Effect of Candida intermedia LAMAP1790 Antimicrobial Peptides against Wine-Spoilage Yeasts Brettanomyces bruxellensis and Pichia guilliermondii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Media
2.2. Obtained Supernatant with Antifungal Activity of C. intermedia and Characterization of the Protein Nature of This Activity
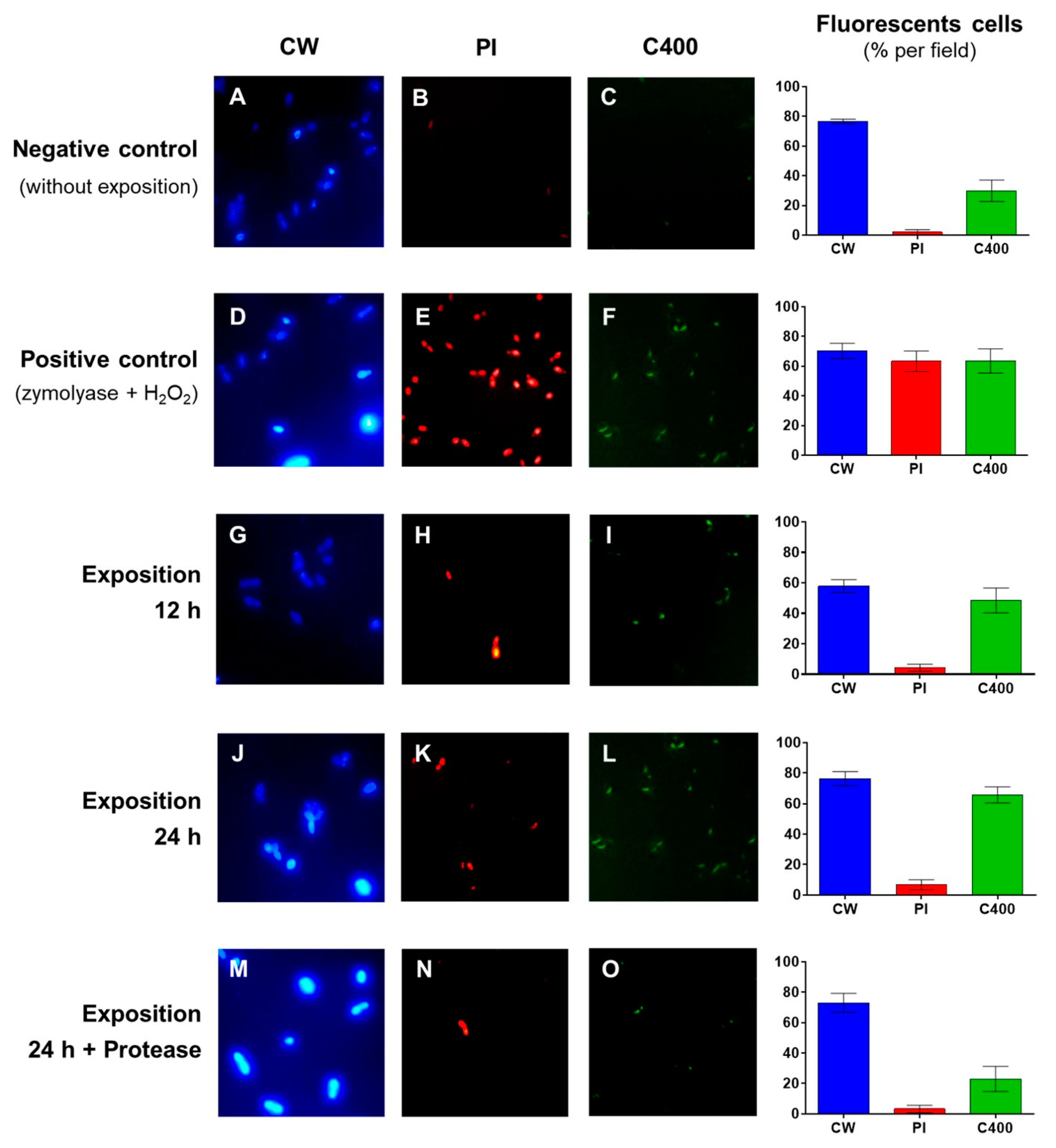
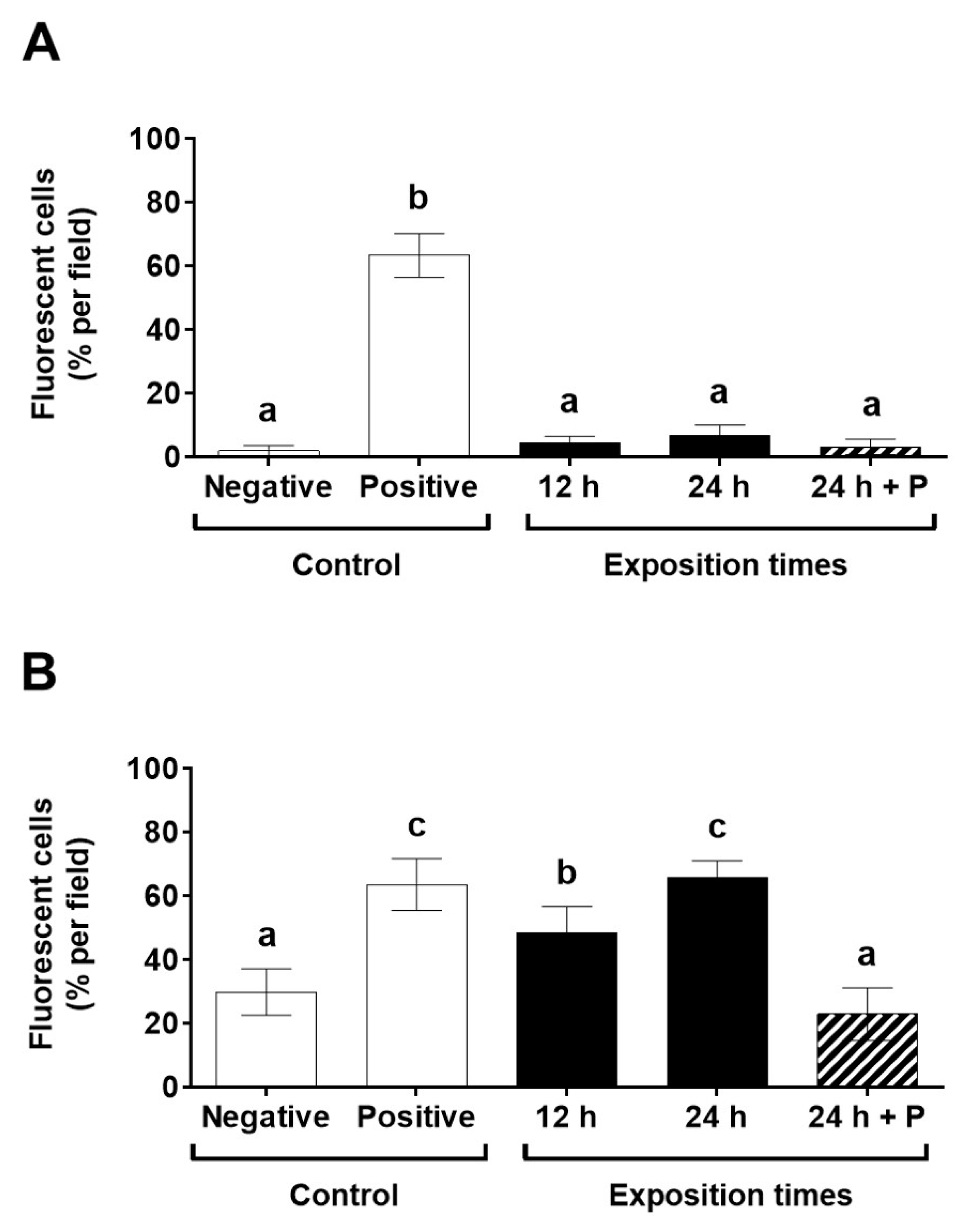
2.3. Determination of the Cellular Damage Produced on B. bruxellensis by Exposure to Antifungal Supernatant of C. intermedia
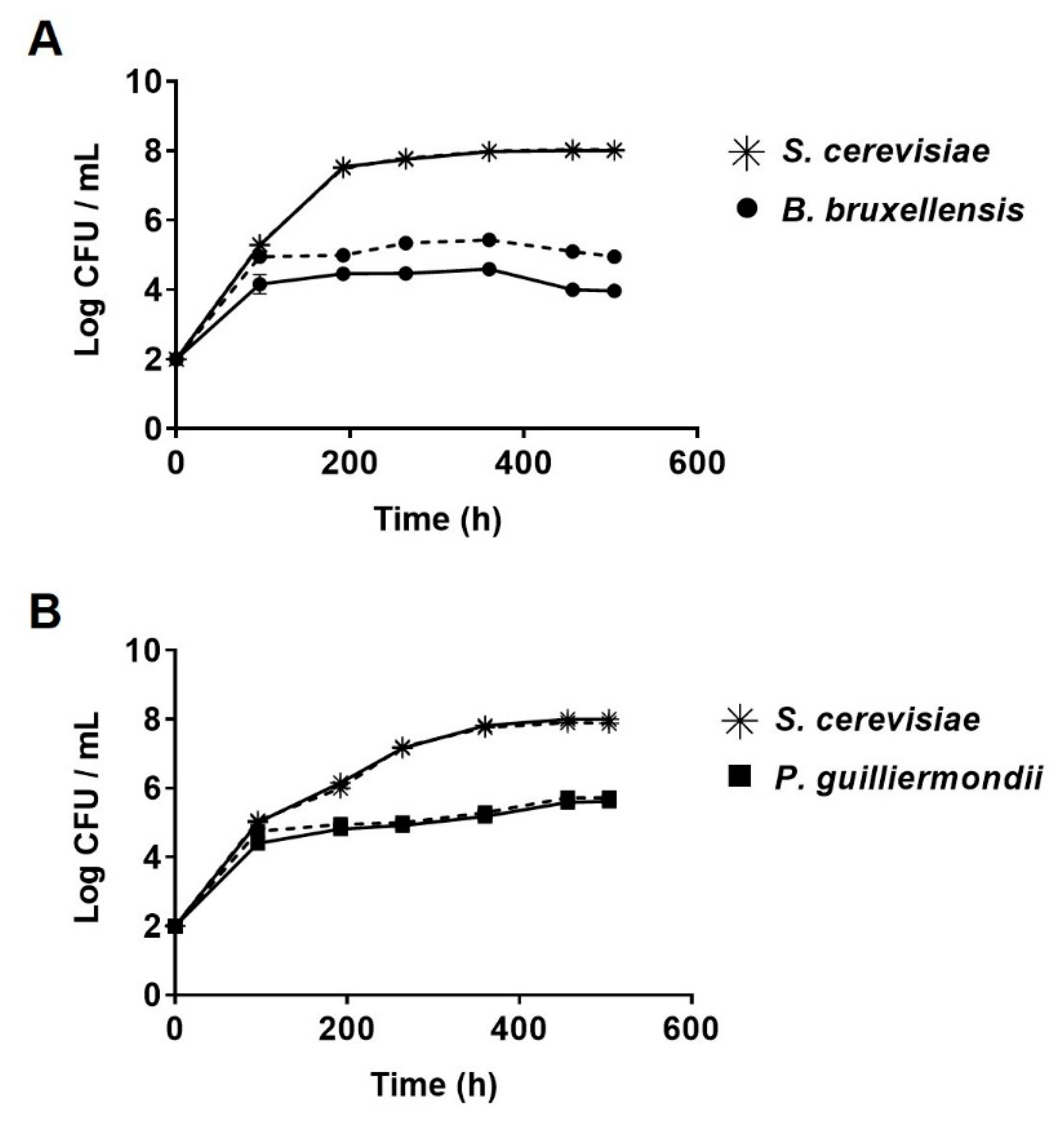
2.4. Screening the Antifungal Activity of C. intermedia LAMAP1790 on B. bruxellensis, P. guilliermondii, and S. cerevisiae
2.5. Antifungal Activity of Low Mass Peptide Fraction Obtained from C. intermedia Antifungal Supernatant against B. bruxellensis, S. cerevisiae and P. guilliermondii in Synthetic Must
2.6. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome: A dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2017, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Oelofse, A.; Pretorius, I.S.; du Toit, M. Significance of Brettanomyces and Dekkera during Winemaking: A Synoptic Review. S. Afr. J. Enol. Vitic. 2008, 29, 128–144. [Google Scholar] [CrossRef] [Green Version]
- Chatonnet, P.; Dubourdie, D.; Boidron, J.; Pons, M. The origin of ethylphenols in wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Godoy, L.; García, V.; Peña, R.; Martínez, C.; Ganga, M.A. Identification of the Dekkera bruxellensis phenolic acid decarboxylase (PAD) gene responsible for wine spoilage. Food Control 2014, 45, 81–86. [Google Scholar] [CrossRef]
- Dias, L.; Dias, S.; Sancho, T.; Stender, H.; Querol, A.; Malfeito-Ferreira, M.; Loureiro, V. Identification of yeasts isolated from wine-related environments and capable of producing 4-ethylphenol. Food Microbiol. 2003, 20, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Devalia, J.L.; Rusznak, C.; Herdman, M.J.; Trigg, C.J.; Davies, R.J.; Tarraf, H. Effect of nitrogen dioxide and sulphur dioxide on airway response of mild asthmatic patients to allergen inhalation. Lancet 1994, 344, 1668–1671. [Google Scholar] [CrossRef]
- Enrique, M.; Marcos, J.F.; Yuste, M.; Martínez, M.; Vallés, S.; Manzanares, P. Antimicrobial action of synthetic peptides towards wine spoilage yeasts. Int. J. Food Microbiol. 2007, 118, 318–325. [Google Scholar] [CrossRef]
- Enrique, M.; Marcos, J.F.; Yuste, M.; Martínez, M.; Vallés, S.; Manzanares, P. Inhibition of the wine spoilage yeast Dekkera bruxellensis by bovine lactoferrin-derived peptides. Int. J. Food Microbiol. 2008, 127, 229–234. [Google Scholar] [CrossRef]
- Albergaria, H.; Francisco, D.; Gori, K.; Arneborg, N.; Gírio, F. Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl. Microbiol. Biotechnol. 2010, 86, 965–972. [Google Scholar] [CrossRef]
- Branco, P.; Viana, T.; Albergaria, H.; Arneborg, N. Antimicrobial peptides (AMPs) produced by Saccharomyces cerevisiae induce alterations in the intracellular pH, membrane permeability and culturability of Hanseniaspora guilliermondii cells. Int. J. Food Microbiol. 2015, 205, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Branco, P.; Francisco, D.; Monteiro, M.; Almeida, M.G.; Caldeira, J.; Arneborg, N.; Prista, C.; Albergaria, H. Antimicrobial properties and death-inducing mechanisms of saccharomycin, a biocide secreted by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 159–171. [Google Scholar] [CrossRef]
- Muñoz, A.; Harries, E.; Contreras-Valenzuela, A.; Carmona, L.; Read, N.D.; Marcos, J.F. Two Functional Motifs Define the Interaction, Internalization and Toxicity of the Cell-Penetrating Antifungal Peptide PAF26 on Fungal Cells. PLoS ONE 2013, 8, e54813. [Google Scholar] [CrossRef]
- Muñoz, A.; Gandía, M.; Harries, E.; Carmona, L.; Read, N.D.; Marcos, J.F. Understanding the mechanism of action of cell-penetrating antifungal peptides using the rationally designed hexapeptide PAF26 as a model. Fungal Biol. Rev. 2013, 26, 146–155. [Google Scholar] [CrossRef]
- Zhang, L.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Peña, R.; Ganga, M.A. Novel antimicrobial peptides produced by Candida intermedia LAMAP1790 active against the wine-spoilage yeast Brettanomyces bruxellensis. Antonie van Leeuwenhoek 2018, 9. [Google Scholar] [CrossRef]
- Peña, R.; Chávez, R.; Rodríguez, A.; Ganga, M.A. A Control Alternative for the Hidden Enemy in the Wine Cellar. Fermentation 2019, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Ganga, M.A.; Martínez, C. Effect of wine yeast monoculture practice on the biodiversity of non-Saccharomyces yeasts. J. Appl. Microbiol. 2004, 96, 76–83. [Google Scholar] [CrossRef]
- Valdes, J.; Tapia, P.; Cepeda, V.; Varela, J.; Godoy, L.; Cubillos, F.A.; Silva, E.; Martinez, C.; Ganga, M.A. Draft genome sequence and transcriptome analysis of the wine spoilage yeast Dekkera bruxellensis LAMAP2480 provides insights into genetic diversity, metabolism and survival. FEMS Microbiol. Lett. 2014, 361, 104–106. [Google Scholar] [CrossRef] [Green Version]
- Sangorrín, M.P.; García, V.; Lopes, C.A.; Sáez, J.S.; Martínez, C.; Ganga, M.A. Molecular and physiological comparison of spoilage wine yeasts. J. Appl. Microbiol. 2013, 114, 1066–1074. [Google Scholar] [CrossRef] [Green Version]
- Roostita, L.B.; Fleet, G.H.; Wendry, S.P.; Apon, Z.M.; Gemilang, L.U. Determination of yeasts antimicrobial activity in milk and meat products. Adv. J. Food Sci. Technol. 2011, 3, 442–445. [Google Scholar]
- Kaiserer, L.; Oberparleiter, C.; Weiler-Görz, R.; Burgstaller, W.; Leiter, E.; Marx, F. Characterization of the Penicillium chrysogenum antifungal protein PAF. Arch. Microbiol. 2003, 180, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Villalba, M.L.; Susana Sáez, J.; del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Chiva, R.; Sancho, M.; Beltran, G.; Arroyo-López, F.N.; Guillamon, J.M. Nitrogen requirements of commercial wine yeast strains during fermentation of a synthetic grape must. Food Microbiol. 2012, 31, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Coronado, P.; Aguilera, S.; Carmona, L.; Godoy, L.; Martínez, C.; Ganga, M.A. Comparison of the behaviour of Brettanomyces bruxellensis strain LAMAP L2480 growing in authentic and synthetic wines. Antonie van Leeuwenhoek 2015, 107, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Sáez, J.S.; Lopes, C.A.; Kirs, V.C.; Sangorrín, M.P. Enhanced volatile phenols in wine fermented with Saccharomyces cerevisiae and spoiled with Pichia guilliermondii and Dekkera bruxellensis. Lett. Appl. Microbiol. 2010, 51, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Harrington, B.J.; Hageage, G.J. your lab focus: Calcofluor White: A Review of its Uses and Applications in Clinical Mycology and Parasitology. Lab. Med. 2003, 34, 361–367. [Google Scholar] [CrossRef]
- Suzuki, S.W.; Onodera, J.; Ohsumi, Y. Starvation induced cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction. PLoS ONE 2011, 6, e17412. [Google Scholar] [CrossRef] [Green Version]
- Reggiori, F.; Klionsky, D.J. Autophagic processes in yeast: Mechanism, machinery and regulation. Genetics 2013, 194, 341–361. [Google Scholar] [CrossRef] [Green Version]
- López-García, B.; Gandía, M.; Muñoz, A.; Carmona, L.; Marcos, J.F. A genomic approach highlights common and diverse effects and determinants of susceptibility on the yeast Saccharomyces cerevisiae exposed to distinct antimicrobial peptides. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.A.; Sangorrín, M.P. Optimization of killer assays for yeast selection protocols. Rev. Argent. Microbiol. 2010, 42, 298–306. [Google Scholar] [CrossRef]
- Stead, D. The effect of hydroxycinnamic acids and potassium sorbate on the growth of 11 strains of spoilage yeasts. J. Appl. Bacteriol. 1995, 78, 82–87. [Google Scholar] [CrossRef]
- Harris, V.; Jiranek, V.; Ford, C.M.; Grbin, P.R. Inhibitory effect of hydroxycinnamic acids on Dekkera spp. Appl. Microbiol. Biotechnol. 2010, 86, 721–729. [Google Scholar] [CrossRef]
- Lentz, M.; Harris, C. Analysis of Growth Inhibition and Metabolism of Hydroxycinnamic Acids by Brewing and Spoilage Strains of Brettanomyces Yeast. Foods 2015, 4, 581–593. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| C. intermedia LAMAP1790 | ||
|---|---|---|
| Inhibition Halo (mm) | ||
| S. cerevisiae | EC1118 | † ND a |
| B. bruxellensis | LAMAP2480 | 19.00 ± 0.62 c |
| P. guilliermondii | LAMAP3202 | 15.33 ± 0.82 b |
| LAMAP3203 | 16.17 ± 0.75 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña, R.; Vílches, J.; G.-Poblete, C.; Ganga, M.A. Effect of Candida intermedia LAMAP1790 Antimicrobial Peptides against Wine-Spoilage Yeasts Brettanomyces bruxellensis and Pichia guilliermondii. Fermentation 2020, 6, 65. https://doi.org/10.3390/fermentation6030065
Peña R, Vílches J, G.-Poblete C, Ganga MA. Effect of Candida intermedia LAMAP1790 Antimicrobial Peptides against Wine-Spoilage Yeasts Brettanomyces bruxellensis and Pichia guilliermondii. Fermentation. 2020; 6(3):65. https://doi.org/10.3390/fermentation6030065
Chicago/Turabian StylePeña, Rubén, Jeniffer Vílches, Camila G.-Poblete, and María Angélica Ganga. 2020. "Effect of Candida intermedia LAMAP1790 Antimicrobial Peptides against Wine-Spoilage Yeasts Brettanomyces bruxellensis and Pichia guilliermondii" Fermentation 6, no. 3: 65. https://doi.org/10.3390/fermentation6030065
APA StylePeña, R., Vílches, J., G.-Poblete, C., & Ganga, M. A. (2020). Effect of Candida intermedia LAMAP1790 Antimicrobial Peptides against Wine-Spoilage Yeasts Brettanomyces bruxellensis and Pichia guilliermondii. Fermentation, 6(3), 65. https://doi.org/10.3390/fermentation6030065