
Purification and Characterization of Strong Simultaneous Enzyme Production of Protease and α-Amylase from an Extremophile-Bacillus sp. FW2 and Its Possibility in Food Waste Degradation





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Organic Compound Degrading-Bacteria
2.2. PCR Amplification, 16S rRNA Gene Sequencing, and Phylogenetic Analysis
2.3. Primary Screening of Potential Alkaline Protease- and Amylase-Producing Bacterial Isolate
2.4. Determination of Food Waste Degradation by Pure Bacterial Culture
2.5. Optimization of Enzyme Production
2.5.1. Optimization of Physio-Chemical Parameters
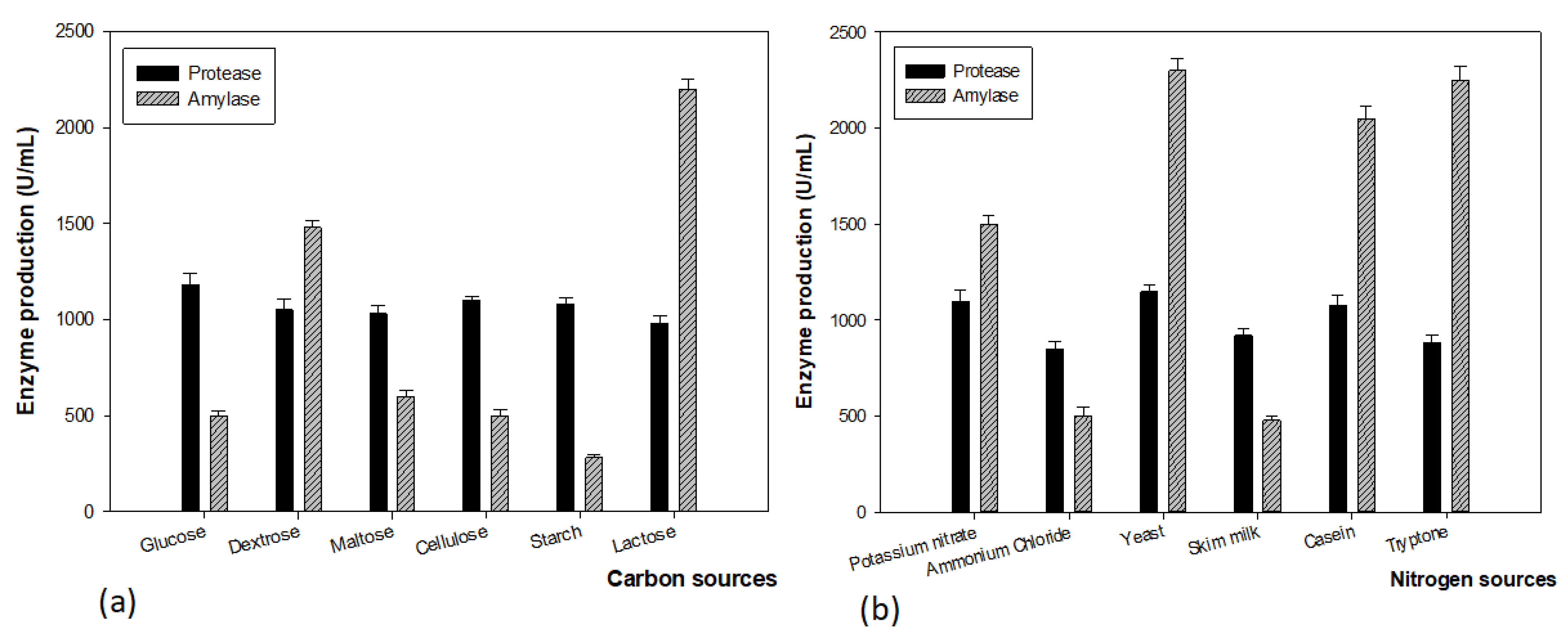
2.5.2. Effect of Carbon and Nitrogen Sources on Enzyme Production
2.6. Enzyme Assay
2.7. Purification of Protease and Amylase Enzymes
2.8. Estimation of Protein Content and Molecular Weight Determination of Enzymes
3. Results
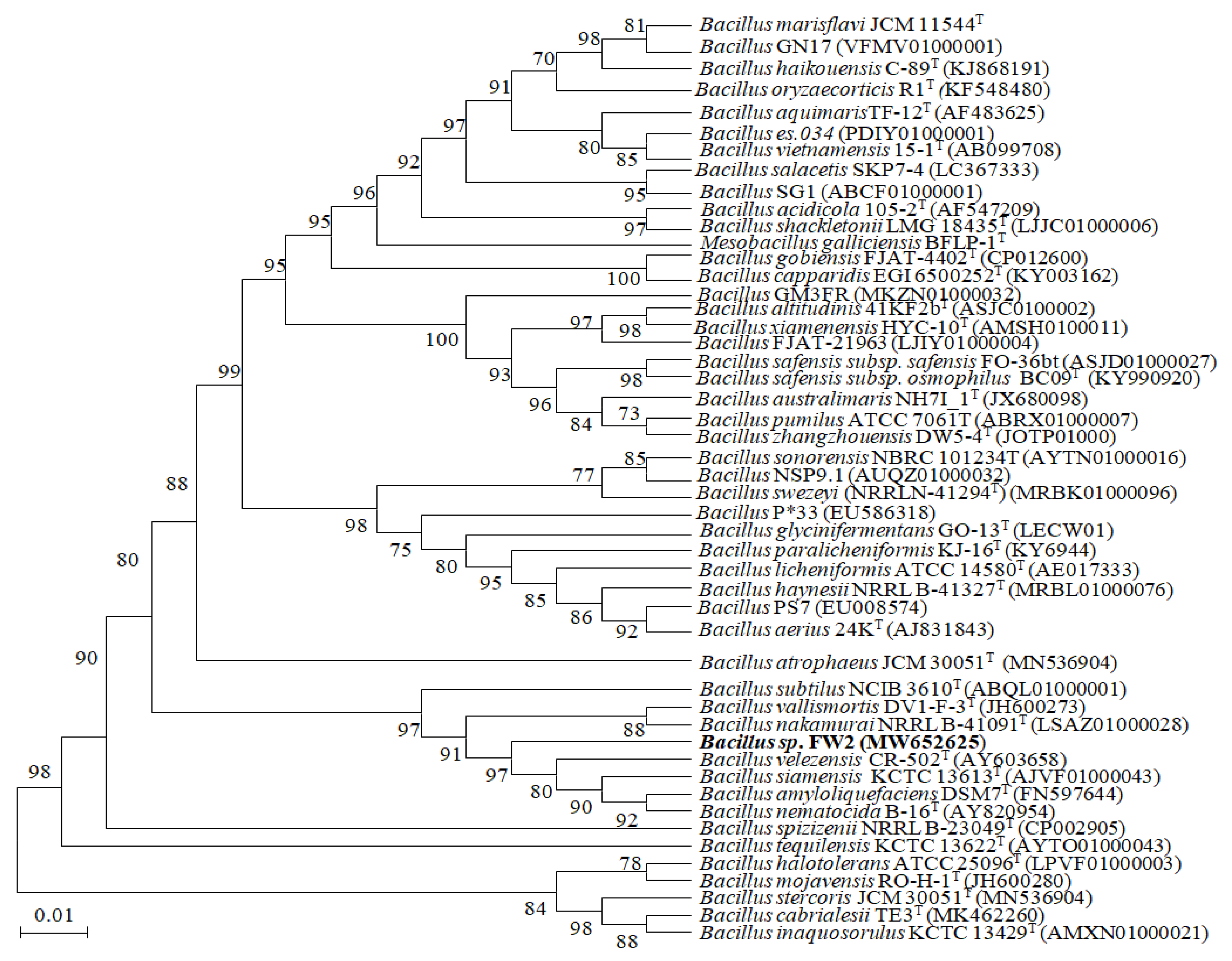
3.1. Physiology and the 16S rRNA Gene Analysis
3.2. Optimization of Fermentation for Enzyme Production
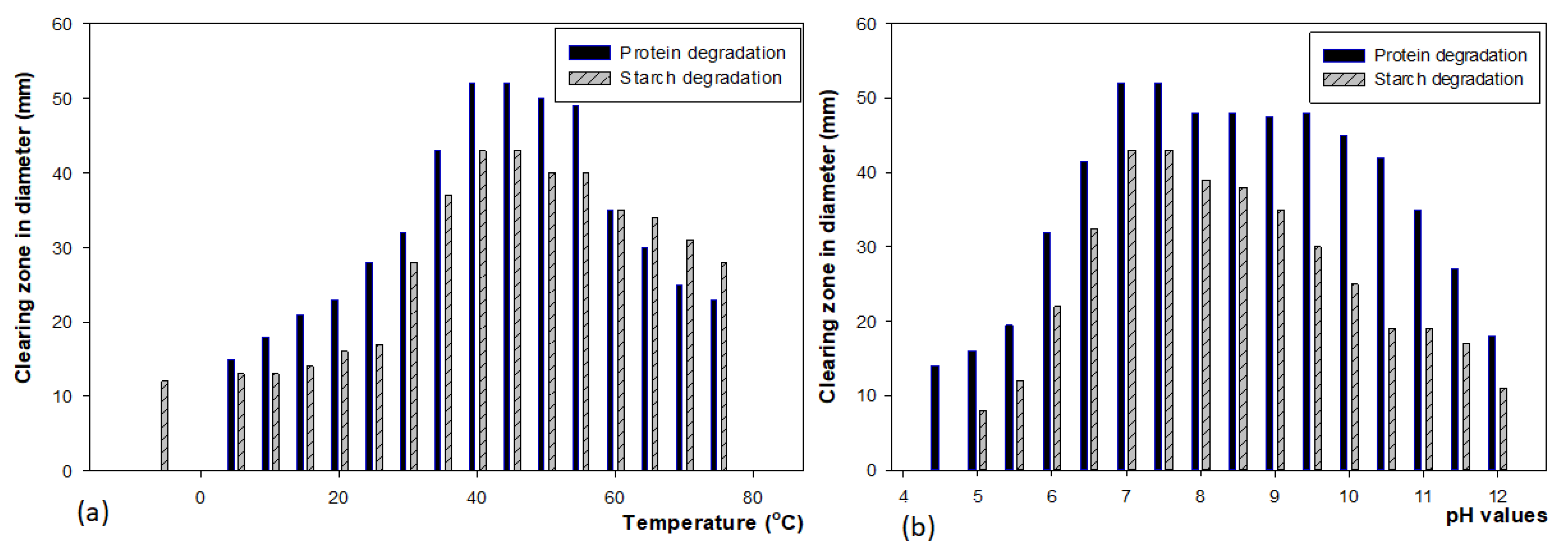
Optimization of Physio-Chemical Parameters
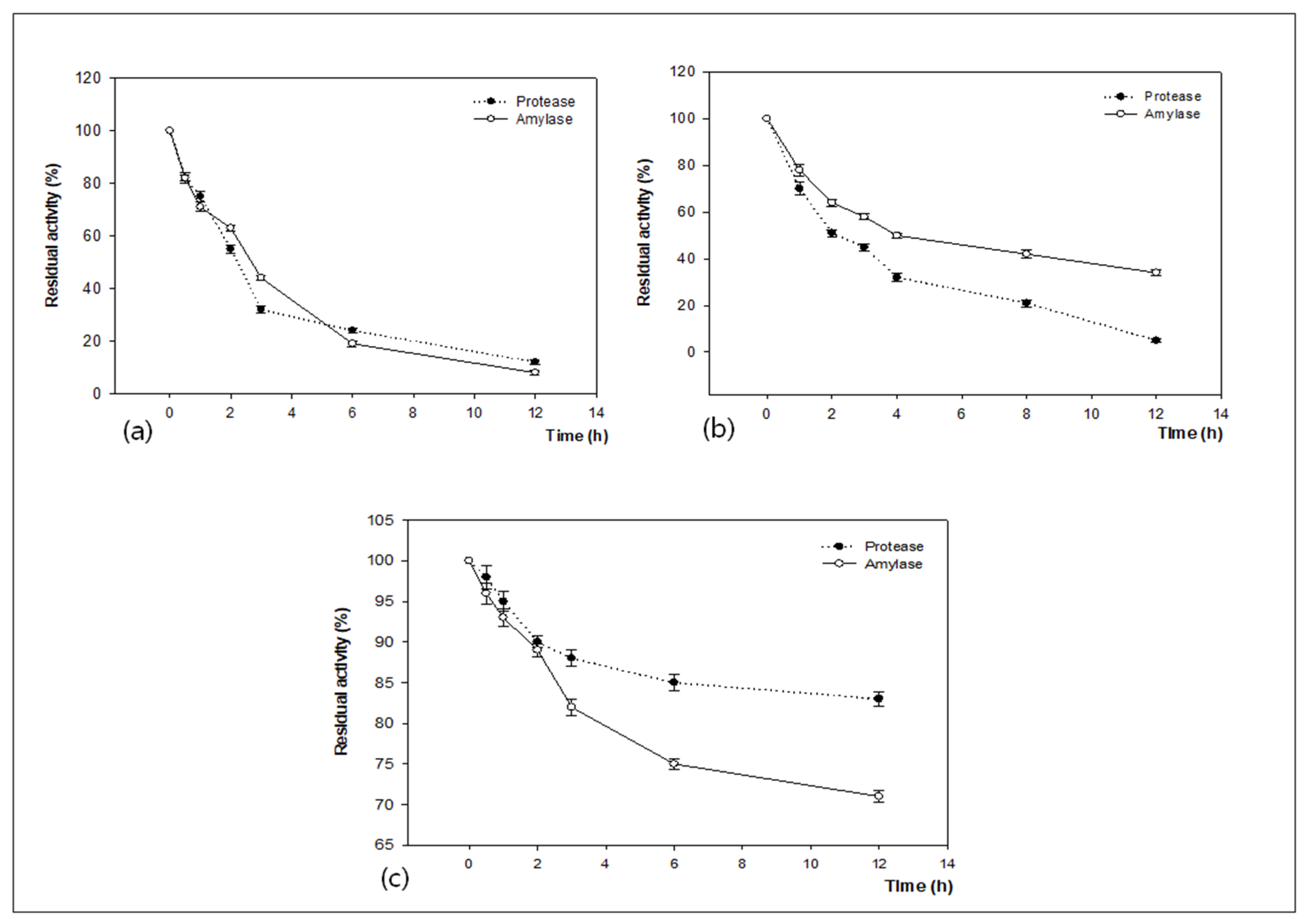
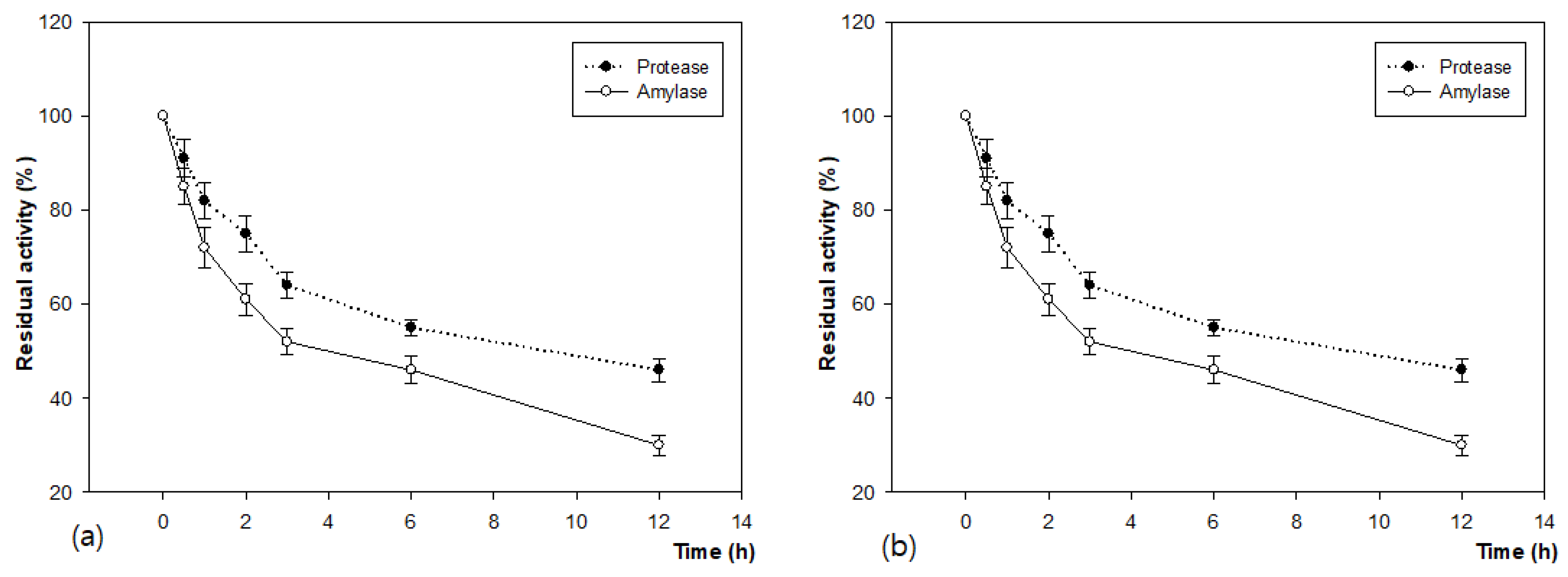
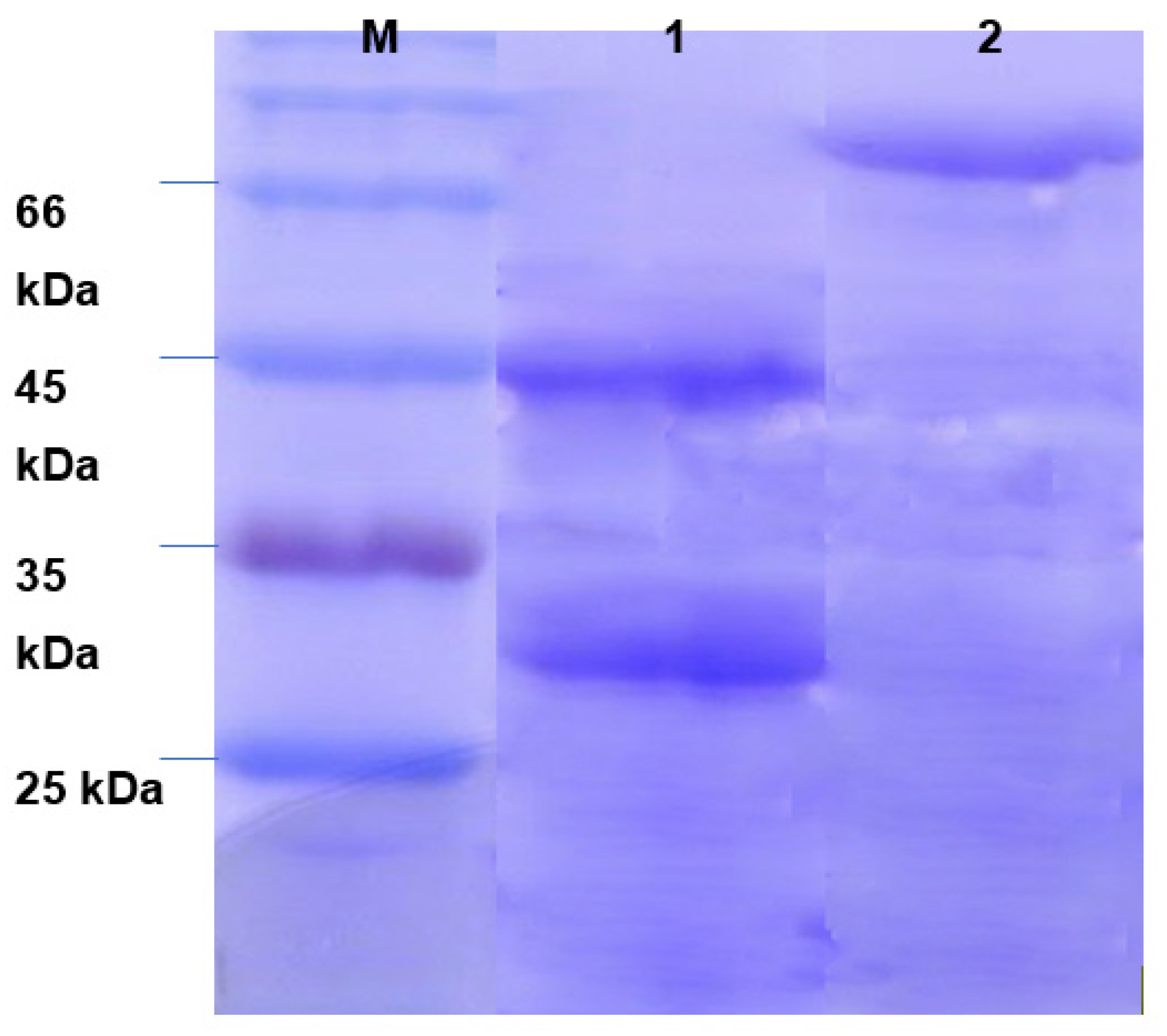
3.3. Determination of Molecular Weight of Produced Enzymes
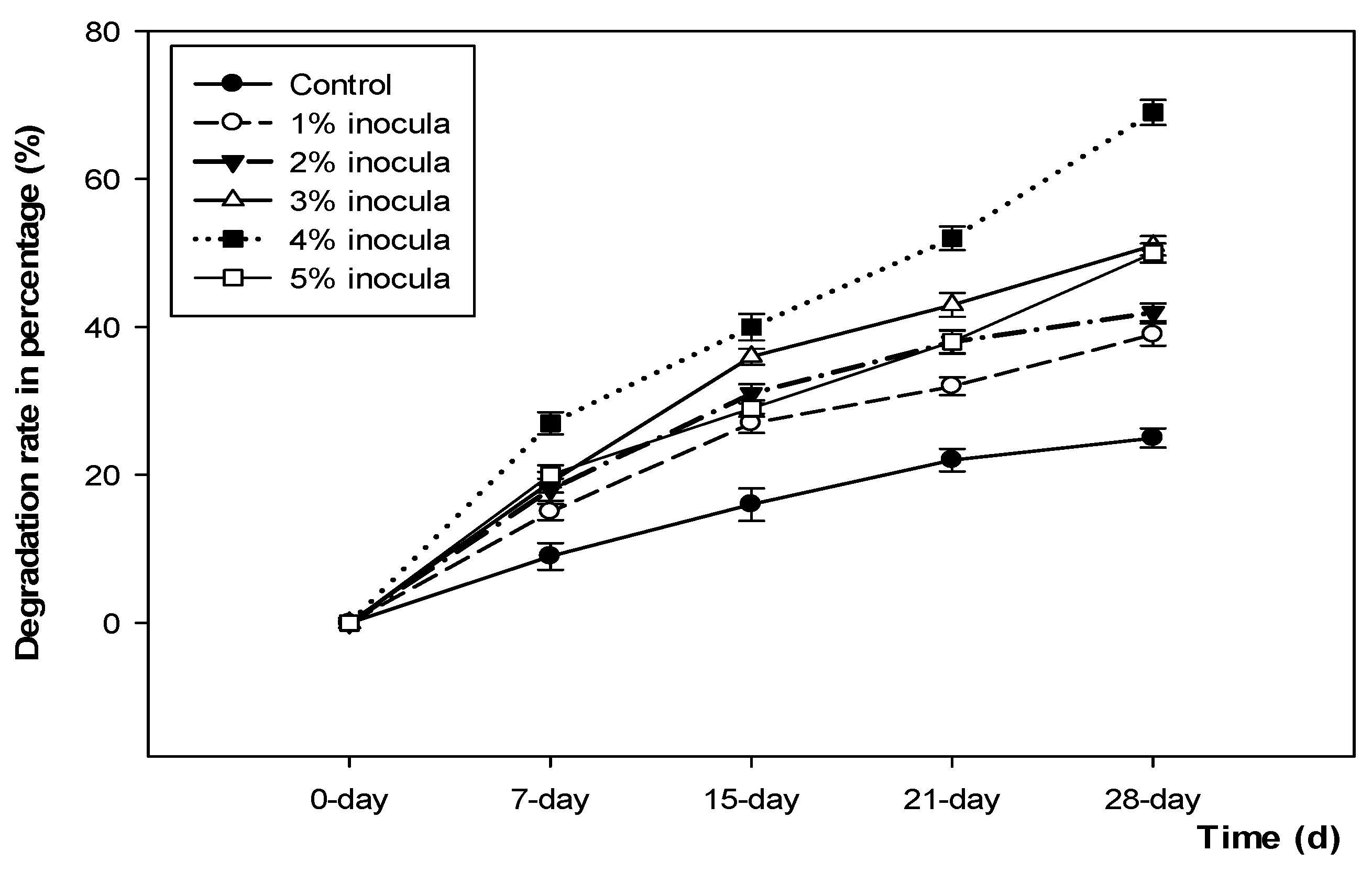
3.4. Degradation of Food Waste in Volume
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dumorné, K.; Camacho Córdova, D.; Astorga-Eló, M.; Renganathan, P. Extremozymes: A potential source for industrial applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Geng, A.; Cheng, Y.; Wang, Y.; Zhu, D.; Le, Y.; Wu, J.; Xie, R.; Yuan, J.S.; Sun, J. Transcriptome analysis of the digestive system of a wood-feeding termite (Coptotermes formosanus) revealed a unique mechanism for effective biomass degradation. Biotechnol. Biofuels 2018, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial Proteases Applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Gurung, N.; Ray, S.; Bose, S.; Rai, V. A Broader View: Microbial Enzymes and Their Relevance in Industries, Medicine, and Beyond. BioMed Res. Int. 2013, 2013, 329121. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, X.; Qiu, J.; Wang, K.; Meng, K.; Luo, H.; Su, X.; Ma, R.; Huang, H.; Yao, B. Development of Bacillus amyloliquefaciens as a high-level recombinant protein expression system. J. Ind. Microbiol. Biotechnol. 2019, 46, 113–123. [Google Scholar] [CrossRef]
- Zhao, H.L.; Chen, X.L.; Xie, B.B.; Zhou, M.Y.; Gao, X.; Zhang, X.Y.; Zhou, B.C.; Weiss, A.S.; Zhang, Y.Z. Elastolytic mechanism of a novel M23 metalloprotease pseudoalterin from deep-sea Pseudoalteromonas sp. CF6-2: Cleaving not only glycyl bonds in the hydrophobic regions but also peptide bonds in the hydrophilic regions involved in cross-linking. J. Biol. Chem. 2012, 287, 39710–39720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, C.; Gemechu, G.; Tafesse, M. Isolation, screening, characterization, and identification of alkaline protease-producing bacteria from leather industry effluent. Ann. Microbiol. 2021, 71, 24. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Santana, J.C.C.; Tambourgi, E.B. Protease (bromelain): The effect of pH on Bromelian Partition from Ananas comosus by PEG4000/Phosphate ATPS. Braz. Arch. Biol. Technol. 2011, 54, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Silveira, E.; Souza-Jr, M.E.; Santana, J.C.C.; Chaves, A.C.; Porto, L.F.; Tambourgi, E.B. Expanded bed adsorption of bromelain (E.C. 3.4.22.33) from Ananas comosus crude extract. Braz. J. Chem. Eng. 2009, 26, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Raval, V.H.; Pillai, S.; Rawal, C.M.; Signgh, S.P. Biochemical and structural characterization of a detergent-stable serine alkaline protease from seawater haloalkaliphilic bacteria. Process Biochem. 2014, 49, 955–962. [Google Scholar] [CrossRef]
- Ibrahim, A.S.S.; Al-Salamah, A.A.; El-Badawi, Y.B.; El-Tayeb, M.A. Detergent-, solvent- and salt-compatible thermoactive alkaline serine protease from halotolerant alkaliphilic Bacillus sp. NPST-AK15: Purification and characterization. Extremophiles 2015, 19, 961–971. [Google Scholar] [CrossRef]
- Sharanappa, A.; Wani, K.; Pallavi, P. Bioprocessing of food industrial waste for α-amylase production by solid state fermentation. Int. J. Adv. Biotechnol. Res. 2011, 2, 473–480. [Google Scholar]
- Sanghvi, G.V.; Koyani, R.D.; Rajput, K.S. Isolation, optimization, and partial purification of amylase from Chrysosporium asperatum by submerged fermentation. J. Microbiol. Biotechnol. 2011, 21, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Arunkumar, P.; Thanalakshmi, M.; Kumar, P.; Premkumar, K. Micrococcus luteus mediated dual mode synthesis of gold nanoparticles: Involvement of extracellular α-amylase and cell wall teichuronic acid. Colloids Surf. B 2013, 103, 517–522. [Google Scholar] [CrossRef]
- Yao, D.; Su, L.; Li, N.; Wu, J. Enhanced extracellular expression of Bacillus stearothermophilus α-amylase in Bacillus subtilis through signal peptide optimization, chaperone overexpression and α-amylase mutant selection. Microb. Cell Fact. 2019, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Kizhakedathil, M.P.J.; Subathra, D.C. Acid stable α-amylase from Pseudomonas balearica VITPS19—Production, purification and characterization. Biotechnol. Rep. 2021, 30, e00603. [Google Scholar] [CrossRef] [PubMed]
- Solihin, J.; Waturangi, D.E.; Purwadaria, T. Induction of amylase and protease as antibiofilm agents by starch, casein, and yeast extract in Arthrobacter sp. CW01. BMC Microbiol. 2021, 21, 232. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Ahn, J.Y.; Ro, Y.H.; Ravindran, B.; Kim, J.S.; Chang, S.W.; Shim, J.H.; Chung, W.J. The efficiency of potential food waste-degrading bacteria under harsh conditions. J. Appl. Microbiol. 2021, 132, 340–350. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Ahn, J.; Kim, J.; Lee, S.; Lee, I.; Kim, S.; Chang, S.; Chung, W. Volatile Fatty Acid Production from Food Waste Leachate Using Enriched Bacterial Culture and Soil Bacteria as Co-Digester. Sustainability 2021, 13, 9606. [Google Scholar] [CrossRef]
- de Carvalho, R.V.; Côrrea, T.L.R.; Silva, J.C.M.; Mansur, L.R.C.O.; Martins, M.L.L. Properties of an amylase from thermophilic Bacillus sp. Braz. J. Microbiol. 2008, 39, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevillano, L.; Vijgenboom, E.; van Wezel, G.P.; Díaz, M.; Santamaría, R.I. New approaches to achieve high level enzyme production in Streptomyces lividans. Microb. Cell Fact. 2016, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Nithya, K.; Muthukumar, C.; Kadaikunnan, S.; Alharbi, N.S.; Khaled, J.M.; Dhanasekaran, D. Purification, characterization, and statistical optimization of a thermostable α-amylase from desert actinobacterium Streptomyces fragilis DA7-7. 3 Biotech. 2017, 7, 350. [Google Scholar] [CrossRef]
- Biazus, J.P.M.; Souza, R.R.; Márquez, J.E.; Franco, T.T.; Santana, J.C.C.; Tambourgi, E.B. Production and Characterization of Amylases from Zea mays Malt. Braz. Arch. Biol. Technol. 2009, 52, 991–1000. [Google Scholar] [CrossRef] [Green Version]
- Toledo, A.L.; Severo Junior, J.B.; Souza, R.R.; Silveira, E.; Santana, J.C.C.; Tambourgi, E.B. Purification by expanded bed adsorption and characterization of an α-amylases FORILASE NTL® from A. niger. J. Chromatogr. B 2007, 846, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Santana, J.C.C.; Ehrhardt, D.D.; Tambourgi, E.B. Optimizing of alcohol production from manioc starch. Food Sci. Technol. 2010, 30, 613–617. [Google Scholar]
- Van Pham, H.T.; Kim, J. Bacillus thaonhiensis sp. nov., a new species, was isolated from the forest soil of Kyonggi University by using a modified culture method. Curr. Microbiol. 2014, 68, 88–95, Erratum in Curr. Microbiol. 2014, 69, 225. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Improvement of hydrogen production during anaerobic fermentation of food waste leachate by enriched bacterial culture using biochar as an additive. Microorganisms 2021, 9, 2438. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limit on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Amoozegar, M.A.; Malekzadeh, F.; Malik, K.A. Production of amylase by newly isolated moderate halophile, Halobacillus sp. strain MA-2. J. Microbiol. Methods 2003, 52, 353–359. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases, α and ρ. Methods Enzymol. 1955, 1, 149–158. [Google Scholar]
- Green, A.; Hughes, W. Protein fractionation on the basis of solubility in aqueous solutions of salts and organic solvents. Methods Enzymol. 1955, 1, 67–90. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Borriss, R.; Chen, X.H.; Rueckert, C.; Blom, J.; Becker, A.; Baumgarth, B.; Fan, B.; Pukall, R.; Schumann, P.; Spröer, C.; et al. Relationship of Bacillus amyloliquefaciens clades associated with strains DSM 7T and FZB42T: A proposal for Bacillus amyloliquefaciens subsp. amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on complete genome sequence comparisons. Int. J. Syst. Evol. Microbiol. 2011, 61, 1786–1801. [Google Scholar]
- Ruiz-Garcı´a, C.; Be´jar, V.; Martı´nez-Checa, F.; Llamas, I.; Quesada, E. Bacillus velezensis sp. nov., a surfactantproducing bacterium isolated from the river Ve´lez in Ma´laga, southern Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Sonika Gupta, S.; Sharma, P.; Dev, K.; Sourirajan, A. Halophilic Bacteria of Lunsu produce an array of industrially important enzymes with salt tolerant activity. Biochem. Res. Int. 2016, 2016, 9237418. [Google Scholar]
- de Veras, B.O.; dos Santos, Y.Q.; Diniz, K.M.; Carelli, G.S.C.; dos Santos, E.A. Screening of protease, cellulase, amylase and xylanase from the salt-tolerant and thermostable marine Bacillus subtilis strain SR60 [version 1; peer review: 1 approved with reservations]. F1000Research 2018, 7, 1704. [Google Scholar] [CrossRef]
- Kim, J.Y.; Hur, S.H.; Hong, J.H. Purification and characterization of an alkaline cellulase from a newly isolated alkalophilic Bacillus sp. HSH-810. Biotechnol. Lett. 2005, 27, 313–316. [Google Scholar] [CrossRef]
- Tahir, S.R.; Bakhsh, A.; Rao, A.Q.; Naz, M.; Sleem, M. Isolation, purification and characterization of extracellular β-glucosidase from Bacillus sp. Adv. Environ. Biol. Rep. 2009, 3, 269–277. [Google Scholar]
- Panda, M.K.; Sahu, M.K.; Tayung, K. Isolation and characterization of a thermophilic Bacillus sp. with protease activity isolated from hot spring of Tarabalo, Odisha. India. Iran. J. Microbiol. 2013, 5, 159–165. [Google Scholar] [PubMed]
- Jogezai, N.; Raza, A.; Abbas, F.; Bajwa, M.; Mohammad, D.; Kakar, W.; Saeed, M.; Awan, A. Optimization of cultural conditions for microbial alpha amylase production. J. Microbiol. Antimicrob. 2011, 3, 221–227. [Google Scholar]
- Gerday, C. Extremophiles: Basic concepts. Knowledge for Sustainable Development. In An Insight into the Encyclopedia of Life Support Systems; UNESCO Publishing/EOLSS Publishers: Oxford, UK, 2002; Volume 1, pp. 573–598. [Google Scholar]
- Gomes, J.; Steiner, W. The biocatalytic potential of extremophiles and extremozymes. Food Technol. Biotech. 2004, 42, 223–235. [Google Scholar]
- Gupta, G.N.; Srivastava, S.; Khare, S.K.; Prakash, V. Extremophiles: An: Overview of microorganism from extreme environment. Int. J. Agric. Environ. Biotech. 2014, 7, 371–380. [Google Scholar] [CrossRef]
- Smith, E.L.; Markland, F.S.; Kasper, C.B.; DeLange, R.J.; Landon, M.; Evans, W.H. The complete amino acid sequence of two types of subtilisin, BPN’ and Carlsberg. J. Biol. Chem. 1966, 241, 5974–5976. [Google Scholar] [CrossRef]
- Jacobs, M.; Eliasson, M.; Uhlén, M.; Flock, J.I. Cloning, sequencing and expression of subtilisin Carlsberg from Bacillus licheniformis. Nucleic Acids Res. 1985, 13, 8913–8926. [Google Scholar] [CrossRef] [Green Version]
- Setyorini, E.; Takenaka, S.; Murakami, S.; Aoki, K. Purification and characterization of two novel halotolerant extracellular protease from Bacillus subtilis strain FP-133. Biosci. Biotechnol. Biochem. 2006, 70, 433–440. [Google Scholar] [CrossRef] [Green Version]
- Deng, A.; Wu, J.; Zhang, Y.; Zhang, G.; Wen, T. Purification and characterization of a surfactant-stable high-alkaline protease from Bacillus sp. B001. Bioresour. Technol. 2010, 101, 7100–7106. [Google Scholar] [CrossRef]
- Adekoya, O.A.; Sylte, I. The thermolysin family (M4) of enzymes: Therapeutic and biotechnological potential. Chem. Biol. Drug Des. 2009, 73, 7–16. [Google Scholar] [CrossRef]
- Bano, S.; Ul Qader, S.A.; Aman, A.; Syed, M.N.; Azhar, A. Purification and characterization of novel α-amylase from Bacillus subtilis KIBGE HAS. AAPS PharmSciTech 2011, 12, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Du, R.; Song, Q.; Zhang, Q.; Zhao, F.; Kim, R.C.; Zhou, Z.; Han, Y. Purification and characterization of novel thermostable and Ca-independent α-amylase produced by Bacillus amyloliquefaciens BH072. Int. J. Biol. Macromol. 2018, 115, 1151–1156. [Google Scholar] [CrossRef]
- Wang, H.; Yang, L.; Ping, Y.; Bai, Y.; Luo, H.; Huang, H.; Yao, B. Engineering of a Bacillus amyloliquefaciens strain with high neutral protease producing capacity and optimization of its fermentation conditions. PLoS ONE 2016, 11, e0146373. [Google Scholar] [CrossRef]
- Vehmaanperä, J.; Steinborn, G.; Hofemeister, J. Genetic manipulation of Bacillus amyloliquefaciens. J. Biotechnol. 1991, 19, 221–240. [Google Scholar] [CrossRef]
- Xia, W.L.; Huang, H.K.; Qi, Z.P.; Wang, Q.S. Experimental studies on dairy manure treatment by static bed composting and microbe reagent inoculating. Trans. CSAE 2006, 22, 215–219. [Google Scholar]
- Magrí, A.; Teira-Esmatges, M.R. Assessment of a composting process for the treatment of beef cattle manure treatment of beef cattle manure. J. Environ. Sci. Health B 2015, 50, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Liu, J.; Guo, H.; Li, E.; Yan, Y. Characteristics and optimization of dairy manure composting for reuse as a dairy mattress in areas with large temperature differences. J. Clean Prod. 2019, 232, 1053–1061. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, V.H.T.; Kim, J.; Shim, J.; Chang, S.; Chung, W. Purification and Characterization of Strong Simultaneous Enzyme Production of Protease and α-Amylase from an Extremophile-Bacillus sp. FW2 and Its Possibility in Food Waste Degradation. Fermentation 2022, 8, 12. https://doi.org/10.3390/fermentation8010012
Pham VHT, Kim J, Shim J, Chang S, Chung W. Purification and Characterization of Strong Simultaneous Enzyme Production of Protease and α-Amylase from an Extremophile-Bacillus sp. FW2 and Its Possibility in Food Waste Degradation. Fermentation. 2022; 8(1):12. https://doi.org/10.3390/fermentation8010012
Chicago/Turabian StylePham, Van Hong Thi, Jaisoo Kim, Jeahong Shim, Soonwoong Chang, and Woojin Chung. 2022. "Purification and Characterization of Strong Simultaneous Enzyme Production of Protease and α-Amylase from an Extremophile-Bacillus sp. FW2 and Its Possibility in Food Waste Degradation" Fermentation 8, no. 1: 12. https://doi.org/10.3390/fermentation8010012
APA StylePham, V. H. T., Kim, J., Shim, J., Chang, S., & Chung, W. (2022). Purification and Characterization of Strong Simultaneous Enzyme Production of Protease and α-Amylase from an Extremophile-Bacillus sp. FW2 and Its Possibility in Food Waste Degradation. Fermentation, 8(1), 12. https://doi.org/10.3390/fermentation8010012