
Rhodaneses Enzyme Addition Could Reduce Cyanide Concentration and Enhance Fiber Digestibility via In Vitro Fermentation Study



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. Animals and Preparation of Rumen Inoculums
2.3. Incubation of Substrates
2.4. Analyses and Samples
2.5. Statistical Analysis
3. Results
3.1. Gas Kinetics and Ruminal Cyanide Concentration
3.2. pH, Ammonia-Nitrogen (NH3-N) and In Vitro Digestibility
3.3. Total Volatile Fatty Acids (TVFAs) and Their Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latif, S.; Zimmermann, S.; Barati, Z.; Müller, J. Detoxification of cassava leaves by thermal, sodium bicarbonate, enzymatic, and ultrasonic treatments. J. Food Sci. 2019, 84, 1986–1991. [Google Scholar] [CrossRef]
- Vennesland, B.; Castric, P.A.; Conn, E.E.; Solomonson, L.P.; Volini, M.; Westley, J. Cyanide metabolism. Fed. Proc. 1982, 41, 2639–2648. [Google Scholar]
- Cherdthong, A.; Khonkhaeng, B.; Seankamsorn, A.; Supapong, C.; Wanapat, M.; Gunun, N.; Gunun, P.; Chanjula, P.; Polyorach, S. Effects of feeding fresh cassava root with high-sulfur feed block on feed utilization, rumen fermentation and blood metabolites in Thai native cattle. Trop. Anim. Health Prod. 2018, 50, 1365–1371. [Google Scholar]
- Supapong, C.; Cherdthong, A. Effect of sulfur and urea fortification of fresh cassava root in fermented total mixed ration on the improvement milk quality of tropical lactating cows. Vet. Sci. 2020, 7, 98. [Google Scholar] [CrossRef]
- Supapong, C.; Cherdthong, A.; Wanapat, M.; Chanjula, P.; Uriyapongson, S. Effects of sulfur levels in fermented total mixed ration containing fresh cassava root on feed utilization, rumen characteristics, microbial protein synthesis and blood metabolites in Thai native beef cattle. Animals 2019, 9, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prachumchai, R.; Cherdthong, A.; Wanapat, M. Screening of cyanide-utilizing bacteria from rumen and in vitro evaluation of fresh cassava root utilization with pellet containing high sulfur diet. Vet. Sci. 2021, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Supapong, C.; Cherdthong, A. Effect of sulfur concentrations in fermented total mixed rations containing fresh cassava root on rumen fermentation. Anim. Prod. Sci. 2020, 60, 1429–1434. [Google Scholar] [CrossRef]
- Sumadong, P.; Cherdthong, A.; So, S.; Wanapat, M. Sulfur, fresh cassava root, and urea independently enhanced gas production, ruminal characteristics, and in vitro degradability. BMC Vet. Res. 2021, 17, 304. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Cherdthong, A.; Suntara, S.; Khota, W. Lactobacillus casei TH14 and additives could modulate the quality, gas kinetics and the in vitro digestibility of ensilaged rice straw. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1690–1703. [Google Scholar] [CrossRef]
- So, S.; Cherdthong, A.; Wanapat, M. Effect of sugarcane bagasse as industrial by-products treated Lactobacillus casei TH14, cellulase, and molasses on feed utilization, ruminal ecology and milk production of mid-lactating Holstein Friesian cows. J. Sci. Food Agric. 2021, 101, 4481–4489. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, T.C.; Kumar, V. Microbial remediation of cyanides. In Bioremediation Current Research and Application; Rathoure, A.K., Ed.; International Publishing House: New Delhi, India, 2017; pp. 88–110. [Google Scholar]
- Nazifi, S.; Aminlari, M.; Alaibakhsh, M.A. Distribution of rhodanese in tissues of goat (Capra hircus). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 134, 515–518. [Google Scholar] [CrossRef]
- Bordo, D.; Forlani, F.; Spallarossa, A.; Colnaghi, R.; Carpen, A.; Bolognesi, M.; Pagani, S. A persulfurated cysteine promotes active site reactivity in Azotobacter vinelandii rhodanese. Biol. Chem. 2001, 382, 1245–1252. [Google Scholar] [CrossRef]
- Cipollone, R.; Ascenzi, P.; Tomao, P.; Imperi, F.; Visca, P. Enzymatic detoxification of cyanide: Clues from Pseudomonas aeruginosa rhodanese. J. Mol. Microbiol. Biotechnol. 2008, 15, 199–211. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; AOAC: Arlington, VA, USA, 1998; Volume 2. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Ørskov, E.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Fisher, F.B.; Brown, J.S. Colorimetric determination of cyanide in stack gas and waste water. Anal. Chem. 1952, 24, 1440–1444. [Google Scholar] [CrossRef]
- Tilley, J.M.A.; Terry, R.A. A two-stage technique for the digestion of forage crops. J. Br. Grass. Soc. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- McMahon, J.M.; White, W.L.B.; Sayre, R.T. Cyanogenesis in cassava (Manihot esculanta Crantz). J. Exp. Bot. 1995, 46, 731–741. [Google Scholar] [CrossRef]
- Aminlari, M.; Malekhusseini, A.; Akrami, F.; Ebrahimnejad, H. Cyanide-metabolizing enzyme rhodanese in human tissues: Comparison with domestic animals. Com. Clin. Pathol. 2007, 16, 47–51. [Google Scholar] [CrossRef]
- Sornyotha, S.; Kyu, K.L.; Ratanakhanokchai, K. An efficient treatment for detoxification process of cassava starch by plant cell wall-degrading enzymes. J. Biosci. Bioeng. 2010, 109, 9–14. [Google Scholar] [CrossRef]
- Wanapat, M.; Cherdthong, A. Use of real-time PCR technique in studying rumen cellulolytic bacteria population as affected by level of roughage in swamp buffaloes. Curr. Microbiol. 2009, 58, 294–299. [Google Scholar] [CrossRef]
- Kondratovich, L.B.; Jhones, O.S.; Carly, A.H.; Michael, A.B.; Sara, J.T.; Pedro, R.B.C. Effects of dietary exogenous fibrolytic enzymes on ruminal fermentation characteristics of beef steers fed high- and low-quality growing diets. J. Anim. Sci. 2019, 97, 3089–3102. [Google Scholar] [CrossRef]
- Suttle, N.F. Mineral Nutrition of Livestock; CABI Publishing: Wallingford, UK, 2010. [Google Scholar]
- Brask-Pedersen, D.N.; Glitso, L.V.; Skov, L.K.; Lund, P.; Sehested, J. Effect of exogenous phytase on degradation of inositol phosphate in dairy cows. J. Dairy Sci. 2013, 96, 1691–1700. [Google Scholar] [CrossRef]
- Dagaew, G.; Cherdthong, A.; Wanapat, M.; So, S.; Polyorach, S. Ruminal fermentation, milk production efficiency and nutrient digestibility of lactating dairy cows receiving fresh cassava root and solid feed-block containing high sulfur. Fermentation 2021, 7, 114. [Google Scholar] [CrossRef]
- Prachumchai, R.; Cherdthong, A.; Wanapat, M.; Supapong, C.; Khonkhaeng, B.; Suntara, C.; Foiklang, S. Effect of feeding a pellet diet containing high sulphur with fresh cassava root supplementation on feed use efficiency, ruminal characteristics, and blood metabolites in Thai native beef cattle. J. Anim. Physiol. Anim. Nutr. 2021, 105, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Gunun, P.; Gunun, N.; Wanapat, M.; Cherdthong, A.; Polyorach, S.; Sirilaophaisan, S.; Wachirapakorn, C.; Kang, S. In vitro rumen fermentation and methane production as affected by rambutan pee powder. J. App. Anim. Res. 2018, 46, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Wanapat, M.; Cherdthong, A. Effect of banana flower powder supplementation as a rumen buffer on rumen fermentation efficiency and nutrient digestibility in dairy steers fed on high concentrate diet. Anim. Feed Sci. Technol. 2014, 196, 32–41. [Google Scholar] [CrossRef]
- So, S.; Cherdthong, A.; Wanapat, M.; Uriyapongson, S. Fermented sugarcane bagasse with Lactobacillus combined with cellulase and molasses promotes in vitro gas kinetics, degradability, and ruminal fermentation patterns compared to rice straw. Anim. Biotechnol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Item | Concentration, % DM |
|---|---|
| Ingredients | |
| Rice straw | 40.00 |
| Soybean meal | 8.35 |
| Palm kernel meal | 13.62 |
| Corn | 29.03 |
| Rice bran | 5.00 |
| Urea | 1.00 |
| Mineral premix * | 1.00 |
| Molasses, liquid | 2.00 |
| Chemical composition | |
| Dry matter (DM), % | 89.50 |
| Organic matter (OM), %DM | 90.23 |
| Ash, %DM | 9.77 |
| Crude protein, %DM | 12.00 |
| Neutral detergent fiber, %DM | 52.19 |
| Acid detergent fiber, %DM | 22.01 |
| Item | 300 ppm KCN | 450 ppm KCN | 600 ppm KCN | p-Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration of Rhodanese Enzymes (mg/104 ppm KCN) | ||||||||||||||||
| 0 | 0.65 | 1 | 1.35 | 0 | 0.65 | 1 | 1.35 | 0 | 0.65 | 1 | 1.35 | A | B | A * B | SEM | |
| Gas production kinetic | ||||||||||||||||
| a | −1.91 | −1.41 | −2.09 | −2.58 | −1.34 | −1.87 | −1.79 | −1.55 | −1.58 | −1.87 | −1.42 | −1.98 | 0.45 | 0.32 | 0.13 | 0.54 |
| b | 79.85 | 83.99 | 85.93 | 87.08 | 79.93 | 77.53 | 79.23 | 82.10 | 41.96 | 45.29 | 53.70 | 56.16 | 0.02 | 0.45 | 0.22 | 1.49 |
| c | 0.018 | 0.021 | 0.021 | 0.021 | 0.011 | 0.011 | 0.018 | 0.017 | 0.014 | 0.014 | 0.015 | 0.018 | 0.19 | 0.21 | 0.65 | 0.05 |
| a + b | 77.94 | 82.58 | 83.84 | 84.5 | 78.59 | 75.66 | 77.44 | 80.55 | 40.38 | 43.42 | 52.28 | 54.18 | 0.03 | 0.36 | 0.36 | 1.42 |
| Cumulative gas, mL | 82.51 | 87.15 | 92.41 | 90.07 | 83.16 | 80.23 | 86.01 | 85.12 | 44.95 | 47.99 | 60.85 | 58.75 | 0.01 | 0.04 | 0.77 | 1.83 |
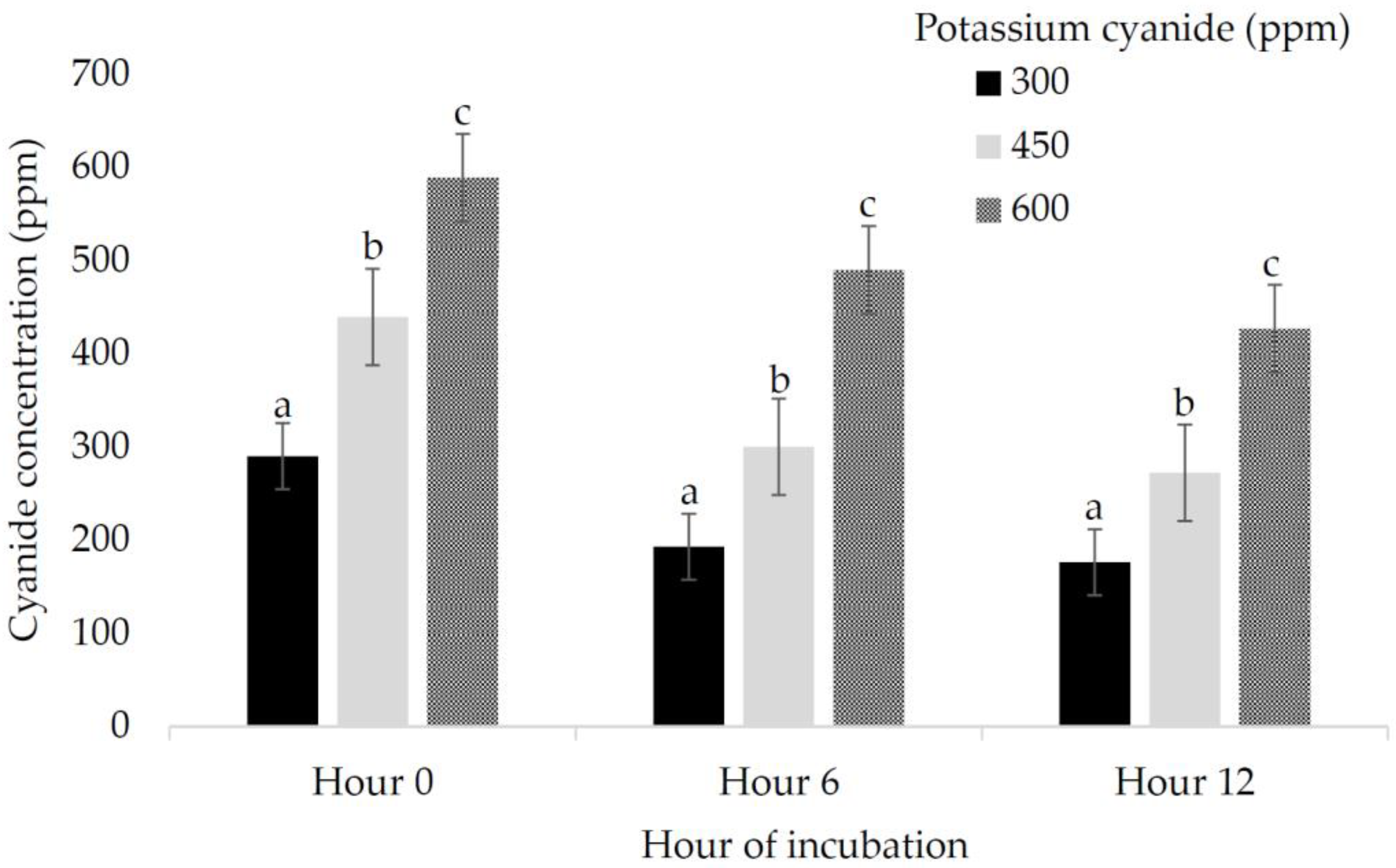
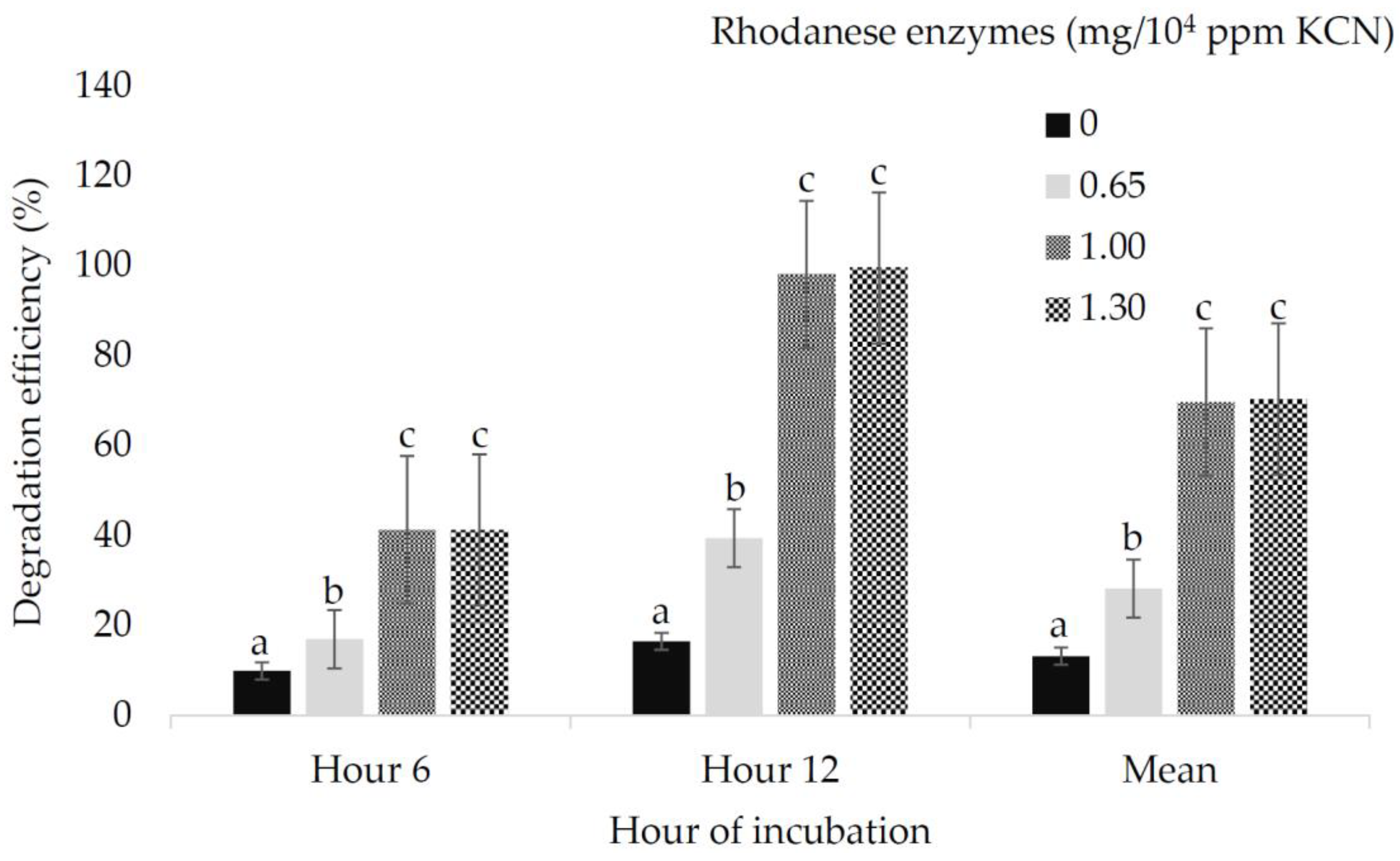
| Ruminal cyanide concentration, ppm | ||||||||||||||||
| H0 | 291.41 | 289.93 | 290.90 | 287.96 | 441.01 | 439.67 | 439.74 | 438.23 | 595.21 | 589.44 | 585.65 | 585.01 | 0.01 | 0.44 | 1.00 | 5.8 |
| H6 | 254.25 | 228.90 | 145.16 | 143.06 | 395.15 | 355.53 | 225.80 | 224.19 | 555.80 | 525.98 | 438.07 | 440.33 | 0.01 | 0.02 | 0.45 | 4.19 |
| H12 | 241.01 | 195.28 | 135.49 | 133.33 | 375.57 | 301.40 | 206.82 | 205.40 | 535.56 | 475.28 | 350.96 | 345.79 | 0.01 | 0.02 | 0.06 | 3.37 |
| Item | 300 ppm KCN | 450 ppm KCN | 600 ppm KCN | p-Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration of Rhodanese Enzymes (mg/104 ppm KCN) | ||||||||||||||||
| 0 | 0.65 | 1 | 1.35 | 0 | 0.65 | 1 | 1.35 | 0 | 0.65 | 1 | 1.35 | A | B | A * B | SEM | |
| pH | 6.99 | 6.98 | 6.99 | 6.98 | 7.02 | 7.03 | 7.02 | 7.04 | 7.07 | 7.06 | 7.05 | 7.07 | 0.11 | 0.97 | 0.99 | 0.53 |
| NH3-N, mg% | 11.88 | 11.99 | 12.21 | 12.33 | 13.10 | 13.58 | 13.55 | 13.44 | 14.01 | 14.09 | 15.64 | 17.81 | 0.02 | 0.97 | 0.19 | 1.07 |
| IVDMD, % | 67.43 | 67.87 | 73.08 | 75.59 | 64.34 | 65.21 | 70.84 | 72.83 | 47.52 | 53.04 | 60.54 | 62.69 | 0.02 | 0.03 | 0.94 | 1.51 |
| IVNDFD, % | 38.30 | 40.31 | 45.69 | 50.74 | 41.81 | 41.76 | 42.36 | 43.55 | 39.53 | 40.67 | 43.83 | 44.05 | 0.29 | 0.16 | 0.49 | 0.99 |
| IVADFD, % | 24.96 | 26.59 | 27.51 | 30.83 | 22.45 | 23.56 | 25.86 | 28.15 | 20.12 | 22.82 | 23.63 | 24.89 | 0.66 | 0.61 | 0.19 | 0.45 |
| Item | 300 ppm KCN | 450 ppm KCN | 600 ppm KCN | p-Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration of Rhodanese Enzymes (mg/104 ppm KCN) | ||||||||||||||||
| 0 | 0.65 | 1 | 1.35 | 0 | 0.65 | 1 | 1.35 | 0 | 0.65 | 1 | 1.35 | A | B | A * B | SEM | |
| Total VFA, mmol/L | 94.28 | 98.55 | 104.86 | 106.35 | 90.68 | 95.12 | 102.15 | 104.15 | 71.12 | 73.58 | 91.26 | 93.50 | 0.25 | 0.02 | 0.4 | 2.33 |
| VFA profiles, mol/100 mol | ||||||||||||||||
| Acetic acid | 64.18 | 59.49 | 66.95 | 65.22 | 68.52 | 64.74 | 63.56 | 64.48 | 66.71 | 66.54 | 67.92 | 60.19 | 0.79 | 0.52 | 0.53 | 2.24 |
| Propionic acid | 27.11 | 29.84 | 25.02 | 25.71 | 23.58 | 27.43 | 27.46 | 26.75 | 25.00 | 25.20 | 24.13 | 29.70 | 0.87 | 0.56 | 0.57 | 1.86 |
| Butyric acid | 8.73 | 10.68 | 8.03 | 9.08 | 7.90 | 7.83 | 9.00 | 8.78 | 8.30 | 8.26 | 7.95 | 10.13 | 0.55 | 0.52 | 0.46 | 1.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supapong, C.; Cherdthong, A. Rhodaneses Enzyme Addition Could Reduce Cyanide Concentration and Enhance Fiber Digestibility via In Vitro Fermentation Study. Fermentation 2021, 7, 207. https://doi.org/10.3390/fermentation7040207
Supapong C, Cherdthong A. Rhodaneses Enzyme Addition Could Reduce Cyanide Concentration and Enhance Fiber Digestibility via In Vitro Fermentation Study. Fermentation. 2021; 7(4):207. https://doi.org/10.3390/fermentation7040207
Chicago/Turabian StyleSupapong, Chanadol, and Anusorn Cherdthong. 2021. "Rhodaneses Enzyme Addition Could Reduce Cyanide Concentration and Enhance Fiber Digestibility via In Vitro Fermentation Study" Fermentation 7, no. 4: 207. https://doi.org/10.3390/fermentation7040207
APA StyleSupapong, C., & Cherdthong, A. (2021). Rhodaneses Enzyme Addition Could Reduce Cyanide Concentration and Enhance Fiber Digestibility via In Vitro Fermentation Study. Fermentation, 7(4), 207. https://doi.org/10.3390/fermentation7040207