Abatement of Inhibitors in Recycled Process Water from Biomass Fermentations Relieves Inhibition of a Saccharomyces cerevisiae Pentose Phosphate Pathway Mutant †


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, Culture Conditions
2.2. Yeast Growth Assays
2.3. Construction of S. cerevisiae Mutants
2.4. Preparation and Analysis of Hydrolysate
2.5. Inhibitor Abatement
2.6. Ethanol Fermentations and Recycle of Hydrolysate
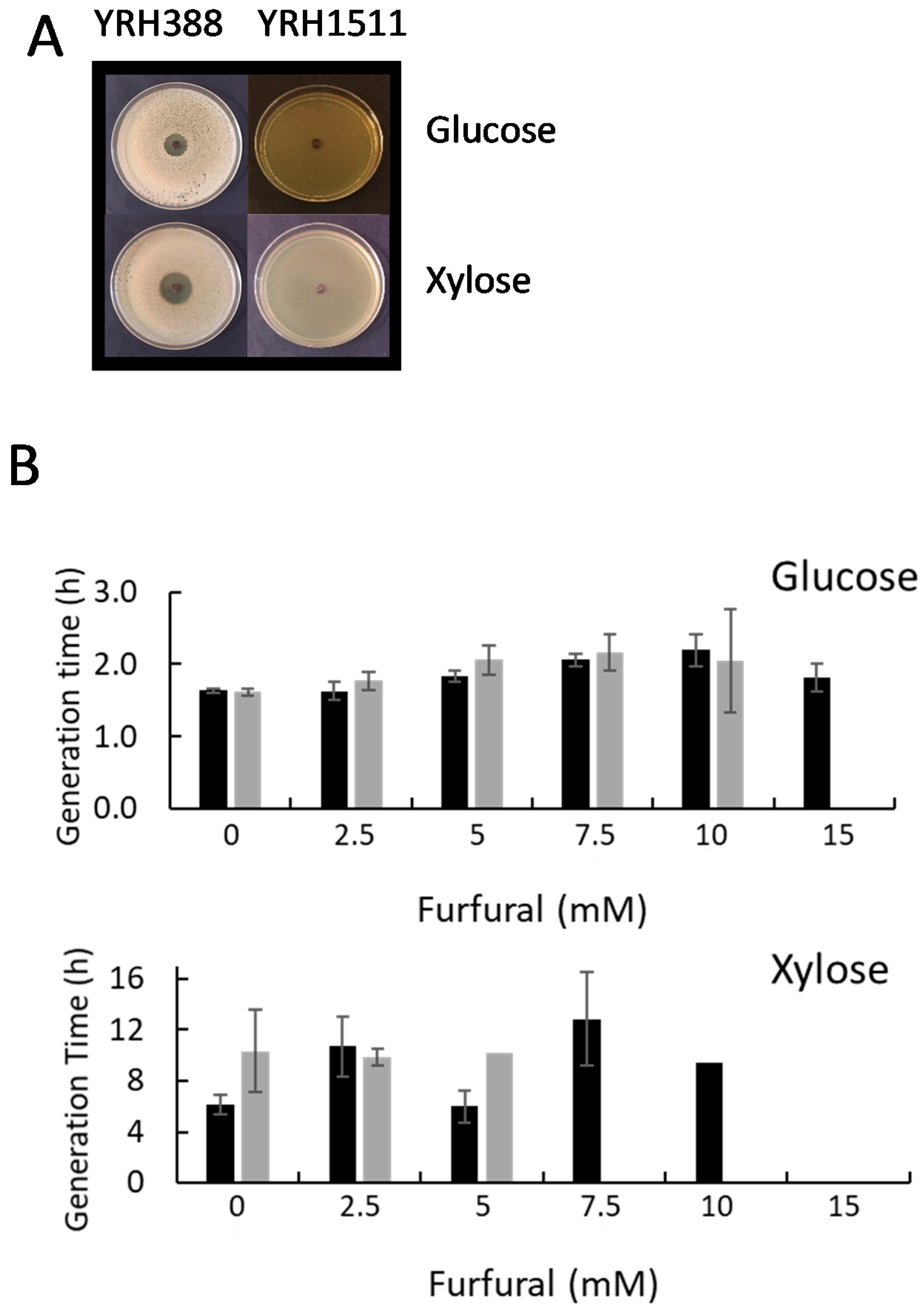
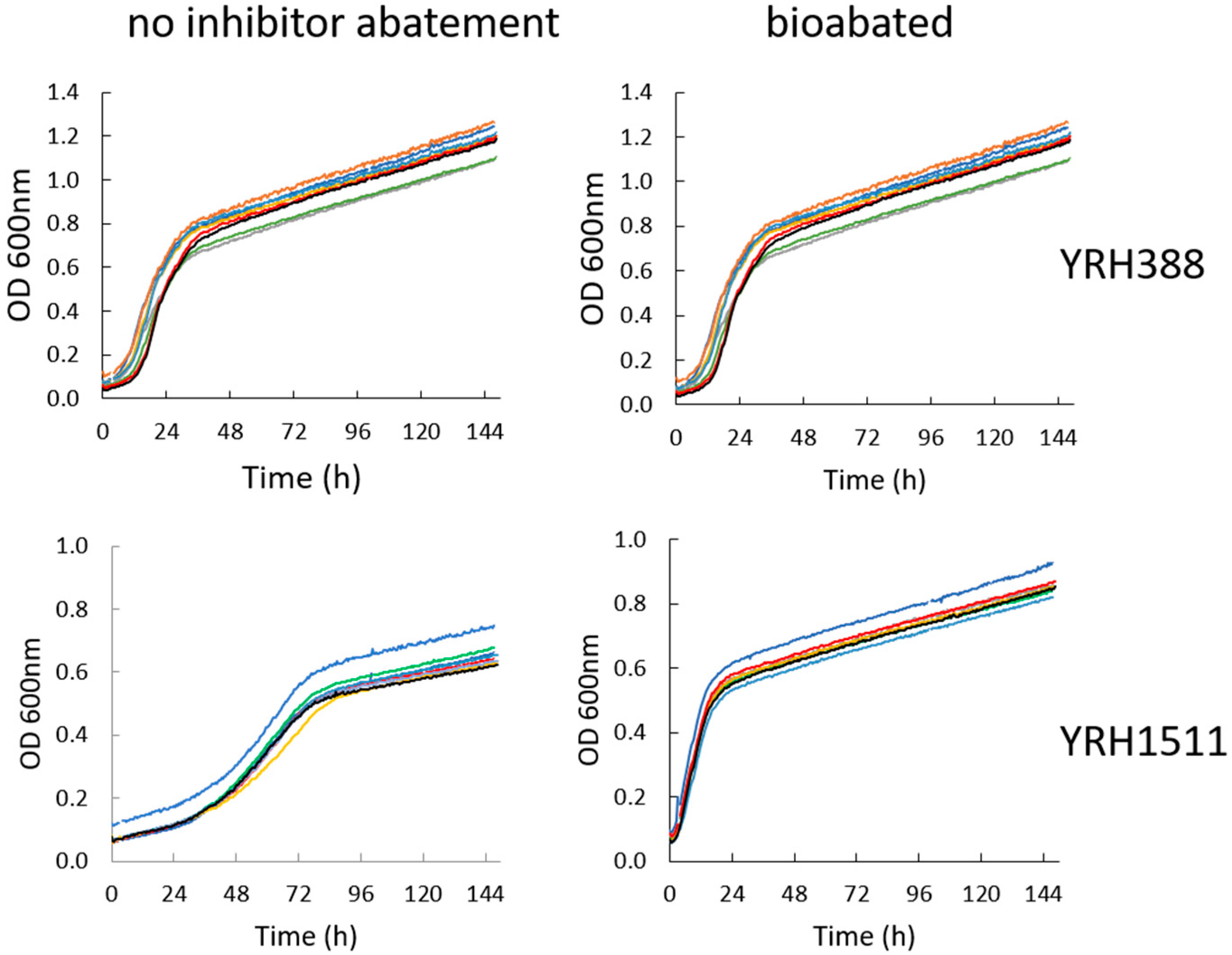
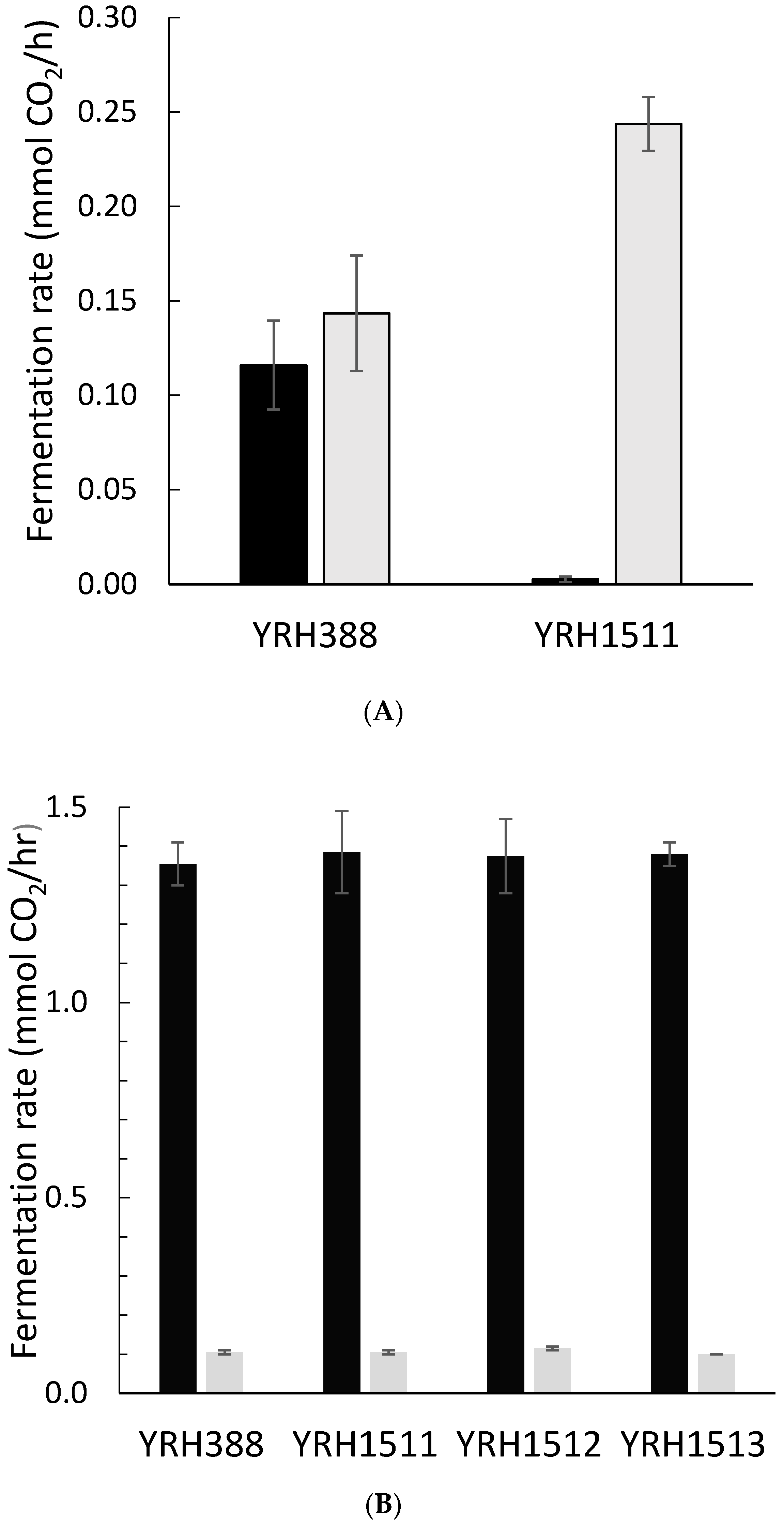
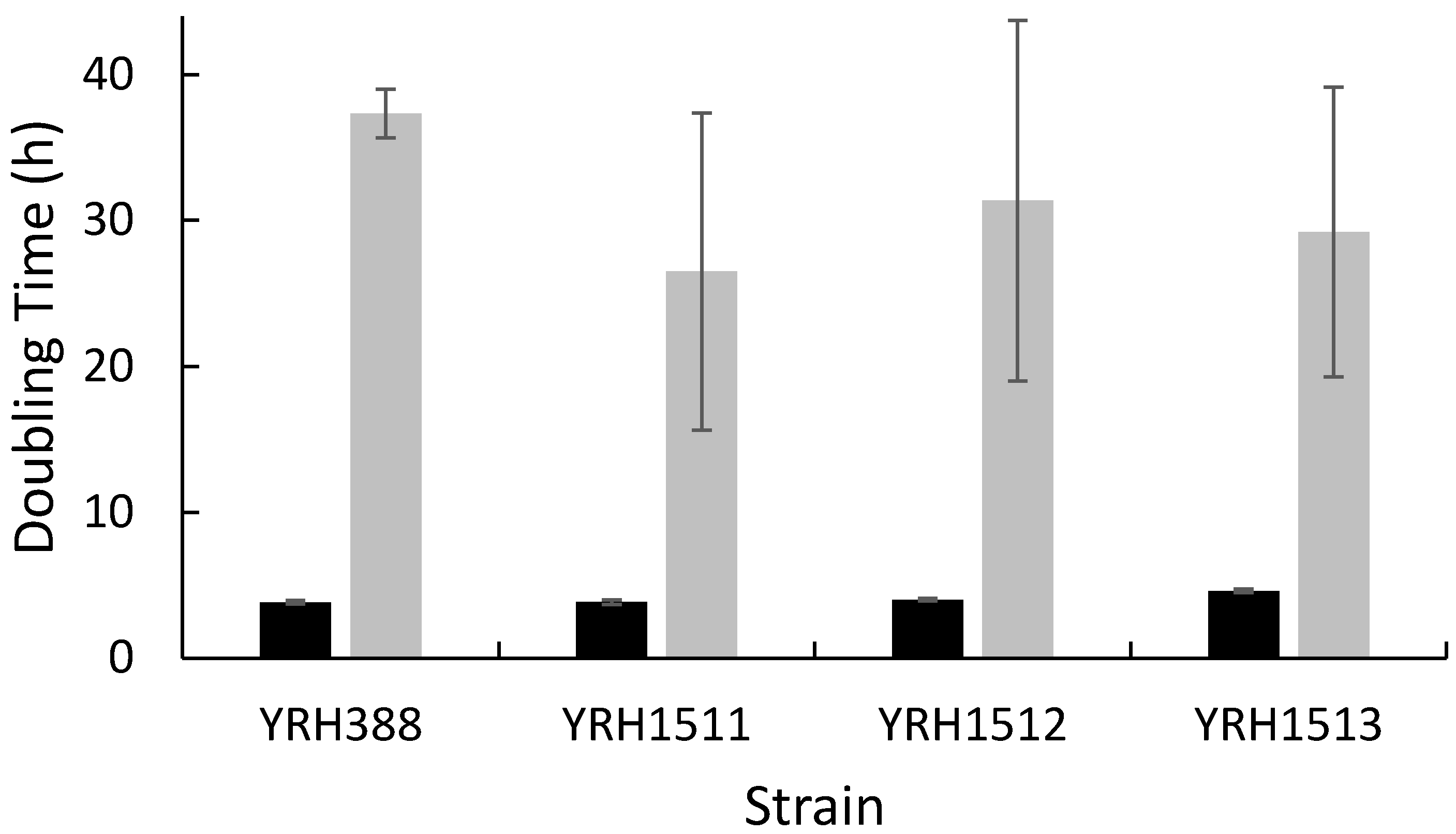
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Esteghlalian, A.; Hashimoto, A.G.; Fenske, J.J.; Penner, M.J. Modeling and optimization of the dilute-sulfuric-acid pretreatment of corn stover, poplar and switch-grass. Bioresour. Technol. 1997, 59, 129–136. [Google Scholar] [CrossRef]
- Modig, T.; Liden, G.; Taherzadeh, M.J. Inhibition effects of furfural on alcohol dehydrogenase, aldehyde dehydrogenase and pyruvate dehydrogenase. Biochem. J. 2002, 363, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.L.; Santo, R.; Reis, A.; Passarinho, P.C. Effect of furfural on Saccharomyces carlsbergensis growth, physiology and ethanol production. Appl. Biochem. Biotechnol. 2017, 182, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Lee, Y.S.; Li, J.; Li, C. Resistance mechanisms and reprogramming of microorganisms for efficient biorefinery under multiple environmental stresses. Synth. Syst. Biotechnol. 2019, 4, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Dessie, W.; Xin, F.; Zhang, W.; Zhou, J.; Wu, H.; Ma, J.; Jiang, M. Inhibitory effects of lignocellulose pretreatment degradation products (hydroxymethylfurfural and furfural) on succinic acid producing Actinobacillus succinogenes. Biochem. Eng. J. 2019, 107263. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. I: Inhibition and detoxification. Bioresour. Technol. 2000, 74, 17–24. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Palmqvist, E.; Grage, H.; Meinander, N.Q.; Hahn-Hägerdal, B. Main and interaction effects of acetic acid, furfural, and p-hydroxybenzoic acid on growth and ethanol productivity of yeasts. Biotechnol. Bioeng. 1999, 63, 46–55. [Google Scholar] [CrossRef]
- Zaldivar, J.; Martinez, A.; Ingram, L.O. Effect of selected aldehydes on the growth and fermentation of ethanologenic Escherichia coli. Biotechnol. Bioeng. 1999, 65, 24–33. [Google Scholar] [CrossRef]
- Martín, M.; Ahmetović, E.; Grossmann, I.E. Optimization of water consumption in second generation bioethanol plants. Ind. Eng. Chem. Res. 2010, 50, 3705–3721. [Google Scholar] [CrossRef]
- Mika, L.T.; Cséfalvay, E.; Németh, A. Catalytic conversion of carbohydrates to initial platform chemicals: Chemistry and sustainability. Chem Rev. 2017, 118, 505–613. [Google Scholar] [CrossRef]
- Nizami, A.-S.; Mohanakrishna, G.; Mishra, U.; Pant, D. Trends & sustainability criteria for liquid biofuels. In Biofuels: Production and Future Perspectives; Singh, R.S., Pandey, A., Gnansounou, E., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2016; pp. 59–98. [Google Scholar]
- López, M.J.; Nichols, N.N.; Dien, B.S.; Moreno, J.; Bothast, R.J. Isolation of microorganisms for biological detoxification of lignocellulosic hydrolysates. Appl. Microbiol. Biotechnol. 2004, 64, 125–131. [Google Scholar] [CrossRef]
- Nichols, N.N.; Sharma, L.N.; Mowery, R.A.; Chambliss, C.K.; van Walsum, G.P.; Dien, B.S.; Iten, L.B. Fungal metabolism of fermentation inhibitors present in corn stover dilute acid hydrolysate. Enzym. Microb. Technol. 2008, 42, 624–630. [Google Scholar] [CrossRef]
- Balderas-Hernández, V.E.; Correia, K.; Mahadevan, R. Inactivation of the transcription factor mig1 (YGL035C) in Saccharomyces cerevisiae improves tolerance towards weak acids: Acetic, formic and levulinic acid. J. Ind. Microbiol. Biotechnol. 2018, 4, 735–751. [Google Scholar] [CrossRef]
- Casey, E.; Sedlak, M.; Ho, N.W.; Mosier, N.S. Effect of acetic acid and pH on the cofermentation of glucose and xylose to ethanol by a genetically engineered strain of Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 385–393. [Google Scholar] [CrossRef]
- Hector, R.E.; Dien, B.S.; Cotta, M.A.; Qureshi, N. Engineering industrial Saccharomyces cerevisiae strains for xylose fermentation and comparison for switchgrass conversion. J. Ind. Microbiol. Biotechnol. 2011, 38, 1193–1202. [Google Scholar] [CrossRef]
- Guo, Z.-P.; Olsson, L. Physiological responses to acid stress by Saccharomyces cerevisiae when applying high initial cell density. FEMS Yeast Res. 2016, 16, fow072. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.B. Another explanation for the toxicity of fermentation acids at low pH: Anion accumulation versus uncoupling. J. Appl. Bacteriol. 1992, 73, 363–370. [Google Scholar] [CrossRef]
- Nichols, N.N.; Mertens, J.A.; Hector, R.E.; Dien, B.S.; Frazer, S.E. Recycle of fermentation process water through mitigation of inhibitors in dilute-acid corn stover hydrolysate. Bioresour. Technol. Rep. 2020, 9, 100349. [Google Scholar] [CrossRef]
- Aden, A. Water usage for current and future ethanol production. Southwest Hydrol. 2007, 6, 22–23. [Google Scholar]
- Bothast, R.; Schlicher, M. Biotechnological process for conversion of corn into ethanol. Appl. Microbiol. Biotechnol. 2005, 67, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Dunham, M.J.; Gartenberg, M.R.; Brown, G.W. Methods in Yeast Genetics and Genomics: A Cold Spring Harbor Laboratory Course Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015; ISBN -13: 978-1621821342. [Google Scholar]
- Hector, R.E.; Bowman, M.J.; Skory, C.D.; Cotta, M.A. The Saccharomyces cerevisiae YMR315W gene encodes an NADP(H)-specific oxidoreductase regulated by the transcription factor Stb5p in response to NADPH limitation. New Biotechnol. 2009, 26, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Lõoke, M.; Kristjuhan, K.; Kristjuhan, A. Extraction of genomic DNA from yeasts for PCR-based applications. Biotechniques 2011, 50, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Nichols, N.N.; Hector, R.E.; Frazer, S.E. Factors affecting production of xylitol by the furfural-metabolizing fungus Coniochaeta ligniaria. Curr. Trends Microbiol. 2018, 12, 109–119. [Google Scholar]
- Somers, T.C.; Ziemelis, G. Spectral evaluation of total phenolic components in Vitas vinifera: Grapes and wines. J. Sci. Food Agric. 1985, 36, 1275–1284. [Google Scholar] [CrossRef]
- Nogae, I.; Johnston, M. Isolation and characterization of the ZWF1 gene of Saccharomyces cerevisiae, encoding glucose-6-phosphate dehydrogenase. Gene 1990, 96, 161–169. [Google Scholar] [CrossRef]
- Mulet, J.M.; Leube, M.P.; Kron, S.J.; Rios, G.; Fink, G.R.; Serrano, R. A novel mechanism of ion homeostasis and salt tolerance in yeast: The Hal4 and Hal5 protein kinases modulate the Trk1-Trk2 potassium transporter. Mol. Cell. Biol. 1999, 19, 3328–3337. [Google Scholar] [CrossRef] [Green Version]
- Albertyn, J.; Hohmann, S.; Thevelein, J.M.; Prior, B.A. GPD1, which encodes glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic stress in Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity glycerol response pathway. Mol. Cell. Biol. 1994, 14, 4135–4144. [Google Scholar] [CrossRef] [Green Version]
- Gorsich, S.W.; Dien, B.S.; Nichols, N.N.; Slininger, P.J.; Liu, Z.; Skory, C.D. Tolerance to furfural-induced stress is associated with pentose phosphate pathway genes ZWF1, GND1, RPEL, and TKL1 in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2005, 71, 339–349. [Google Scholar] [CrossRef]
- Saha, B.C.; Nichols, N.N.; Cotta, M.A. Ethanol production from wheat straw by recombinant Escherichia coli strain FBR5 at high solid loading. Bioresour. Technol. 2011, 102, 10892–10897. [Google Scholar] [CrossRef]
- Nichols, N.N.; Dien, B.S.; Cotta, M.A. Fermentation of bioenergy crops into ethanol using biological abatement for removal of inhibitors. Bioresour. Technol. 2010, 101, 7545–7550. [Google Scholar] [CrossRef] [PubMed]
- Jeppsson, M.; Träff, K.; Johansson, B.; Hahn-Hägerdal, B.; Gorwa-Grauslund, M.F. Effect of enhanced xylose reductase activity on xylose consumption and product distribution in xylose-fermenting recombinant Saccharomyces cerevisiae. FEMS Yeast Res. 2003, 3, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Hector, R.E.; Mertens, J.A.; Bowman, M.J.; Nichols, N.N.; Cotta, M.A.; Hughes, S.R. Saccharomyces cerevisiae engineered for xylose metabolism requires gluconeogenesis and the oxidative branch of the pentose phosphate pathway for aerobic xylose assimilation. Yeast 2011, 28, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, E.; Almeida, J.S.; Hahn-Hägerdal, B. Influence of furfural on anaerobic glycolytic kinetics of Saccharomyces cerevisiae in batch culture. Biotechnol. Bioeng. 1999, 62, 447–454. [Google Scholar] [CrossRef]
- Suko, A.V.; Bura, R. Enhanced xylitol and ethanol yields by fermentation inhibitors in steam-pretreated lignocellulosic biomass. Ind. Biotechnol. 2016, 12, 187–194. [Google Scholar] [CrossRef]
- Walker, G.M.; Basso, T.O. Mitigating stress in industrial yeasts. Fungal Biol. 2020, 124, 387–397. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Source or Reference |
|---|---|---|
| S. cerevisiae CEN.PK2-1C | MATaura3-52 trp1-289 leu2-3,112 his3Δ1 MAL2-8CSUC2 | EUROSCARF |
| S. cerevisiae YRH388 | CEN.PK2-1C hoΔ::(KanMX4; PPGK1-XYL1-TPGK1; PADH1-XYL2-TADH1; PHXT7-XKS1-THXT7) | [17] |
| S. cerevisiae YRH512 | CEN.PK2-1C zwf1Δ::LEU2 | This work |
| S. cerevisiae YRH1511 | YRH388 zwf1Δ::LEU2 | This work |
| S. cerevisiae YRH1512 | YRH388 hal4Δ::LEU2 | This work |
| S. cerevisiae YRH1513 | YRH388 gpd1Δ::LEU2 | This work |
| C. ligniaria NRRL 30616 | Bioabatement strain | [13] |
| Oligonucleotide | Sequence | |
| 229 | GGCAGTGTCTAAGACTTTACAGGGA | |
| 675 | gtacggaAAACTCGCCAAGGCTATCC | |
| 676 | CCAAAATGTCACTGACCGCGGCTAAA | |
| 677 | AAAAAAAAGTAATCAGATTTTATTTTATTTCGACATTACCCCTCAAATATAGATTGTACTGAGAGTGCAC | |
| 678 | AATACCGAATTGTTCCAAAATATCGTGATATTTGAAAGTGAAGTACTCGTCTGTGCGGTATTTCACACCG | |
| 679 | TATATTGTACACCCCCCCCCTCCACAAACACAAATATTGATAATATAAAGAGATTGTACTGAGAGTGCAC | |
| 680 | CCTCGAAAAAAGTGGGGGAAAGTATGATATGTTATCTTTCTCCAATAAATCTGTGCGGTATTTCACACCG | |
| 681 | ATGTGGGATTTTTGGCTCAAGGTGT | |
| 682 | TTTCCTGTCGGTTTCG | |
| 683 | TGCCGGATAACAAAGC | |
| 684 | CCGCACAACAAGTATCAGAATGGG | |
| 685 | AAATGCGGAAGAGGTGTACAGC |
| Hydrolysate 1 | HMF (mM) | Furfural (mM) | Phenolics (AU) | Glucose (% w/v) | Xylose (% w/v) | Arabinose (% w/v) | Galactose (% w/v) |
|---|---|---|---|---|---|---|---|
| No abatement | 0.60 ± 0.07 | 12.72 ± 0.84 | 167 ± 28 | 0.32 ± 0.00 | 1.82 ± 0.02 | 0.29 ± 0.02 | 0.14 ± 0.00 |
| Bioabated | 0.35 ± 0.13 | 2.86 ± 2.38 | 82 ± 73 | 0.24 ± 0.06 | 1.78 ± 0.02 | 0.28 ± 0.01 | 0.14 ± 0.00 |
| Recycled | 0.00 | 0.00 | 88 ± 9 | 0.00 | 0.38 | 0.15 | 0.04 |
| S. cerevisiae | Relevant | Phase of Growth | Doubling Time (h) | Maximum OD | ||
|---|---|---|---|---|---|---|
| Strain | Mutation | Unabated CSH | Bioabated CSH | Unabated CSH | Bioabated CSH | |
| YRH388 | --- | Early | 4.5 ± 0.4 | 3.7 ± 0.4 | ||
| YRH1511 | ZWF1 | Early | 19.5 ± 1.7 | 3.4 ± 0.2 | ||
| YRH1512 | HAL4 | Early | 4.8 ± 0.4 | 3.4 ± 0.7 | ||
| YRH1513 | GPD1 | Early | 5.7 ± 0.2 | 4.6 ± 0.4 | ||
| YRH388 | --- | Late | 90.1 ± 8.9 | 98.0 ± 6.7 | 1.19 ± 0.06 | 1.15 ± 0.06 |
| YRH1511 | ZWF1 | Late | 138.8 ± 13.6 | 123.9 ± 19.5 | 0.66 ± 0.04 | 0.76 ± 0.29 |
| YRH1512 | HAL4 | Late | 108.7 ± 13.3 | 91.5 ± 9.1 | 1.14 ± 0.04 | 1.14 ± 0.05 |
| YRH1513 | GPD1 | Late | 128.8 ± 16.0 | 49.6 ± 1.9 | 1.12 ± 0.05 | 1.56 ± 0.03 |
| Glucose Consumed 1 | Xylose Consumed | Residual Glucose | Residual Xylose | Ethanol | Glycerol | Xylitol | Ethanol Yield 2 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | ||||||||||||
| Unabated | YRH388 | 0.240 ± 0.079 | 0.192 ± 0.011 | 0.010 ± 0.003 | 1.548 ± 0.006 | 0.172 ± 0.011 | 0.010 ± 0.008 | 0.040 ± 0.023 | 0.398 | |||
| YRH1511 | 0.066 ± 0.059 | 0.020 ± 0.006 | 0.224 ± 0.034 | 1.726 ± 0.008 | 0.030 ± 0.031 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.349 | ||||
| Bioabated | YRH388 | 0.220 ± 0.025 | 0.317 ± 0.016 | 0.011 ± 0.001 | 1.415 ± 0.024 | 0.159 ± 0.015 | 0.066 ± 0.005 | 0.068 ± 0.001 | 0.296 | |||
| YRH1511 | 0.224 ± 0.023 | 0.160 ± 0.035 | 0.010 ± 0.002 | 1.568 ± 0.042 | 0.171 ± 0.017 | 0.008 ± 0.001 | 0.007 ± 0.010 | 0.445 | ||||
| B | ||||||||||||
| YRH388 | 0.956 ± 0.057 | 0.832 ± 0.059 | 0.000 ± 0.000 | 0.150 ± 0.014 | 0.544 ± 0.034 | 0.062 ± 0.003 | 0.350 ± 0.020 | 0.304 | ||||
| YRH1511 | 0.970 ± 0.048 | 0.670 ± 0.096 | 0.000 ± 0.000 | 0.326 ± 0.059 | 0.564 ± 0.023 | 0.052 ± 0.011 | 0.192 ± 0.028 | 0.344 | ||||
| YRH1512 | 0.942 ± 0.054 | 0.792 ± 0.071 | 0.000 ± 0.000 | 0.206 ± 0.020 | 0.562 ± 0.025 | 0.066 ± 0.008 | 0.302 ± 0.037 | 0.324 | ||||
| YRH1513 | 0.976 ± 0.011 | 0.880 ± 0.034 | 0.000 ± 0.000 | 0.164 ± 0.006 | 0.556 ± 0.023 | 0.064 ± 0.006 | 0.388 ± 0.000 | 0.300 | ||||
| Doubling Time (h) | Maximum OD | ||||
|---|---|---|---|---|---|
| S. cerevisiae Strain | Phase of Growth | Bioabated + Glucose 1 | Bioabated + Xylose 2 | Bioabated + Glucose | Bioabated + Xylose |
| YRH388 | Early | 1.1 ± 0.2 | 1.4 ± 0.1 | ||
| YRH1511 | Early | 1.6 ± 0.83 | 4.1 ± 3.9 | ||
| YRH388 | Late | 78.4 ± 37.9 | 62.8 ± 10.1 | 0.50 ± 0.11 | 0.75 ± 0.03 |
| YRH1511 | Late | 94.3 ± 24.2 | 47.6 ± 17.4 | 0.67 ± 0.05 | 0.70 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nichols, N.N.; Hector, R.E.; Mertens, J.A.; Frazer, S.E. Abatement of Inhibitors in Recycled Process Water from Biomass Fermentations Relieves Inhibition of a Saccharomyces cerevisiae Pentose Phosphate Pathway Mutant. Fermentation 2020, 6, 107. https://doi.org/10.3390/fermentation6040107
Nichols NN, Hector RE, Mertens JA, Frazer SE. Abatement of Inhibitors in Recycled Process Water from Biomass Fermentations Relieves Inhibition of a Saccharomyces cerevisiae Pentose Phosphate Pathway Mutant. Fermentation. 2020; 6(4):107. https://doi.org/10.3390/fermentation6040107
Chicago/Turabian StyleNichols, Nancy N., Ronald E. Hector, Jeffrey A. Mertens, and Sarah E. Frazer. 2020. "Abatement of Inhibitors in Recycled Process Water from Biomass Fermentations Relieves Inhibition of a Saccharomyces cerevisiae Pentose Phosphate Pathway Mutant" Fermentation 6, no. 4: 107. https://doi.org/10.3390/fermentation6040107
APA StyleNichols, N. N., Hector, R. E., Mertens, J. A., & Frazer, S. E. (2020). Abatement of Inhibitors in Recycled Process Water from Biomass Fermentations Relieves Inhibition of a Saccharomyces cerevisiae Pentose Phosphate Pathway Mutant. Fermentation, 6(4), 107. https://doi.org/10.3390/fermentation6040107