 | ||
| III | 49–56 | 1 | 30 | 0.00 | - |
| 2 | 30 | 0.00 | - | ||
| 3 | 45 | 0.00 | - | ||
| 4 | 45 | 6.12 | 1:1 | ||
| IV | 56–74 | 1 | 30 | 9.18 | 1:1 |
| 2 | 30 | 9.18 | 1:1 | ||
| 3 | 45 | 6.12 | 1:1 | ||
| 4 | 45 | 6.12 | 1:1 | ||
| V | 74–83 | 1 | 30 | 0.00 | - |
| 2 | 30 | 0.00 | - | ||
| 3 | 45 | 0.00 | - | ||
| 4 | 45 | 0.00 | - | ||
| VI | 83–95 | 1 | 30 | 9.18 | 1:1 |
| 2 | 30 | 9.18 | 1:1 | ||
| 3 | 45 | 6.12 | 1:1 | ||
| 4 | 45 | 6.12 | 1:1 |
| HRT (d) | Stage | Feed Substrate | Gas Volume (mL L−1 d−1) | Gas Yield (mL gVSadded−1 d−1) | CH4 Volume (mL L−1 d−1) | CH4 Yield (mL gVSadded−1 d−1) |
|---|---|---|---|---|---|---|
| 30 | I | A | 719.8 ± 94.9 | 162.0 ± 21.4 | 243.3 ± 50.2 | 54.7 ± 11.3 |
| 30 | II | A + B | 451.2 ± 66.3 | 95.5 ± 14 | 107.0 ± 12.3 | 22.7 ± 2.6 |
| 30 | III | - * | 180.3 ± 82.7 | 38.2 ± 17.5 | 55.8 ± 25.7 | 11.8 ± 5.4 |
| 30 | IV | A + B | 329.5 ± 246.3 | 69.8 ± 52.2 | 86.7 ± 75.8 | 18.4 ± 16 |
| 30 | V | - | 198.9 ± 101.7 | 42.1 ± 21.5 | 52.5 ± 30.8 | 11.1 ± 6.5 |
| 30 | VI | A + B | 634.3 ± 258.7 | 134.3 ± 54.8 | 150.4 ± 95.5 | 31.9 ± 20.2 |
| 45 | I | A | 883.3 ± 145.5 | 302.9 ± 49.9 | 387.9 ± 70.3 | 133.0 ± 24.1 |
| 45 | II | A + B | 1047.2 ± 123.3 | 342.7 ± 40.4 | 427.9 ± 94.2 | 140.0 ± 30.8 |
| 45 | III | - * | ||||
| 45 | IV | A + B | 396.5 ± 283.1 | 129.8 ± 92.7 | 163.7 ± 134.8 | 53.6 ± 44.1 |
| 45 | V | - | 137.3 ± 64 | 44.9 ± 20.9 | 37.1 ± 19.5 | 12.1 ± 6.4 |
| 45 | VI | A + B | 483.8 ± 142.5 | 158.3 ± 46.6 | 124.1 ± 37.6 | 40.6 ± 12.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elliott, J.A.K.; Krohn, C.; Ball, A.S. Source-Separated Industrial Wastewater Is a Candidate for Biogas Production through Anaerobic Digestion. Fermentation 2024, 10, 165. https://doi.org/10.3390/fermentation10030165
Elliott JAK, Krohn C, Ball AS. Source-Separated Industrial Wastewater Is a Candidate for Biogas Production through Anaerobic Digestion. Fermentation. 2024; 10(3):165. https://doi.org/10.3390/fermentation10030165
Chicago/Turabian StyleElliott, Jake A. K., Christian Krohn, and Andrew S. Ball. 2024. "Source-Separated Industrial Wastewater Is a Candidate for Biogas Production through Anaerobic Digestion" Fermentation 10, no. 3: 165. https://doi.org/10.3390/fermentation10030165
APA StyleElliott, J. A. K., Krohn, C., & Ball, A. S. (2024). Source-Separated Industrial Wastewater Is a Candidate for Biogas Production through Anaerobic Digestion. Fermentation, 10(3), 165. https://doi.org/10.3390/fermentation10030165