
Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria



Abstract
:1. Introduction
2. Classification of LAB Bacteriocins
3. Sequence Properties of Identified Class II Bacteriocins
4. Distribution of Class II Bacteriocins in Food-Associated LAB
4.1. Approaches for Studying Bacteriocin Diversity
4.2. In Silico Prediction of the Distribution of Class II Bacteriocins in LAB
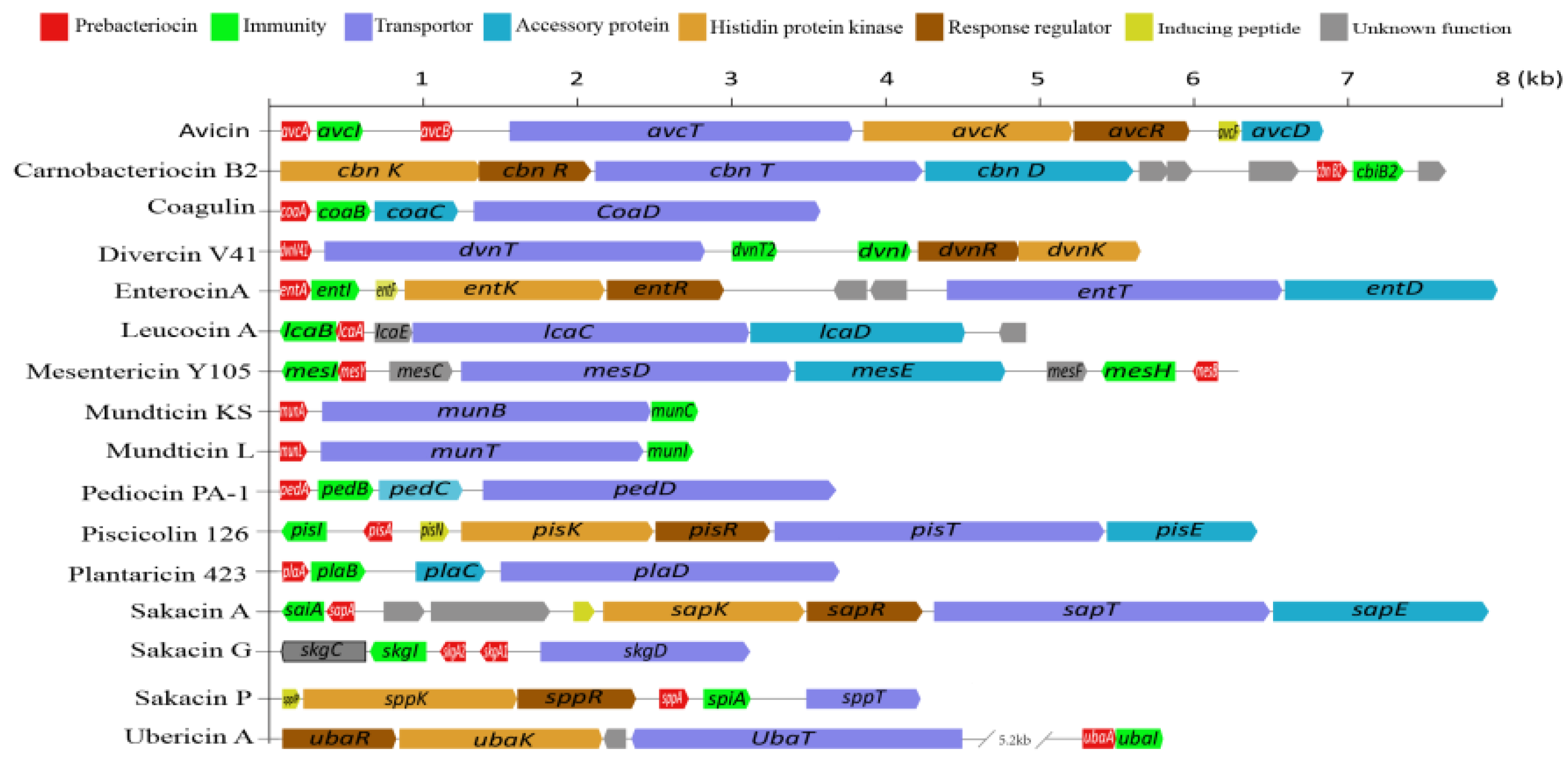
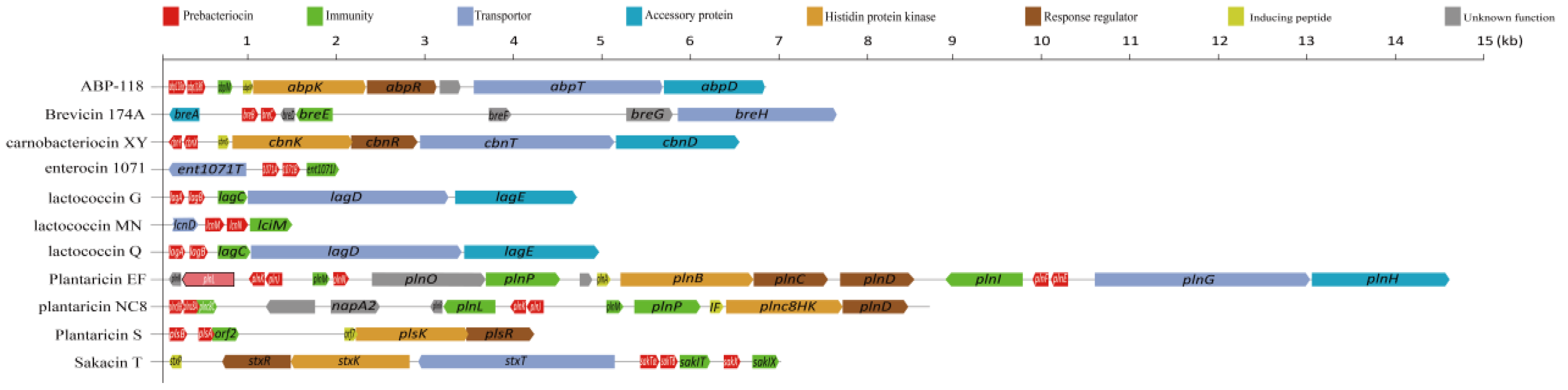
4.2.1. Carnobacteria
4.2.2. Enterococci
4.2.3. Lactobacilli Group
4.2.4. Lactococci
4.2.5. Leuconostoc Spp.
4.2.6. Pediococci
4.2.7. Others
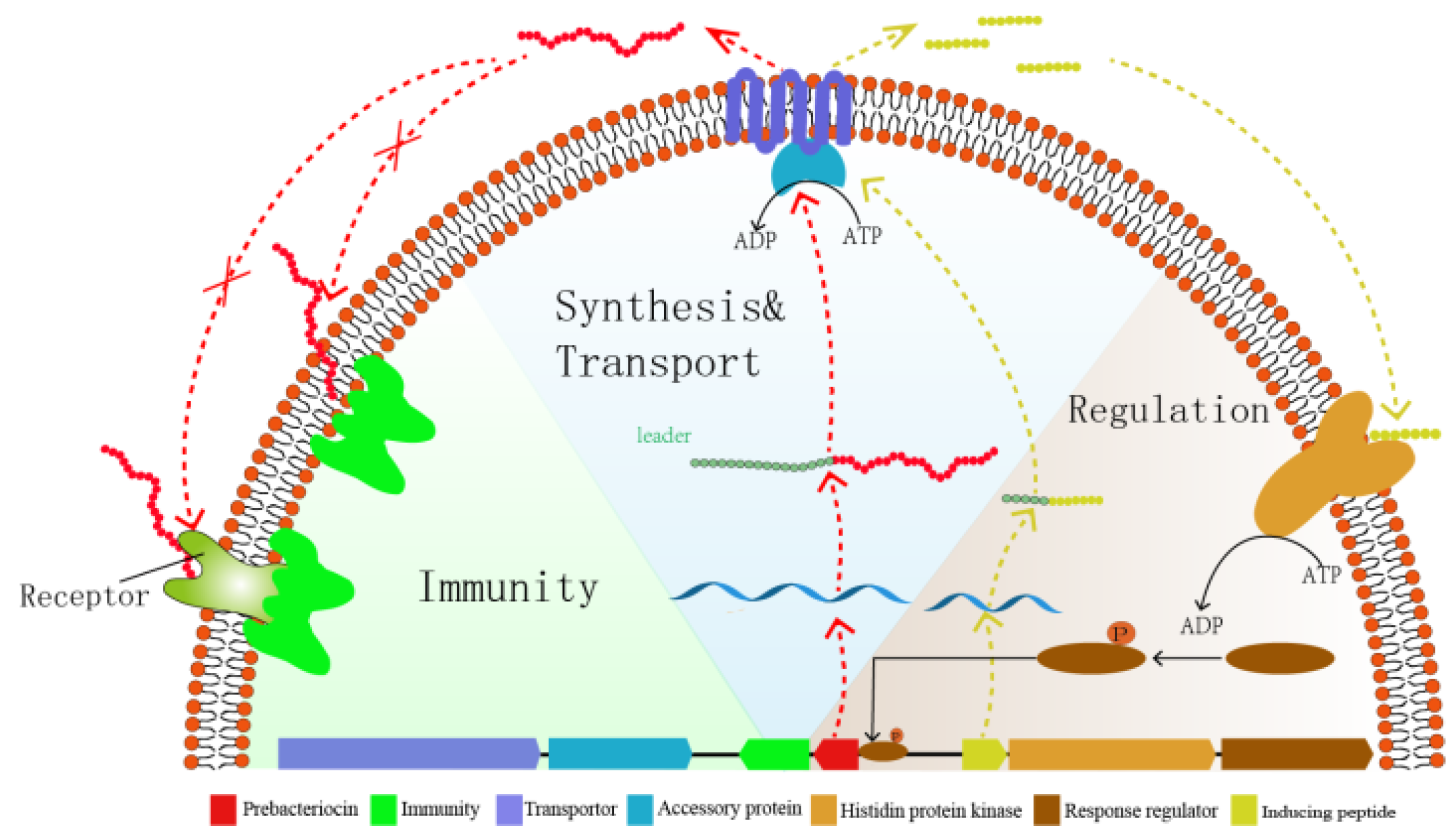
5. Biosynthesis and Genetics
5.1. IIa
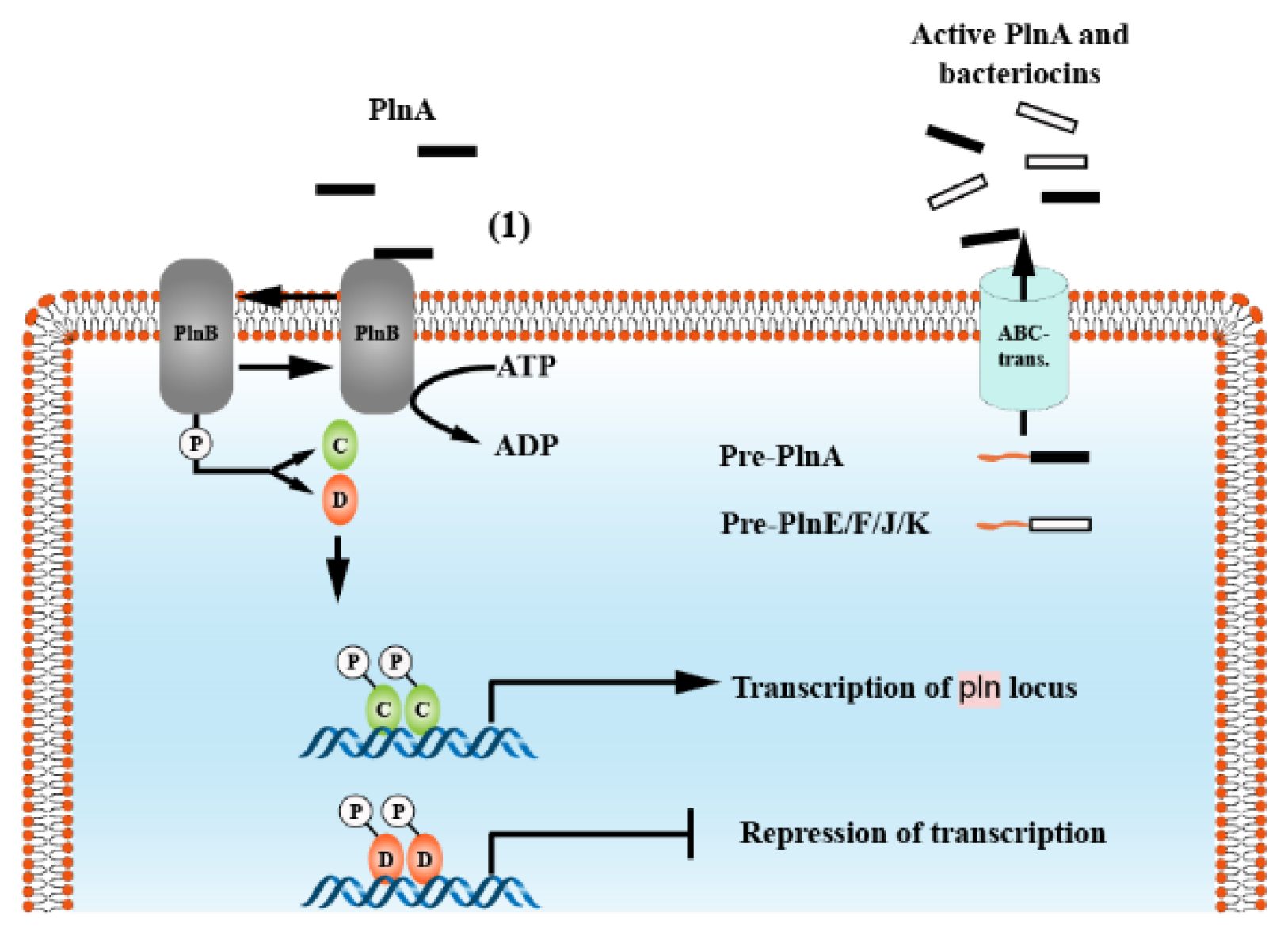
5.2. IIb
5.3. IIc
5.4. IId
6. Production of Class II Bacteriocins by Microbial Fermentation
6.1. Natural Fermentation
6.2. Improving Class II Bacteriocin Production by Optimizing Fermentation Conditions
6.3. Improving Class II Bacteriocin Production by Genetic Engineering
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruiz Rodríguez, L.G.; Mohamed, F.; Bleckwedel, J.; Medina, R.; De Vuyst, L.; Hebert, E.M.; Mozzi, F. Diversity and functional properties of lactic acid bacteria isolated from wild fruits and flowers present in Northern Argentina. Front. Microbiol. 2019, 10, 1091. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Li, Y.; Xu, L.; Wu, D.; Zhou, Q.; Li, P.; Gu, Q. A novel bacteriocin from Lactobacillus pentosus ZFM94 and its antibacterial mode of action. Front. Nutr. 2021, 8, 710862. [Google Scholar] [CrossRef] [PubMed]
- Umu, Ö.C.; Bäuerl, C.; Oostindjer, M.; Pope, P.B.; Hernandez, P.E.; Perez-Martinez, G.; Diep, D.B. The potential of class II bacteriocins to modify gut microbiota to improve host health. PLoS ONE 2016, 11, e0164036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yount, N.Y.; Weaver, D.C.; de Anda, J.; Lee, E.Y.; Lee, M.W.; Wong, G.C.L.; Yeaman, M.R. Discovery of Novel Type II Bacteriocins Using a New High-Dimensional Bioinformatic Algorithm. Front. Immunol. 2020, 11, 1873. [Google Scholar] [CrossRef]
- Ishibashi, N.; Himeno, K.; Masuda, Y.; Perez, R.H.; Iwatani, S.; Zendo, T.; Wilaipun, P.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K.J.A.; et al. Gene cluster responsible for secretion of and immunity to multiple bacteriocins, the NKR-5-3 enterocins. Appl. Environ. Microbiol. 2014, 80, 6647–6655. [Google Scholar] [CrossRef] [Green Version]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalban-Lopez, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Heng, N.C.; Wescombe, P.A.; Burton, J.P.; Jack, R.W.; Tagg, J.R. The diversity of bacteriocins in Gram-positive bacteria. In Bacteriocins: Ecology and Evolution; Riley, M.A., Chavan, M.A., Eds.; Springer: Berlin/Heidelberg, Gertmany, 2007; pp. 45–92. [Google Scholar]
- Heng, N.C.K.; Tagg, J.R. What’s in a name? Class distinction for bacteriocins. Nat. Rev. Microbiol. 2006, 4, 160. [Google Scholar] [CrossRef]
- Willey, J.M.; van der Donk, W.A. Lantibiotics: Peptides of diverse structure and function. Annu. Rev. Microbiol. 2007, 61, 477–501. [Google Scholar] [CrossRef]
- Sandiford, S.K. An overview of lantibiotic biosynthetic machinery promiscuity and its impact on antimicrobial discovery. Expert Opin. Drug Discov. 2020, 15, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAuliffe, O.; Ross, R.P.; Hill, C. Lantibiotics: Structure, biosynthesis and mode of action. FEMS Microbiol. Rev. 2001, 25, 285–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennahar, S.; Sashihara, T.; Sonomoto, K.; Ishizaki, A. Class IIa bacteriocins: Biosynthesis, structure and activity. FEMS Microbiol. Rev. 2000, 24, 85–106. [Google Scholar] [PubMed]
- Nissen-Meyer, J.; Oppegård, C.; Rogne, P.; Haugen, H.S.; Kristiansen, P.E. Structure and mode-of-action of the two-peptide (class-IIb) bacteriocins. Probiotics Antimicrob. Proteins 2010, 2, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and Leaderless Bacteriocins: Biosynthesis, Mode of Action, Applications, and Prospects. Front. Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef]
- Iwatani, S.; Zendo, T.; Sonomoto, K. Class IId or linear and non-pediocin-like bacteriocins. In Prokaryotic Antimicrobial Peptides; Springer: Berlin/Heidelberg, Gertmany, 2011; pp. 237–252. [Google Scholar]
- Yi, Y.; Li, P.; Zhao, F.; Zhang, T.; Shan, Y.; Wang, X.; Liu, B.; Chen, Y.; Zhao, X.; Lü, X. Current status and potentiality of class II bacteriocins from lactic acid bacteria: Structure, mode of action and applications in the food industry. Trends Food Sci. Technol. 2022, 120, 387–401. [Google Scholar] [CrossRef]
- Kanatani, K.; Oshimura, M.; Sano, K. Isolation and characterization of acidocin A and cloning of the bacteriocin gene from Lactobacillus acidophilus. Appl. Environ. Microbiol. 1995, 61, 1061–1067. [Google Scholar]
- Tomita, H.; Fujimoto, S.; Tanimoto, K.; Ike, Y. Cloning and genetic organization of the bacteriocin 31 determinant encoded on the Enterococcus faecalis pheromone-responsive conjugative plasmid pYI17. J. Bacteriol. 1996, 178, 3585–3593. [Google Scholar] [CrossRef] [Green Version]
- Quadri, L.; Sailer, M.; Roy, K.L.; Vederas, J.C.; Stiles, M.E. Chemical and genetic characterization of bacteriocins produced by Carnobacterium piscicola LV17B. J. Biol. Chem. 1994, 269, 12204–12211. [Google Scholar] [CrossRef]
- De Kwaadsteniet, M.; Fraser, T.; Van Reenen, C.; Dicks, L. Bacteriocin T8, a novel class IIa sec-dependent bacteriocin produced by Enterococcus faecium T8, isolated from vaginal secretions of children infected with human immunodeficiency virus. Appl. Environ. Microbiol. 2006, 72, 4761–4766. [Google Scholar] [CrossRef] [Green Version]
- Quadri, L.; Sailer, M.; Terebiznik, M.R.; Roy, K.L.; Vederas, J.C.; Stiles, M.E. Characterization of the protein conferring immunity to the antimicrobial peptide carnobacteriocin B2 and expression of carnobacteriocins B2 and BM1. J. Bacteriol. 1995, 177, 1144–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tichaczek, P.S.; Nissen-Meyer, J.; Nes, I.F.; Vogel, R.F.; Hammes, W.P. Characterization of the bacteriocins curvacin A from Lactobacillus curvatus LTH1174 and sakacin P from L. sake LTH673. Syst. Appl. Microbiol. 1992, 15, 460–468. [Google Scholar] [CrossRef]
- Metivier, A.; Pilet, M.-F.; Dousset, X.; Sorokine, O.; Anglade, P.; Zagorec, M.; Piard, J.-C.; Marlon, D.; Cenatiempo, Y.; Fremaux, C. Divercin V41, a new bacteriocin with two disulphide bonds produced by Carnobacterium divergens V41: Primary structure and genomic organization. Microbiology 1998, 144, 2837–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aymerich, T.; Holo, H.; Håvarstein, L.S.; Hugas, M.; Garriga, M.; Nes, I.F. Biochemical and genetic characterization of enterocin A from Enterococcus faecium, a new antilisterial bacteriocin in the pediocin family of bacteriocins. Appl. Environ. Microbiol. 1996, 62, 1676–1682. [Google Scholar] [CrossRef] [Green Version]
- Barraza, D.E.; Ríos Colombo, N.S.; Galván, A.E.; Acuña, L.; Minahk, C.J.; Bellomio, A.; Chalón, M.C. New insights into enterocin CRL35: Mechanism of action and immunity revealed by heterologous expression in Escherichia coli. Mol. Microbiol. 2017, 105, 922–933. [Google Scholar] [CrossRef] [Green Version]
- Arbulu, S.; Lohans, C.T.; van Belkum, M.J.; Cintas, L.M.; Herranz, C.; Vederas, J.C.; Hernandez, P.E. Solution structure of enterocin HF, an antilisterial bacteriocin produced by Enterococcus faecium M3K31. J. Agric. Food Chem. 2015, 63, 10689–10695. [Google Scholar] [CrossRef]
- Cintas, L.M.; Casaus, P.; Håvarstein, L.S.; Hernandez, P.E.; Nes, I.F. Biochemical and genetic characterization of enterocin P, a novel sec-dependent bacteriocin from Enterococcus faecium P13 with a broad antimicrobial spectrum. Appl. Environ. Microbiol. 1997, 63, 4321–4330. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, T.; Kaminaka, K.; Shima, J.; Kawamoto, S.; Mori, K.; Choi, S.H.; Doi, K.; Ohmomo, S.; Ogata, S. Isolation and characterization of Enterocin SE-K4 produced by thermophilic enterococci, Enterococcus faecalis K-4. Biosci. Biotechnol. Biochem. 2001, 65, 247–253. [Google Scholar] [CrossRef]
- van Belkum, M.J.; Stiles, M.E. Molecular characterization of genes involved in the production of the bacteriocin leucocin A from Leuconostoc gelidum. Appl. Environ. Microbiol. 1995, 61, 3573–3579. [Google Scholar] [CrossRef] [Green Version]
- Héchard, Y.; Dérijard, B.; Letellier, F.; Cenatiempo, Y. Characterization and purification of mesentericin Y105, an anti-Listeria bacteriocin from Leuconostoc mesenteroides. Microbiology 1992, 138, 2725–2731. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, S.; Shima, J.; Sato, R.; Eguchi, T.; Ohmomo, S.; Shibato, J.; Horikoshi, N.; Takeshita, K.; Sameshima, T. Biochemical and genetic characterization of mundticin KS, an antilisterial peptide produced by Enterococcus mundtii NFRI 7393. Appl. Environ. Microbiol. 2002, 68, 3830–3840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, J.T.; Chopko, A.L.; Van Wassenaar, P.D. Purification and primary structure of pediocin PA-1 produced by Pediococcus acidilactici PAC-1.0. Arch. Biochem. Biophys. 1992, 295, 5–12. [Google Scholar] [CrossRef]
- Jack, R.W.; Wan, J.; Gordon, J.; Harmark, K.; Davidson, B.E.; Hillier, A.J.; Wettenhall, R.; Hickey, M.W.; Coventry, M.J. Characterization of the chemical and antimicrobial properties of piscicolin 126, a bacteriocin produced by Carnobacterium piscicola JG126. Appl. Environ. Microbiol. 1996, 62, 2897–2903. [Google Scholar] [PubMed]
- Reenen, V. Isolation, purification and partial characterization of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum. J. Appl. Microbiol. 1998, 84, 1131–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holck, A.; Axelsson, L.; Birkeland, S.-E.; Aukrust, T.; Blom, H. Purification and amino acid sequence of sakacin A, a bacteriocin from Lactobacillus sake Lb706. Microbiology 1992, 138, 2715–2720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tichaczek, P.S.; Vogel, R.F.; Hammes, W.P. Cloning and sequencing of sakP encoding sakacin P, the bacteriocin produced by Lactobacillus sake LTH 673. Microbiology 1994, 140, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; Fremaux, C.; Cenatiempo, Y.; Berjeaud, J. Sakacin G, a new type of antilisterial bacteriocin. Appl. Environ. Microbiol. 2002, 68, 6416–6420. [Google Scholar] [CrossRef] [Green Version]
- Svetoch, E.A.; Eruslanov, B.V.; Levchuk, V.P.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Stepanshin, J.; Dyatlov, I.; Seal, B.S.; Stern, N.J. Isolation of Lactobacillus salivarius 1077 (NRRL B-50053) and characterization of its bacteriocin, including the antimicrobial activity spectrum. Appl. Environ. Microbiol. 2011, 77, 2749–2754. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, A.L.; Montville, T.J. Purification of the bacteriocin bavaricin MN and characterization of its mode of action against Listeria monocytogenes Scott A cells and lipid vesicles. Appl. Environ. Microbiol. 1996, 62, 4529–4535. [Google Scholar] [CrossRef] [Green Version]
- Larsen, A.G.; Vogensen, F.; Josephsen, J. Antimicrobial activity of lactic acid bacteria isolated from sour doughs: Purification and characterization of bavaricin A, a bacteriocin produced by Lactobacillus bavaricus MI401. J. Appl. Bacteriol. 1993, 75, 113–122. [Google Scholar] [CrossRef]
- Yildirim, Z.; Winters, D.; Johnson, M. Purification, amino acid sequence and mode of action of bifidocin B produced by Bifidobacterium bifidum NCFB 1454. J. Appl. Microbiol. 1999, 86, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferchichi, M.; Frère, J.; Mabrouk, K.; Manai, M. Lactococcin MMFII, a novel class IIa bacteriocin produced by Lactococcus lactis MMFII, isolated from a Tunisian dairy product. FEMS Microbiol. Lett. 2001, 205, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Fimland, G.; Sletten, K.; Nissen-Meyer, J. The complete amino acid sequence of the pediocin-like antimicrobial peptide leucocin C. Biochem. Biophys. Res. Commun. 2002, 295, 826–827. [Google Scholar] [CrossRef]
- Bhugaloo-Vial, P.; Dousset, X.; Metivier, A.; Sorokine, O.; Anglade, P.; Boyaval, P.; Marion, D. Purification and amino acid sequences of piscicocins V1a and V1b, two class IIa bacteriocins secreted by Carnobacterium piscicola V1 that display significantly different levels of specific inhibitory activity. Appl. Environ. Microbiol. 1996, 62, 4410–4416. [Google Scholar] [CrossRef] [Green Version]
- Atrih, A.; Rekhif, N.; Moir, A.; Lebrihi, A.; Lefebvre, G. Mode of action, purification and amino acid sequence of plantaricin C19, an anti-Listeria bacteriocin produced by Lactobacillus plantarum C19. Int. J. Food Microbiol. 2001, 68, 93–104. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Y.; Xie, Q.; Zhang, Y.; Hu, J.; Li, P. Purification and characterization of plantaricin LPL-1, a novel class IIa bacteriocin produced by Lactobacillus plantarum LPL-1 isolated from fermented fish. Front. Microbiol. 2018, 9, 2276. [Google Scholar] [CrossRef]
- Flynn, S.; Van Sinderen, D.; Thornton, G.M.; Holo, H.; Nes, I.F.; Collins, J.K. Characterization of the genetic locus responsible for the production of ABP-118, a novel bacteriocin produced by the probiotic bacterium Lactobacillus salivarius subsp. salivarius UCC118 The GenBank accession number for the sequence reported in this paper is AF408405. Microbiology 2002, 148, 973–984. [Google Scholar]
- Neysens, P.; De Vuyst, L. Carbon dioxide stimulates the production of amylovorin L by Lactobacillus amylovorus DCE 471, while enhanced aeration causes biphasic kinetics of growth and bacteriocin production. Int. J. Food Microbiol. 2005, 105, 191–202. [Google Scholar] [CrossRef]
- Majhenič, A.Č.; Venema, K.; Allison, G.; Matijašić, B.B.; Rogelj, I.; Klaenhammer, T. DNA analysis of the genes encoding acidocin LF221 A and acidocin LF221 B, two bacteriocins produced by Lactobacillus gasseri LF221. Appl. Microbiol. Biotechnol. 2004, 63, 705–714. [Google Scholar] [CrossRef]
- Noda, M.; Miyauchi, R.; Danshiitsoodol, N.; Matoba, Y.; Kumagai, T.; Sugiyama, M. Expression of genes involved in bacteriocin production and self-resistance in Lactobacillus brevis 174A is mediated by two regulatory proteins. Appl. Environ. Microbiol. 2018, 84, e02707–e02717. [Google Scholar] [CrossRef] [Green Version]
- Acedo, J.Z.; Towle, K.M.; Lohans, C.T.; Miskolzie, M.; McKay, R.T.; Doerksen, T.A.; Vederas, J.C.; Martin-Visscher, L.A. Identification and three-dimensional structure of carnobacteriocin XY, a class IIb bacteriocin produced by Carnobacteria. FEBS Lett. 2017, 591, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-B.; Malaphan, W.; Zendo, T.; Nakayama, J.; Sonomoto, K. Enterocin X, a novel two-peptide bacteriocin from Enterococcus faecium KU-B5, has an antibacterial spectrum entirely different from those of its component peptides. Appl. Environ. Microbiol. 2010, 76, 4542–4545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, C.M.; Grube, A.; Herrmann, A.; Abriouel, H.; Stärke, J.; Lombardi, A.; Tauscher, B.; Holzapfel, W.H. Biochemical and genetic characterization of the two-peptide bacteriocin enterocin 1071 produced by Enterococcus faecalis FAIR-E 309. Appl. Environ. Microbiol. 2002, 68, 2550–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasuga, G.; Tanaka, M.; Harada, Y.; Nagashima, H.; Yamato, T.; Wakimoto, A.; Arakawa, K.; Kawai, Y.; Kok, J.; Masuda, T. Homologous expression and characterization of gassericin T and gassericin S, a novel class IIb bacteriocin produced by Lactobacillus gasseri LA327. Appl. Environ. Microbiol. 2019, 85, e02815–e02818. [Google Scholar] [CrossRef] [Green Version]
- Qi, F.; Chen, P.; Caufield, P.W. The group I strain of Streptococcus mutans, UA140, produces both the lantibiotic mutacin I and a nonlantibiotic bacteriocin, mutacin IV. Appl. Environ. Microbiol. 2001, 67, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Moll, G.; Ubbink-Kok, T.; Hildeng-Hauge, H.; Nissen-Meyer, J.; Nes, I.F.; Konings, W.N.; Driessen, A. Lactococcin G is a potassium ion-conducting, two-component bacteriocin. J. Bacteriol. 1996, 178, 600–605. [Google Scholar] [CrossRef] [Green Version]
- Zendo, T.; Koga, S.; Shigeri, Y.; Nakayama, J.; Sonomoto, K. Lactococcin Q, a novel two-peptide bacteriocin produced by Lactococcus lactis QU 4. Appl. Environ. Microbiol. 2006, 72, 3383–3389. [Google Scholar] [CrossRef] [Green Version]
- Castellano, P.; Farias, M.E.; Holzapfel, W.; Vignolo, G. Sensitivity variations of Listeria strains to the bacteriocins, lactocin 705, enterocin CRL35 and nisin. Biotechnol. Lett. 2001, 23, 605–608. [Google Scholar] [CrossRef]
- Maldonado, A.; Ruiz-Barba, J.L.; Jiménez-Díaz, R. Production of plantaricin NC8 by Lactobacillus plantarum NC8 is induced in the presence of different types of gram-positive bacteria. Arch. Microbiol. 2004, 181, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Stephens, S.K.; Floriano, B.; Cathcart, D.P.; Bayley, S.A.; Witt, V.F.; Jiménez-Díaz, R.; Warner, P.J.; Ruiz-Barba, J.L. Molecular analysis of the locus responsible for production of plantaricin S, a two-peptide bacteriocin produced by Lactobacillus plantarum LPCO10. Appl. Environ. Microbiol. 1998, 64, 1871–1877. [Google Scholar] [CrossRef] [Green Version]
- Ekblad, B.; Kyriakou, P.K.; Oppegård, C.; Nissen-Meyer, J.; Kaznessis, Y.N.; Kristiansen, P.E. Structure–function analysis of the two-peptide bacteriocin plantaricin EF. Biochemistry 2016, 55, 5106–5116. [Google Scholar] [CrossRef] [PubMed]
- Rogne, P.; Haugen, C.; Fimland, G.; Nissen-Meyer, J.; Kristiansen, P.E. Three-dimensional structure of the two-peptide bacteriocin plantaricin JK. Peptides 2009, 30, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.; Hayes, M.; O’Connor, P.; Gardiner, G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P.; Hill, C. Salivaricin P, one of a family of two-component antilisterial bacteriocins produced by intestinal isolates of Lactobacillus salivarius. Appl. Environ. Microbiol. 2007, 73, 3719–3723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciset, O.; Jeronimus-Stratingh, M.C.; Mollet, B.; Poolman, B. Thermophilin 13, a nontypical antilisterial poration complex bacteriocin, that functions without a receptor. J. Biol. Chem. 1997, 272, 14277–14284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.-C.; Liu, C.-F.; Lin, J.-F.; Li, A.-C.; Lo, T.-C.; Lin, T.-H. Characterization of putative class II bacteriocins identified from a non-bacteriocin-producing strain Lactobacillus casei ATCC 334. Appl. Environ. Microbiol. 2013, 97, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Martin-Visscher, L.A.; van Belkum, M.J.; Garneau-Tsodikova, S.; Whittal, R.M.; Zheng, J.; McMullen, L.M.; Vederas, J.C. Isolation and characterization of carnocyclin A, a novel circular bacteriocin produced by Carnobacterium maltaromaticum UAL307. Appl. Environ. Microbiol. 2008, 74, 4756–4763. [Google Scholar] [CrossRef] [Green Version]
- Himeno, K.; Rosengren, K.J.; Inoue, T.; Perez, R.H.; Colgrave, M.L.; Lee, H.S.; Chan, L.Y.; Henriques, S.n.T.; Fujita, K.; Ishibashi, N. Identification, characterization, and three-dimensional structure of the novel circular bacteriocin, enterocin NKR-5-3B, from Enterococcus faecium. Biochemistry 2015, 54, 4863–4876. [Google Scholar] [CrossRef]
- Grande Burgos, M.J.; Pulido, R.P.; Del Carmen López Aguayo, M.; Gálvez, A.; Lucas, R. The cyclic antibacterial peptide enterocin AS-48: Isolation, mode of action, and possible food applications. Int. J. Mol. Sci. 2014, 15, 22706–22727. [Google Scholar] [CrossRef] [Green Version]
- Borrero, J.; Brede, D.A.; Skaugen, M.; Diep, D.B.; Herranz, C.; Nes, I.F.; Cintas, L.M.; Hernández, P.E. Characterization of garvicin ML, a novel circular bacteriocin produced by Lactococcus garvieae DCC43, isolated from mallard ducks (Anas platyrhynchos). Appl. Environ. Microbiol. 2011, 77, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Masuda, Y.; Ono, H.; Kitagawa, H.; Ito, H.; Mu, F.; Sawa, N.; Zendo, T.; Sonomoto, K. Identification and characterization of leucocyclicin Q, a novel cyclic bacteriocin produced by Leuconostoc mesenteroides TK41401. Appl. Environ. Microbiol. 2011, 77, 8164–8170. [Google Scholar] [CrossRef] [Green Version]
- Sawa, N.; Zendo, T.; Kiyofuji, J.; Fujita, K.; Himeno, K.; Nakayama, J.; Sonomoto, K. Identification and characterization of lactocyclicin Q, a novel cyclic bacteriocin produced by Lactococcus sp. strain QU 12. Appl. Environ. Microbiol. 2009, 75, 1552–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrero, J.; Kelly, E.; O’Connor, P.M.; Kelleher, P.; Scully, C.; Cotter, P.D.; Mahony, J.; van Sinderen, D. Plantaricyclin A, a novel circular bacteriocin produced by Lactobacillus plantarum NI326: Purification, characterization, and heterologous production. Appl. Environ. Microbiol. 2018, 84, e01801–e01817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golneshin, A.; Gor, M.-C.; Williamson, N.; Vezina, B.; Van, T.T.H.; May, B.K.; Smith, A.T. Discovery and characterisation of circular bacteriocin plantacyclin B21AG from Lactiplantibacillus plantarum B21. Heliyon 2020, 6, e04715. [Google Scholar] [CrossRef] [PubMed]
- Wirawan, R.E.; Swanson, K.M.; Kleffmann, T.; Jack, R.W.; Tagg, J.R. Uberolysin: A novel cyclic bacteriocin produced by Streptococcus uberis. Microbiology 2007, 153, 1619–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acedo, J.Z.; van Belkum, M.J.; Lohans, C.T.; McKay, R.T.; Miskolzie, M.; Vederas, J.C. Solution structure of acidocin B, a circular bacteriocin produced by Lactobacillus acidophilus M46. Appl. Environ. Microbiol. 2015, 81, 2910–2918. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Kemperman, R.; Kok, J.; Saito, T. The circular bacteriocins gassericin A and circularin A. Curr. Protein Pept. Sci. 2004, 5, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Toba, T.; Samant, S.; Yoshioka, E.; Itoh, T. Reutericin 6, a new bacteriocin produced by Lactobacillus reuteri LA 6. Lett. Appl. Microbiol. 1991, 13, 281–286. [Google Scholar] [CrossRef]
- Liu, Y.-X.; Li, Z.-F.; Lv, Y.-J.; Dong, B.; Fan, Z.-C. Chlamydomonas reinhardtii-expressed multimer of Bacteriocin LS2 potently inhibits the growth of bacteria. Process Biochem. 2020, 95, 139–147. [Google Scholar] [CrossRef]
- Gálvez, A.; Valdivia, E.; Abriouel, H.; Camafeita, E.; Mendez, E.; Martínez-Bueno, M.; Maqueda, M. Isolation and characterization of enterocin EJ97, a bacteriocin produced by Enterococcus faecalis EJ97. Arch. Microbiol. 1998, 171, 59–65. [Google Scholar] [CrossRef]
- Criado, R.; Diep, D.B.; Aakra, A.; Gutiérrez, J.; Nes, I.F.; Hernández, P.E.; Cintas, L.M. Complete sequence of the enterocin Q-encoding plasmid pCIZ2 from the multiple bacteriocin producer Enterococcus faecium L50 and genetic characterization of enterocin Q production and immunity. Appl. Environ. Microbiol. 2006, 72, 6653–6666. [Google Scholar] [CrossRef] [Green Version]
- Uzelac, G.; Kojic, M.; Lozo, J.; Aleksandrzak-Piekarczyk, T.; Gabrielsen, C.; Kristensen, T.; Nes, I.F.; Diep, D.B.; Topisirovic, L. A Zn-dependent metallopeptidase is responsible for sensitivity to LsbB, a class II leaderless bacteriocin of Lactococcus lactis subsp. lactis BGMN1-5. J. Bacteriol. 2013, 195, 5614–5621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Ichimasa, S.; Zendo, T.; Koga, S.; Yoneyama, F.; Nakayama, J.; Sonomoto, K. Structural analysis and characterization of lacticin Q, a novel bacteriocin belonging to a new family of unmodified bacteriocins of gram-positive bacteria. Appl. Environ. Microbiol. 2007, 73, 2871–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwatani, S.; Zendo, T.; Yoneyama, F.; Nakayama, J.; Sonomoto, K. Characterization and structure analysis of a novel bacteriocin, lacticin Z, produced by Lactococcus lactis QU 14. Biosci. Biotechnol. Biochem. 2007, 71, 1984–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozo, J.; Mirkovic, N.; O’Connor, P.M.; Malesevic, M.; Miljkovic, M.; Polovic, N.; Jovcic, B.; Cotter, P.D.; Kojic, M. Lactolisterin BU, a novel class II broad-spectrum bacteriocin from Lactococcus lactis subsp. lactis bv. diacetylactis BGBU1-4. Appl. Environ. Microbiol. 2017, 83, e01519-17. [Google Scholar]
- Masuda, Y.; Zendo, T.; Sawa, N.; Perez, R.; Nakayama, J.; Sonomoto, K. Characterization and identification of weissellicin Y and weissellicin M, novel bacteriocins produced by Weissella hellenica QU 13. J. Appl. Microbiol. 2012, 112, 99–108. [Google Scholar] [CrossRef]
- Cintas, L.M.; Casaus, P.; Herranz, C.; Håvarstein, L.S.; Holo, H.; Hernández, P.E.; Nes, I.F. Biochemical and genetic evidence that Enterococcus faecium L50 produces enterocins L50A and L50B, the sec-dependent enterocin P, and a novel bacteriocin secreted without an N-terminal extension termed enterocin Q. J. Bacteriol. 2000, 182, 6806–6814. [Google Scholar] [CrossRef] [Green Version]
- Martín-Platero, A.M.; Valdivia, E.; Ruíz-Rodríguez, M.; Soler, J.J.; Martín-Vivaldi, M.; Maqueda, M.; Martínez-Bueno, M. Characterization of antimicrobial substances produced by Enterococcus faecalis MRR 10-3, isolated from the uropygial gland of the hoopoe (Upupa epops). Appl. Environ. Microbiol. 2006, 72, 4245–4249. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.; Diep, D.B.; Evensen, Ø.; Munang’andu, H.M. Garvicin KS, a Broad-Spectrum Bacteriocin Protects Zebrafish Larvae against Lactococcus garvieae Infection. Int. J. Mol. Sci. 2022, 23, 2833. [Google Scholar] [CrossRef]
- Whitford, M.; McPherson, M.; Forster, R.; Teather, R. Identification of bacteriocin-like inhibitors from rumen Streptococcus spp. and isolation and characterization of bovicin 255. Appl. Environ. Microbiol. 2001, 67, 569–574. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, E.; Wagner, C.; Marchioni, E.; Aoude-Werner, D.; Ennahar, S. Enterocin 96, a novel class II bacteriocin produced by Enterococcus faecalis WHE 96, isolated from Munster cheese. Appl. Environ. Microbiol. 2009, 75, 4273–4276. [Google Scholar] [CrossRef] [Green Version]
- Tosukhowong, A.; Zendo, T.; Visessanguan, W.; Roytrakul, S.; Pumpuang, L.; Jaresitthikunchai, J.; Sonomoto, K. Garvieacin Q, a novel class II bacteriocin from Lactococcus garvieae BCC 43578. Appl. Environ. Microbiol. 2012, 78, 1619–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado-Barragán, A.; Cárdenas, N.; Martínez, B.; Ruiz-Barba, J.L.; Fernández-Garayzábal, J.F.; Rodríguez, J.M.; Gibello, A. Garvicin A, a novel class IId bacteriocin from Lactococcus garvieae that inhibits septum formation in L. garvieae strains. Appl. Environ. Microbiol. 2013, 79, 4336–4346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, B.; Rodríguez, A.; Suárez, J.E. Lactococcin 972, a bacteriocin that inhibits septum formation in lactococci. Microbiology 2000, 146, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Holo, H.; Nilssen, Ø.; Nes, I. Lactococcin A, a new bacteriocin from Lactococcus lactis subsp. cremoris: Isolation and characterization of the protein and its gene. J. Bacteriol. 1991, 173, 3879–3887. [Google Scholar]
- Venema, K.; Abee, T.; Haandrikman, A.J.; Leenhouts, K.J.; Kok, J.; Konings, W.N.; Venema, G. Mode of action of lactococcin B, a thiol-activated bacteriocin from Lactococcus lactis. Appl. Environ. Microbiol. 1993, 59, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Casaus, P.; Nilsen, T.; Cintas, L.M.; Nes, I.F.; Hernández, P.E.; Holo, H. Enterocin B, a new bacteriocin from Enterococcus faecium T136 which can act synergistically with enterocin A. Microbiology 1997, 143, 2287–2294. [Google Scholar] [CrossRef] [Green Version]
- Worobo, R.W.; Henkel, T.; Sailer, M.; Roy, K.L.; Vederas, J.C.; Stiles, M.E. Characteristics and genetic determinant of a hydrophobic peptide bacteriocin, carnobacteriocin A, produced by Carnobacterium piscicola LV17A. Microbiology 1994, 140, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Nes, I.F.; Brede, D.A.; Diep, D.B. Chapter 16-Class II Non-Lantibiotic Bacteriocins. In Handbook of Biologically Active Peptides; Academic Press: Cambridge, MA, USA, 2013; pp. 85–92. [Google Scholar]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Factories 2014, 13, S3. [Google Scholar] [CrossRef] [Green Version]
- Nes, I.F.; Johnsborg, O. Exploration of antimicrobial potential in LAB by genomics. Curr. Opin. Biotechnol. 2004, 15, 100–104. [Google Scholar] [CrossRef]
- Aucher, W.; Lacombe, C.; Héquet, A.; Frère, J.; Berjeaud, J.-M. Influence of Amino Acid Substitutions in the Leader Peptide on Maturation and Secretion of Mesentericin Y105 by Leuconostoc mesenteroides. J. Bacteriol. 2005, 187, 2218–2223. [Google Scholar] [CrossRef] [Green Version]
- Letzel, A.-C.; Pidot, S.J.; Hertweck, C. Genome mining for ribosomally synthesized and post-translationally modified peptides (RiPPs) in anaerobic bacteria. BMC Genom. 2014, 15, 983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, A.; Nasi, A.; Ferrocino, I.; Di Monaco, R.; Mauriello, G.; Villani, F.; Ercolini, D. Spoilage-related activity of Carnobacterium maltaromaticum strains in air-stored and vacuum-packed meat. Appl. Environ. Microbiol. 2011, 77, 7382–7393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray Mohapatra, A.; Jeevaratnam, K. Inhibiting bacterial colonization on catheters: Antibacterial and antibiofilm activities of bacteriocins from Lactobacillus plantarum SJ33. J. Glob. Antimicrob. Resist. 2019, 19, 85–92. [Google Scholar] [CrossRef]
- Khouiti, Z.; Simon, J.-P. Carnocin KZ213 produced by Carnobacterium piscicola 213 is adsorbed onto cells during growth. Its biosynthesis is regulated by temperature, pH and medium composition. J. Ind. Microbiol. Biotechnol. 2004, 31, 5–10. [Google Scholar] [CrossRef]
- Koné, A.P.; Zea, J.M.V.; Gagné, D.; Cinq-Mars, D.; Guay, F.; Saucier, L. Application of Carnobacterium maltaromaticum as a feed additive for weaned rabbits to improve meat microbial quality and safety. Meat Sci. 2018, 135, 174–188. [Google Scholar] [CrossRef]
- Moreno, M.F.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef]
- Javed, A.; Masud, T.; ul Ain, Q.; Imran, M.; Maqsood, S. Enterocins of Enterococcus faecium, emerging natural food preservatives. Ann. Microbiol. 2011, 61, 699–708. [Google Scholar] [CrossRef]
- Sun, Z.; Harris, H.M.; McCann, A.; Guo, C.; Argimon, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef]
- Hill, D.; Sugrue, I.; Tobin, C.; Hill, C.; Stanton, C.; Ross, R.P. The Lactobacillus casei Group: History and Health Related Applications. Front. Microbiol. 2018, 9, 2107. [Google Scholar] [CrossRef] [Green Version]
- Yerlikaya, O. Probiotic potential and biochemical and technological properties of Lactococcus lactis ssp. lactis strains isolated from raw milk and kefir grains. J. Dairy Sci. 2019, 102, 124–134. [Google Scholar]
- Frantzen, C.A.; Kot, W.; Pedersen, T.B.; Ardö, Y.M.; Broadbent, J.R.; Neve, H.; Hansen, L.H.; Dal Bello, F.; Østlie, H.M.; Kleppen, H.P.; et al. Genomic Characterization of Dairy Associated Leuconostoc Species and Diversity of Leuconostocs in Undefined Mixed Mesophilic Starter Cultures. Front. Microbiol. 2017, 8, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkroth, K.J.; Vandamme, P.; Korkeala, H.J. Identification and characterization of Leuconostoc carnosum, associated with production and spoilage of vacuum-packaged, sliced, cooked ham. Appl. Environ. Microbiol. 1998, 64, 3313–3319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Kim, E.; Lee, Y.; Han, S.K.; Kim, C.G.; Choo, D.W.; Kim, Y.R.; Kim, H.Y. Rapid and accurate identification of species of the genus Pediococcus isolated from Korean fermented foods by matrix-assisted laser desorption/ionization time-of-flight MS with local database extension. Int. J. Syst. Evol. Microbiol. 2017, 67, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M.; Anastasiadou, S. Pediocins: The bacteriocins of Pediococci. Sources, production, properties and applications. Microb. Cell Factories 2009, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugster, E.; Fuchsmann, P.; Schlichtherle-Cerny, H.; Bütikofer, U.; Irmler, S. Formation of alanine, α-aminobutyrate, acetate, and 2-butanol during cheese ripening by Pediococcus acidilactici FAM18098. Int. Dairy J. 2019, 96, 21–28. [Google Scholar] [CrossRef]
- Cui, Y.; Xu, T.; Qu, X.; Hu, T.; Jiang, X.; Zhao, C. New Insights into Various Production Characteristics of Streptococcus thermophilus Strains. Int. J. Mol. Sci. 2016, 17, 1701. [Google Scholar] [CrossRef] [PubMed]
- Nes, I.F.; Diep, D.B.; Holo, H. Bacteriocin diversity in Streptococcus and Enterococcus. J. Bacteriol. 2007, 189, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Kot, W.; Neve, H.; Heller, K.J.; Vogensen, F.K. Bacteriophages of Leuconostoc, Oenococcus, and Weissella. Front. Microbiol. 2014, 5, 186. [Google Scholar] [CrossRef] [Green Version]
- Oppegaård, C.; Fimland, G.; Anonsen, J.H.; Nissen-Meyer, J. The pediocin PA-1 accessory protein ensures correct disulfide bond formation in the antimicrobial peptide pediocin PA-1. Biochemistry 2015, 54, 2967–2974. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, C.; Wang, Y.; Shi, J.; Zhang, L.; Ding, Z.; Qu, X.; Cui, H. Class IIa bacteriocins: Diversity and new developments. Int. J. Mol. Sci. 2012, 13, 16668–16707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, L.; Nielsen, M.K.; Ng, Y.; Gram, L. Role of acetate in production of an autoinducible class IIa bacteriocin in Carnobacterium piscicola A9b. Appl. Environ. Microbiol. 2002, 68, 2251–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fimland, G.; Eijsink, V.G.H.; Nissen-Meyer, J. Comparative studies of immunity proteins of pediocin-like bacteriocins. Microbiology 2002, 148, 3661–3670. [Google Scholar] [CrossRef] [Green Version]
- Martin-Visscher, L.A.; Sprules, T.; Gursky, L.J.; Vederas, J.C. Nuclear Magnetic Resonance Solution Structure of PisI, a Group B Immunity Protein that Provides Protection Against the Type IIa Bacteriocin Piscicolin 126, PisA. Biochemistry 2008, 47, 6427–6436. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, G.; Wang, C.; Ren, F.; Hao, Y. Both IIC and IID Components of Mannose Phosphotransferase System Are Involved in the Specific Recognition between Immunity Protein PedB and Bacteriocin-Receptor Complex. PLoS ONE 2016, 11, e0164973. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, A.; Eijsink, V.G.; Van Sinderen, D. Functional characterization of a composite bacteriocin locus from malt isolate Lactobacillus sakei 5. Appl. Environ. Microbiol. 2003, 69, 7194–7203. [Google Scholar] [CrossRef] [Green Version]
- Diep, D.B.; Havarstein, L.S.; Nes, I.F. Characterization of the locus responsible for the bacteriocin production in Lactobacillus plantarum C11. J. Bacteriol. 1996, 178, 4472–4483. [Google Scholar] [CrossRef] [Green Version]
- Diep, D.B.; Johnsborg, O.; Risøen, P.A.; Nes, I.F. Evidence for dual functionality of the operon plnABCD in the regulation of bacteriocin production in Lactobacillus plantarum. Mol. Microbiol. 2001, 41, 633–644. [Google Scholar] [CrossRef]
- Straume, D.; Kjos, M.; Nes, I.F.; Diep, D.B. Quorum-sensing based bacteriocin production is down-regulated by N-terminally truncated species of gene activators. Mol. Genet. Genom. 2007, 278, 283–293. [Google Scholar] [CrossRef]
- Maldonado, A.; Jiménez-Díaz, R.; Ruiz-Barba, J.L. Induction of plantaricin production in Lactobacillus plantarum NC8 after coculture with specific gram-positive bacteria is mediated by an autoinduction mechanism. J. Bacteriol. 2004, 186, 1556–1564. [Google Scholar] [CrossRef] [Green Version]
- Kjos, M.; Borrero, J.; Opsata, M.; Birri, D.J.; Holo, H.; Cintas, L.M.; Snipen, L.; Hernandez, P.E.; Nes, I.F.; Diep, D.B. Target recognition, resistance, immunity and genome mining of class II bacteriocins from Gram-positive bacteria. Microbiology 2011, 157, 3256–3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppegård, C.; Emanuelsen, L.; Thorbek, L.; Fimland, G.; Nissen-Meyer, J. The lactococcin G immunity protein recognizes specific regions in both peptides constituting the two-peptide bacteriocin lactococcin G. Appl. Environ. Microbiol. 2010, 76, 1267–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, A.P.; van der Ende, S.R.; van Belkum, M.J.; Martin-Visscher, L.A. The membrane topology of immunity proteins for the two-peptide bacteriocins carnobacteriocin XY, lactococcin G, and lactococcin MN shows structural diversity. MicrobiologyOpen 2020, 9, e00957. [Google Scholar] [CrossRef] [PubMed]
- Cebrián, R.; Martínez-Bueno, M.; Valdivia, E.; Albert, A.; Maqueda, M.; Sánchez-Barrena, M.J. The bacteriocin AS-48 requires dimer dissociation followed by hydrophobic interactions with the membrane for antibacterial activity. J. Struct. Biol. 2015, 190, 162–172. [Google Scholar] [CrossRef]
- Sanchez-Barrena, M.J.; Martinez-Ripoll, M.; Galvez, A.; Valdivia, E.; Maqueda, M.; Cruz, V.; Albert, A. Structure of bacteriocin AS-48: From soluble state to membrane bound state. J. Mol. Biol. 2003, 334, 541–549. [Google Scholar] [CrossRef]
- González, C.; Langdon, G.M.; Bruix, M.; Gálvez, A.; Valdivia, E.; Maqueda, M.; Rico, M. Bacteriocin AS-48, a microbial cyclic polypeptide structurally and functionally related to mammalian NK-lysin. Proc. Natl. Acad. Sci. USA 2000, 97, 11221–11226. [Google Scholar] [CrossRef] [Green Version]
- Cebrián, R.; Maqueda, M.; Neira, J.L.; Valdivia, E.; Martínez-Bueno, M.; Montalbán-López, M. Insights into the Functionality of the Putative Residues Involved in Enterocin AS-48 Maturation. Appl. Environ. Microbiol. 2010, 76, 7268–7276. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.H.; Sugino, H.; Ishibashi, N.; Zendo, T.; Wilaipun, P.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. Mutations near the cleavage site of enterocin NKR-5-3B prepeptide reveal new insights into its biosynthesis. Microbiology 2017, 163, 431–441. [Google Scholar] [CrossRef]
- van Belkum, M.J.; Martin-Visscher, L.A.; Vederas, J.C. Structure and genetics of circular bacteriocins. Trends Microbiol. 2011, 19, 411–418. [Google Scholar] [CrossRef]
- Kawai, Y.; Kusnadi, J.; Kemperman, R.; Kok, J.; Ito, Y.; Endo, M.; Arakawa, K.; Uchida, H.; Nishimura, J.; Kitazawa, H.; et al. DNA sequencing and homologous expression of a small peptide conferring immunity to gassericin A, a circular bacteriocin produced by Lactobacillus gasseri LA39. Appl. Environ. Microbiol. 2009, 75, 1324–1330. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Bueno, M.; Valdivia, E.; Gálvez, A.; Coyette, J.; Maqueda, M. Analysis of the gene cluster involved in production and immunity of the peptide antibiotic AS-48 in Enterococcus faecalis. Mol. Microbiol. 1998, 27, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Belkum, M.J.; Martin-Visscher, L.A.; Vederas, J.C. Cloning and Characterization of the Gene Cluster Involved in the Production of the Circular Bacteriocin Carnocyclin A. Probiotics Antimicrob. Proteins 2010, 2, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, C.; Brede, D.A.; Nes, I.F.; Diep, D.B. Circular bacteriocins: Biosynthesis and mode of action. Appl. Environ. Microbiol. 2014, 80, 6854–6862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acedo, J.Z.; van Belkum, M.J.; Lohans, C.T.; Towle, K.M.; Miskolzie, M.; Vederas, J.C. Nuclear Magnetic Resonance Solution Structures of Lacticin Q and Aureocin A53 Reveal a Structural Motif Conserved among Leaderless Bacteriocins with Broad-Spectrum Activity. Biochemistry 2016, 55, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Gajic, O.; Buist, G.; Kojic, M.; Topisirovic, L.; Kuipers, O.P.; Kok, J. Novel Mechanism of Bacteriocin Secretion and Immunity Carried Out by Lactococcal Multidrug Resistance Proteins. J. Biol. Chem. 2003, 278, 34291–34298. [Google Scholar] [CrossRef] [Green Version]
- Iwatani, S.; Horikiri, Y.; Zendo, T.; Nakayama, J.; Sonomoto, K. Bifunctional gene cluster lnqBCDEF mediates bacteriocin production and immunity with differential genetic requirements. Appl. Environ. Microbiol. 2013, 79, 2446–2449. [Google Scholar] [CrossRef] [Green Version]
- Ladjouzi, R.; Lucau-Danila, A.; Benachour, A.; Drider, D. A Leaderless Two-Peptide Bacteriocin, Enterocin DD14, Is Involved in Its Own Self-Immunity: Evidence and Insights. Front. Bioeng. Biotechnol. 2020, 8, 644. [Google Scholar] [CrossRef]
- Kristiansen, P.E.; Persson, C.; Fuochi, V.; Pedersen, A.; Karlsson, G.B.; Nissen-Meyer, J.; Oppegård, C. Nuclear Magnetic Resonance Structure and Mutational Analysis of the Lactococcin a Immunity Protein. Biochemistry 2016, 55, 6250–6257. [Google Scholar] [CrossRef]
- Wang, J.; Xu, H.; Liu, S.; Song, B.; Liu, H.; Li, F.; Deng, S.; Wang, G.; Zeng, H.; Zeng, X.; et al. Toyoncin, a novel leaderless bacteriocin that is produced by Bacillus toyonensis XIN-YC13 and specifically targets B. Cereus and Listeria monocytogenes. Appl. Environ. Microbiol. 2021, 87, e00185-21. [Google Scholar] [CrossRef]
- Iwatani, S.; Ishibashi, N.; Flores, F.P.; Zendo, T.; Nakayama, J.; Sonomoto, K. LnqR, a TetR-family transcriptional regulator, positively regulates lacticin Q production in Lactococcus lactis QU 5. FEMS Microbiol. Lett. 2016, 363, fnw200. [Google Scholar] [CrossRef] [Green Version]
- Criado, R.; Gutiérrez, J.; Martín, M.; Herranz, C.; Hernández, P.E.; Cintas, L.M. Immunochemical characterization of temperature-regulated production of enterocin L50 (EntL50A and EntL50B), enterocin P, and enterocin Q by Enterococcus faecium L50. Appl. Environ. Microbiol. 2006, 72, 7634–7643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, Y.; Perez, R.H.; Zendo, T.; Sonomoto, K. Nutrition-adaptive control of multiple-bacteriocin production by Weissella hellenica QU 13. J. Appl. Microbiol. 2016, 120, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telke, A.A.; Ovchinnikov, K.V.; Vuoristo, K.S.; Mathiesen, G.; Thorstensen, T.; Diep, D.B. Over 2000-Fold Increased Production of the Leaderless Bacteriocin Garvicin KS by Increasing Gene Dose and Optimization of Culture Conditions. Front. Microbiol. 2019, 10, 389. [Google Scholar] [CrossRef]
- Sidooski, T.; Brandelli, A.; Bertoli, S.L.; Souza, C.K.d.; Carvalho, L.F.d. Physical and nutritional conditions for optimized production of bacteriocins by lactic acid bacteria—A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2839–2849. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Bashokouh, F.; Ramakrishnan, N.R.; Mustafa, S.; Ariff, A.B. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria: A review. RSC Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Fahim, H.A.; Rouby, W.M.A.E.; El-Gendy, A.O.; Khairalla, A.S.; Naguib, I.A.; Farghali, A.A. Enhancement of the productivity of the potent bacteriocin avicin A and improvement of its stability using nanotechnology approaches. Sci. Rep. 2017, 7, 10604. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Bai, J.; Cai, Z.; Ouyang, F. Optimization of a cultural medium for bacteriocin production by Lactococcus lactis using response surface methodology. J. Biotechnol. 2002, 93, 27–34. [Google Scholar] [CrossRef]
- Avonts, L.; Van Uytven, E.; De Vuyst, L. Cell growth and bacteriocin production of probiotic Lactobacillus strains in different media. Int. Dairy J. 2004, 14, 947–955. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; West, S.A. The cost and benefit of quorum sensing-controlled bacteriocin production in Lactobacillus plantarum. J. Evol. Biol. 2020, 33, 101–111. [Google Scholar] [CrossRef]
- Man, L.-L.; Xiang, D.-J. LuxS-mediated quorum sensing system in Lactobacillus plantarum NMD-17 from koumiss: Induction of plantaricin MX in co-cultivation with certain lactic acid bacteria. Folia Microbiol. 2021, 66, 855–871. [Google Scholar] [CrossRef]
- Piazentin, A.C.M.; Mendonça, C.M.N.; Vallejo, M.; Mussatto, S.I.; de Souza Oliveira, R.P. Bacteriocin-like inhibitory substances production by Enterococcus faecium 135 in co-culture with Ligilactobacillus salivarius and Limosilactobacillus reuteri. Braz. J. Microbiol. 2022, 53, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Cortés, C.; Suarez, H.; Buitrago, G.; Nero, L.A.; Todorov, S.D. Enhanced Bacteriocin Production by Pediococcus pentosaceus 147 in Co-culture with Lactobacillus plantarum LE27 on Cheese Whey Broth. Front. Microbiol. 2018, 9, 2952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawan, R.; Abbasiliasi, S.; Tan, J.S.; Mustafa, S.; Halim, M.; Ariff, A.B. Influence of culture conditions and medium compositions on the production of bacteriocin-like inhibitory substances by Lactococcus lactis Gh1. Microorganisms 2020, 8, 1454. [Google Scholar] [CrossRef] [PubMed]
- Lajis, A.F.B. Biomanufacturing process for the production of bacteriocins from Bacillaceae family. Bioresour. Bioprocess. 2020, 7, 8. [Google Scholar] [CrossRef]
- Delgado, A.; Arroyo López, F.N.; Brito, D.; Peres, C.; Fevereiro, P.; Garrido-Fernández, A. Optimum bacteriocin production by Lactobacillus plantarum 17.2b requires absence of NaCl and apparently follows a mixed metabolite kinetics. J. Biotechnol. 2007, 130, 193–201. [Google Scholar]
- Parlindungan, E.; May, B.K.; Jones, O.A.H. Metabolic Insights into the Effects of Nutrient Stress on Lactobacillus plantarum B21. Front. Mol. Biosci. 2019, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neysens, P.; Messens, W.; De Vuyst, L. Effect of sodium chloride on growth and bacteriocin production by Lactobacillus amylovorus DCE 471. Int. J. Food Microbiol. 2003, 88, 29–39. [Google Scholar] [CrossRef]
- Abo-Amer, A.E. Optimization of bacteriocin production by Lactobacillus acidophilus AA11, a strain isolated from Egyptian cheese. Ann. Microbiol. 2011, 61, 445–452. [Google Scholar] [CrossRef]
- Radha, K.; Padmavathi, T. Statistical optimization of bacteriocin produced from Lactobacillus delbrueckii subsp bulgaricus isolated from yoghurt. Int. Food Res. J. 2017, 24, 803–809. [Google Scholar]
- Suganthi, V.; Mohanasrinivasan, V. Optimization studies for enhanced bacteriocin production by Pediococcus pentosaceus KC692718 using response surface methodology. J. Food Sci. Technol. 2015, 52, 3773–3783. [Google Scholar] [CrossRef]
- Wang, P.; Wang, T.; Ismael, M.; Wang, X.; Yi, Y.; Lü, X. Development of an electroporation method and expression patterns of bacteriocin-encoding genes in Companilactobacillus crustorum MN047. Food Biosci. 2021, 44, 101420. [Google Scholar] [CrossRef]
- Borrero, J.; Jiménez, J.J.; Gútiez, L.; Herranz, C.; Cintas, L.M.; Hernández, P.E. Use of the usp45 lactococcal secretion signal sequence to drive the secretion and functional expression of enterococcal bacteriocins in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2011, 89, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Arbulu, S.; Jiménez, J.J.; Gútiez, L.; Cintas, L.M.; Herranz, C.; Hernández, P.E. Cloning and Expression of Synthetic Genes Encoding the Broad Antimicrobial Spectrum Bacteriocins SRCAM 602, OR-7, E-760, and L-1077, by Recombinant Pichia pastoris. BioMed Res. Int. 2015, 2015, 767183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Wu, S.; Wang, X.; Gu, Q.; Li, P. Antimicrobial activity and preliminary mode of action of PlnEF expressed in Escherichia coli against Staphylococci. Protein Expr. Purif. 2018, 143, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Ma, J.; Chen, X.; Tan, Y.; Chen, P.; Zhu, X.; Liu, L. Expression and purification of recombinant Lactobacillus casei bacteriocin and analysis of its antibacterial activity. Cyta-J. Food 2020, 18, 301–308. [Google Scholar] [CrossRef]
- Acuña, L.; Corbalán, N.; Quintela-Baluja, M.; Barros-Velázquez, J.; Bellomio, A. Expression of the hybrid bacteriocin Ent35-MccV in Lactococcus lactis and its use for controlling Listeria monocytogenes and Escherichia coli in milk. Int. Dairy J. 2020, 104, 104650. [Google Scholar] [CrossRef]
- O’Connor, P.M.; Kuniyoshi, T.M.; Oliveira, R.P.; Hill, C.; Ross, R.P.; Cotter, P.D. Antimicrobials for food and feed; a bacteriocin perspective. Curr. Opin. Biotechnol. 2020, 61, 160–167. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Bacteriocins | Sequence | Producer Species | Strain | Reference | |
|---|---|---|---|---|---|
| ClassIIa | |||||
| Acidocin A | MISMISSHQKTLTDKELALISGGKTYYGTNGVHCTKRSLWGKVRLKNVIPGTLCRKQSLPIKQDLKILLGWATGAFGKTFH | Lactobacillus acidophilus | TK9201 | [19] | |
| Bacteriocin 31 | MKKKLVICGIIGIGFTALGTNVEAATATYYGNGLYCNKQKCWVDWNKASREIGKIIVNGWVQHGPWAPR | Enterococcus faecalis | YI717 | [20] | |
| Bacteriocin BM1 | MKSVKELNKKEMQQINGGAISYGNGVYCNKEKCWVNKAENKQAITGIVIGGWASSLAGMGH | Carnobacterium piscicola | LV17B | [21] | |
| Bacteriocin B2 | MNSVKELNVKEMKQLHGGVNYGNGVSCSKTKCSVNNGQAFQERYTAGINSFVSGVASGAGSIGRRP | Carnobacterium piscicola | LV17B | [21] | |
| Bacteriocin T8 | MKKKVLKHCVILGILGTCLAGIGTGIKVDAATYYGNGLYCNKEKCWVDWNQAKGEIGKIIVNGWVNHGPWAPRR | Enterococcus faecium | T8 | [22] | |
| Carnobacteriocin B2 | MNSVKELNVKEMKQLHGGVNYGNGVSCSKTKCSVNWGQAFQERYTAGINSFVSGVASGAGSIGRRP | Carnobacterium piscicola | LV17 | [23] | |
| Carnobacteriocin BM1 | MKSVKELNKKEMQQIIGGAISYGNGVYCNKEKCWVNKAENKQAITGIVIGGWASSLAGMGH | Carnobacterium piscicola | LV17B | [21] | |
| Curvacin A | MNNVKELSMTELQTITGGARSYGNGVYCNNKKCWVNRGEATQSIIGGMISGWASGLAGM | Latilactobacillus curvatus | LTH1174 | [24] | |
| Divercin V41 | MKNLKEGSYTAVNTDELKSINGGTKYYGNGVYCNSKKCWVDWGQASGCIGQTVVGGWLGGAIPGKC | Carnobacterium divergens | V41 | [25] | |
| Enterocin A | MKHLKILSIKETQLIYGGTTHSGKYYGNGVYCTKNKCTVDWAKATTCIAGMSIGGFLGGAIPGKC | Enterococcus faecium | CTCA92/T136 | [26] | |
| Enterocin CRL35 | MKKLTSKEMAQVVGGKYYGNGVSCNKKGCSVDWGKAIGIIGNNSAANLATGGAAGWKS | Enterococcus faecium | CRL 35 | [27] | |
| Enterocin HF | MEKLTVKEMSQVVGGKYYGNGVSCNKKGCSVDWGKAIGIIGNNAAANLTTGGKAGWKG | Enterococcus faecium | M3K31 | [28] | |
| Enterocin P | MRKKLFSLALIGIFGLVVTNFGTKVDAATRSYGNGVYCNNSKCWVNWGEAKENIAGIVISGWASGLAGMGH | Enterococcus faecium | P13 | [29] | |
| Enterocin SE-K4 | MKKKLVKGLVICGMIGIGFTALGTNVEAATYYGNGVYCNKQKCWVDWSRARSEIIDRGVKAYVNGFTKVLG | Enterococcus faecalis | K-4 | [30] | |
| Leucocin A | MMNMKPTESYEQLDNSALEQVVGGKYYGNGVHCTKSGCSVNWGEAFSAGVHRLANGGNGFW | Leuconostoc gelidum | UAL 187 | [31] | |
| Mesentericin Y105 | MTNMKSVEAYQQLDNQNLKKVVGGKYYGNGVHCTKSGCSVNWGEAASAGIHRLANGGNGFW | Leuconostoc mesenteroides | Y105 | [32] | |
| Mundticin KS | MKKLTAKEMSQVVGGKYYGNGVSCNKKGCSVDWGKAIGIIGNNSAANLATGGAAGWKS | Enterococcus mundtii | NFRI 7393 | [33] | |
| Pediocin PA-1 | MKKIEKLTEKEMANIIGGKYYGNGVTCGKHSCSVDWGKATTCIINNGAMAWATGGHQGNHKC | Pediococcus acidilactici | PAC 1.0 | [34] | |
| Piscicolin 126 | MKTVKELSVKEMQLTTGGKYYGNGVSCNKNGCTVDWSKAIGIIGNNAAANLTTGGAAGWNKG | Carnobacterium piscicola | JG126 | [35] | |
| Plantaricin 423 | MMKKIEKLTEKEMANIIGGKYYGNGVTCGKHSCSVNWGQAFSCSVSHLANFGHGKC | Lactiplantibacillus plantarum | 423 | [36] | |
| Sakacin A | MNNVKELSMTELQTITGGARSYGNGVYCNNKKCWVNRGEATQSIIGGMISGWASGLAGM | Latilactobacillus sakei | 706 | [37] | |
| Sakacin P | MEKFIELSLKEVTAITGGKYYGNGVHCGKHSCTVDWGTAIGNIGNNAAANWATGGNAGWNK | Latilactobacillus sakei | MI401 | [38] | |
| Sakacin G | MKNTRSLTIQEIKSITGGKYYGNGVSCNSHGCSVNWGQAWTCGVNHLANGGHGGVC | Latilactobacillus sakei | 2512 | [39] | |
| Bacteriocin L-1077 | TNYGNGVGVPDAIMAGIIKLIFIFNIRQGYNFGKKAT | Ligilactobacillus salivarius | 1077 | [40] | |
| Bavaricin MN | TKYYGNGVYCNSKKCWVDWGQAAGGIGQTVVXGWLGGAIPGK | Lactobacillus bavaricus | MN | [41] | |
| Bavaricin A | KYYGNGVHCGKHSCTVDWGTAIGNIGNNAAANXATGXNAGG | Latilactobacillus sakei | MI401 | [42] | |
| Bifidocin B | KYYGNGVTCGLHDCRVDRGKATCGIINNGGMWGDIG | Bifidobacterium bifidum | NCFB 1454 | [43] | |
| Lactococcin MMFII | TSYGNGVHCNKSKCWIDVSELETYKAGTVSNPKDILW | Lactococcus lactis | MMFII | [44] | |
| Leucocin C | KNYGNGVHCTKKGCSVDWGYAWTNIANNSVMNGLTGGNAGWHN | Leuconostoc mesenteroides | TA33a | [45] | |
| Mundticin | KYYGNGVSCNKKGCSVDWGKAIGIIGNNSAANLATGGAAGWSK | Enterococcus mundtii | ATO6 | [33] | |
| Piscicocin VIa | KYYGNGVSCNKNGCTVDWSKAIGIIGNNAAANLTTGGAAGWNKG | Carnobacterium piscicola | V1 | [46] | |
| Plantaricin C19 | KYYGNGLSCSKKGCTVNWGQAFSCGVNRVATAGHGK | Lactiplantibacillus plantarum | C19 | [47] | |
| plantaricin LPL-1 | VIADKYYGNGVSCGKHTCTVDWGEAFSCSVSHLANFGHGKC | Lactiplantibacillus plantarum | LPL-1 | [48] | |
| Class IIb | |||||
| ABP-118 | 118α | MMKEFTVLTECELAKVDGGKRGPNCVGNFLGGLFAGAAAGVPLGPAGIVGGANLGMVGGALTCL | Ligilactobacillus salivarius | UCC118 | [49] |
| 118β | MKNLDKRFTIMTEDNLASVNGGKNGYGGSGNRWVHCGAGIVGGALIGAIGGPWSAVAGGISGGFTSCR | ||||
| Amylovorin L | amyLa | MSKGEVLNEDELTAVVGGSKGKGRNNWAGNTIGIVSSAATGAALGSAICGPGCGFVGAHWGAVGWTAVASFSGAFGKIRK | Lactobacillus amylovorus | DCE 471 | [50] |
| amyLb | MKQLNSEQLQNIIGGNRWTNAYSAALGCAVPGVKYGKKLGGVWGAVIGGVGGAAVCGLAGYVRKG | ||||
| Acidocin LF221B | LF221B | MIEKVSKNELSRIYGGNNVNWGSVAGSCGKGAVMEIYFGNPILGCANGAATSLVLQTASGIYKNYQKKR | Lactobacillus gasseri | LF221 | [51] |
| LF221β | MALKTLEKHELRNVMGGNKWGNAVIGAATGATRGVSWCRGFGPWGMTACALGGAAIGGYLGYKSN | ||||
| Brevicin 174A | 174A-β | MEKFAVLSLSDLVDIQGGKKKKKYTGPNYRCMVKSGGGLVSGAIGGSPFGVGGIVGGGMAGLVGGAISCLNNK | Levilactobacillus brevis | 174A | [52] |
| 174A-γ | MYKELTVDELALIDGGKKKKKKVACTWGNAATAAASGAVKGILGGPTGALAGAIWGVSQCASNNLHGMH | ||||
| Carnobacteriocin XY | CbnX | MKSVKELNVKEMQQTIGGWGWKEVVQNGQTIFSAGQKLGNMVGKIVPLPFG | Carnobacteria | [53] | |
| CbnY | MNKEFKSLNEVEMKKINGGSAILAITLGIFATGYGMGVQKAINDRRKK | ||||
| Enterocin X | EntXα | MQNVKEVSVKEMKQIIGGSNDSLWYGVGQFMGKQANCITNHPVKHMIIPGYCLSKILG | Enterococcus faecium | KU-B5 | [54] |
| EntXβ | MKKYNELSKKELLQIQGGIAPIIVAGLGYLVKDAWDHSDQIISGFKKGWNGGRRK | ||||
| Enterocin 1071 | Entα | MKQYKVLNEKEMKKPIGGESVFSKIGNAVGPAAYWILKGLGNMSDVNQADRINRKKH | Enterococcus faecalis | FAIR-E 309 | [55] |
| Entβ | MKNIKNASNIKVIEDNELKAITGGGPGKWLPWLQPAYDFVTGLAKGIGKEGNKNKWKNV | ||||
| Gassericin S | GasA | MKVLNECQLQTVVGGKNWSVAKCGGTIGTNIAIGAWRGARAGSFFGQPVSVGAGALIGASAGAIGGSVQCVGWLAGGGR | Lactobacillus gasseri | LA327 | [56] |
| GasX | MIEKVSKNELSRIYGGNNVNWGSVAGSCGKGAVMGIYFGNPILGCANGAATSLVLQTTSGIYKNYQKKR | ||||
| Gassericin T | GatA | MKNFNTLSFETLANIVGGRNNWAANIGGVGGATVAGWALGNAVCGPACGFVGAHYVPIAWAGVTAATGGFGKIRK | Lactobacillus gasseri | LA327 | [56] |
| GatX | MALKTLEKHELRNVMGGNKWGNAVIGAATGATRGVSWCRGFGPWGMTACGLGGAAIGGYLGYKSN | ||||
| Mutacin IV | nlm A | MDTQAFEQFDVMDSQTLSTVEGGKVSGGEAVAAIGICATASAAIGGLAGATLVTPYCVGTWGLIRSH | Streptococcus mutans | UA159 | [57] |
| nlm B | MELNVNNYKSLTNDELSEVFGGDKQAADTFLSAVGGAASGFTYCASNGVWHPYILAGCAGVGAVGSVVFPH | ||||
| Lactococcin G | LcnGα | MKELSEKELRECVGGGTWDDIGQGIGRVAYWVGKAMGNMSDVNQASRINRKKKH | Lactococcus lactis | [58] | |
| LcnGβ | MKNNNNFFKGMEIIEDQELVSITGGKKWGWLAWVDPAYEFIKGFGKGAIKEGNKDKWKNI | ||||
| Lactococcin Q | LcnQα | MKELSEKELRECVGGSIWGDIGQGVGKAAYWVGKAMGNMSDVNQASRINRKKKH | Lactococcus lactis | QU4 | [59] |
| LcnQβ | MKNNNNNFFKDMEIIEDQELVSITGGKKWGWLAWVEPAGEFLKGFGKGAIKEGNKDKWKNI | ||||
| Lactocin 705 | 705α | MESNKLEKFANISNKDLNKITGGGFWGGLGYIAGRVGAAYGHAQASANNHHSPING | Lacticaseibacillus casei | CRL 705 | [60] |
| 705β | MDNLNKFKKLSDNKLQATIGGGMSGYIQGIPDFLKGYLHGISAANKHKKGRLGY | ||||
| Plantaricin NC8 | PlnC8α | MDKFEKISTSNLEKISGGDLTTKLWSSWGYYLGKKARWNLKHPYVQF | Lactiplantibacillus plantarum | NC8 | [61] |
| PlnC8β | MNNLNKFSTLGKSSLSQIEGGSVPTSVYTLGIKILWSAYKHRKTIEKSFNKGFYH | ||||
| Plantaricin S | Plsα | MNNALSFEQQFTDFSTLSDSELESVEGGRNKLAYNMGHYAGKATIFGLAAWALLA | Lactiplantibacillus plantarum | LPCO10 | [62] |
| Plsβ | MDKIIKFQGISDDQLNAVIGGKKKKQSWYAAAGDAIVSFGEGFLNAW | ||||
| Plantaricin EF | PlnE | MLQFEKLQYSRLPQKKLAKISGGFNRGGFNRGGYNFGKSVRHVVDAIGSVAGIRGILKSIR | Lactiplantibacillus plantarum | C11 | [63] |
| PlnF | MKKFLVLRDRELNAISGGVFHAYSARGVRNNYKSAVGPADWVISAVRGFIHG | ||||
| Plantaricin JK | PlnJ | MTVNKMIKDLDVVDAFAPISNNKLNGVVGGGAWKNFWSSLRKGFYDGEAGRAIRR | Lactiplantibacillus plantarum | C11 | [64] |
| PlnK | MKIKLTVLNEFEELTADAEKNISGGRRSRKNGIGYAIGYAFGAVERAVLGGSRDYNK | ||||
| Salivaricin P | SalPα | MMKEFTVLTECELAKVDGGKRGPNCVGNFLGGLFAGAAAGVPLGPAGIVGGANLGMVGGALTCL | Ligilactobacillus salivarius | DPC6005 | [65] |
| SalPβ | MKNLDKRFTIMTEDNLASVNGGKNGYGGSGNRWVHCGAGIVGGALIGAIGGPWSAVAGGISGGFASCH | ||||
| Thermophilin 13 | ThmA | MNTITICKFDVLDAELLSTVEGGYSGKDCLKDMGGYALAGAGSGALWGAPAGGVGALPGAFVGAHVGAIAGGFACMGGMIGNKFN | Streptococcus thermophiles | SPi13 | [66] |
| ThmB | MKQYNGFEVLHELDLANVTGGQINWGSVVGHCIGGAIIGGAFSGGAAAGVGCLVGSGKAIINGL | ||||
| Uncharacterized | LSEI_2392 | MYTMTNLKDKELSQITGGFAFGIPVAAILGFLASDAWSHADEIAGGATSGWSLADKSHSL | Lacticaseibacillus casei | ATCC 334 | [67] |
| LSEI_2393 | MQQFMTLDNSSLEKIAGGENGGLWSIIGFGLGFSARSVLTGSLFVPSRGPVIDLVKQLTPKN | ||||
| Uncharacterized | LSEI_2405 | MISKEVGITLKQHDLVLIQGGAKRRNKPSGCIVSTIGGAVAGAAGLNPFTTVAGAAIGLSLPRLQ | Lacticaseibacillus casei | ATCC 334 | [67] |
| LSEI_2406 | MSYNYRQIDDFQLSGVSGGKKKFDCAATFVTGITAGIGSGTITGLAGGPFGIIGGAVVGGNLGAVGSAIKCLGDGMQ | ||||
| Class IIc | |||||
| Carnocyclin-A | MLYELVAYGIAQGTAEKVVSLINAGLTVGSIISILGGVTVGLSGVFTAVKAAIAKQGIKKAIQL | Carnobacterium maltaromaticum | UAL307 | [68] | |
| Enterocin NKR-5-3B | MKKNLLLVLPIVGIVGLFVGAPMLTANLGISSYAAKKVIDIINTGSAVATIIALVTAVVGGGLITAGIVATAKSLIKKYGAKYAAAW | Enterococcus faecium | NKR-5-3 | [69] | |
| Enterocin AS-48 | MVKENKFSKIFILMALSFLGLALFSASLQFLPIAHMAKEFGIPAAVAGTVLNVVEAGGWVTTIVSILTAVGSGGLSLLAAAGRESIKAYLKKEIKKKGKRAVIAW | Enterococcus faecalis | S-48 | [70] | |
| Garvicin ML | MFDLVATGMAAGVAKTIVNAVSAGMDIATALSLFSGAFTAAGGIMALIKKYAQKKLWKQLIAA | Lactococcus garvieae | DCC43 | [71] | |
| Leucocyclicin Q | MFLVNQLGISKSLANTILGAIAVGNLASWLLALVPGPGWATKAALATAETIVKHEGKAAAIAW | Leuconostoc mesenteroides | TK41401 | [72] | |
| Lactocyclicin Q | MFLIDHLGAPRWAVDTILGAIAVGNLASWVLALVPGPGWAVKAGLATAAAIVKHQGKAAAAAW | Lactococcus sp. | QU 12 | [73] | |
| Plantaricyclin A | MLSAYRSKLGLNKFEVTVLMIISLFILLFATVNIVWIAKQFGVHLTTSLTQKALDLLSAGSSLGTVAAAVLGVTLPAWAVAAAGALGGTAA | Lactiplantibacillus plantarum | NI326 | [74] | |
| Plantacyclin B21AG | MLSAYRSRLGLNKFEVAILMIISLFILLFATVNIVWIARQFGVHLTTKLTQKALDLLSSGASLGTVAAVILGVTLPGWAVAAAGALGGTAA | Lactiplantibacillus plantarum | B21 | [75] | |
| Uberolysin | MDILLELAGYTGIASGTAKKVVDAIDKGAAAFVIISIISTVISAGALGAVSASADFIILTVKNYISRNLKAQAVIW | Streptococcus uberis | 42 | [76] | |
| Acidocin B | MVTKYGRNLGLSKVELFAIWAVLVVALLLATANIYWIADQFGIHLATGTARKLLDAVASGASLGTAFAAILGVTLPAWALAAAGALGATAA | Lactobacillus acidophilus | M46 | [77] | |
| Gassericin A | MVTKYGRNLGLNKVELFAIWAVLVVALLLTTANIYWIADQFGIHLATGTARKLLDAMASGASLGTAFAAILGVTLPAWALAAAGALGATAA | Lactobacillus gasseri | LA39 | [78] | |
| Reutericin 6 | MVTKYGRNLGLNKVELFAIWAVLVVALLLTTANIYWIADQFGIHLATGTARKLLDAMASGASLGTAFAAILGVTLPAWALAAAGALGATAA | Limosilactobacillus reuteri | LA 6 | [79] | |
| Class IId (leaderless) | |||||
| Bacteriocin LS2 | TNWKKIGKCYAGTLGSAVLGFGAMGPVGYWAGAGVGYASFC | Ligilactobacillus salivarius | BGHO1 | [80] | |
| Enterocin EJ97 | MLAKIKAMIKKFPNPYTLAAKLTTYEINWYKQQYGRYPWERPVA | Enterococcus faecalis | EJ97 | [81] | |
| Enterocin Q | MNFLKNGIAKWMTGAELQAYKKKYGCLPWEKISC | Enterococcus faecium | L50 | [82] | |
| LsbB | MKTILRFVAGYDIASHKKKTGGYPWERGKA | Lactococcus lactis | BGMN1-5 | [83] | |
| Lacticin Q | MAGFLKVVQLLAKYGSKAVQWAWANKGKILDWLNAGQAIDWVVSKIKQILGIK | Lactococcus lactis | QU 5 | [84] | |
| Lacticin Z | MAGFLKVVQILAKYGSKAVQWAWANKGKILDWINAGQAIDWVVEKIKQILGIK | Lactococcus lactis | QU 14 | [85] | |
| Lactolisterin BU | MWGRILGTVAKYGPKAVSWAWQHKWELINMGDLAFRYIQRIWG | Lactococcus lactis | BGBU1-4 | [86] | |
| Weisselicin Y | MANIVLRVGSVAYNYAPKIFKWIGEGVSYNQIIKWGHNKGWW | Weissella hellenica | QU 13 | [87] | |
| Weisselicin M | MVSAAKVALKVGWGLVKKYYTKVMQFIGEGWSVDQIADWLKRH | Weissella hellenica | QU 13 | [87] | |
| Enterocin L50 | L50A | MGAIAKLVAKFGWPIVKKYYKQIMQFIGEGWAINKIIEWIKKHI | Enterococcus faecalis | L50 | [88] |
| L50B | MGAIAKLVTKFGWPLIKKFYKQIMQFIGQGWTIDQIEKWLKRH | ||||
| Enterocin MR10 | MR10A | MGAIAKLVAKFGWPIVKKYYKQIMQFIGEGWAINKIIDWIKKHI | Enterococcus faecalis | MRR 10-3 | [89] |
| MR10B | MGAIAKLVAKFGWPFIKKFYKQIMQFIGQGWTIDQIEKWLKRH | ||||
| Garvicin KS | KS-A | MGAIIKAGAKIVGKGVLGGGASWLGWNVGEKIWK | Lactococcus garvieae | KS1546 | [90] |
| KS-B | MGAIIKAGAKIIGKGLLGGAAGGATYGGLKKIFG | ||||
| KS-C | MGAIIKAGAKIVGKGALTGGGVWLAEKLFGGK | ||||
| ClassIId (non-pediocin liner bacteriocins) | |||||
| Bovicin 255 | MNTKTFEQFDVMTDEALSTVEGGGKGYCKPVYYAANGYSCRYSNGEWGYVVTKGAFQATTDVIANGWVSSLGGGYFGKP | Streptococcus bovis | LRC 0255 | [91] | |
| Enterocin 96 | MLNKKLLENGVVNAVTIDELDAQFGGMSKRDCNLMKACCAGQAVTYAIHSLLNRLGGDSSDPAGCNDIVRKYCK | Enterococcus faecalis | WHE 96 | [92] | |
| Garvieacin Q | MENKNYTVLSDEELQKIDGGEYHLMNGANGYLTRVNGKYVYRVTKDPVSAVFGVISNGWGSAGAGFGPQH | Lactococcus garvieae | BCC 43578 | [93] | |
| Garvicins A | MENNNYTVLSDEELQKIDGGIGGALGNALNGLGTWANMMNGGGFVNQWQVYANKGKINQYRPY | Lactococcus garvieae | 21881 | [94] | |
| Lactococcin 972 | MKTKSLVLALSAVTLFSAGGIVAQAEGTWQHGYGVSSAYSNYHHGSKTHSATVVNNNTGRQGKDTQRAGVWAKATVGRNLTEKASFYYNFW | Lactococcus lactis | IPLA 972 | [95] | |
| Lactococcin A | MKNQLNFNIVSDEELSEANGGKLTFIQSTAAGDLYYNTNTHKYVYQQTQNAFGAAANTIVNGWMGGAAGGFGLHH | Lactococcus lactis | LMG 2130 | [96] | |
| Lactococcin B | MKNQLNFNIVSDEELAEVNGGSLQYVMSAGPYTWYKDTRTGKTICKQTIDTASYTFGVMAEGWGKTFH | Lactococcus lactis | 9B4 | [97] | |
| Enterocin B | MQNVKELSTKEMKQIIGGENDHRMPNELNRPNNLSKGGAKCGAAIAGGLFGIPKGPLAWAAGLANVYSKCN | Enterococcus faecium | T136 | [98] | |
| Carnobacteriocin A | MNNVKELSIKEMQQVTGGDQMSDGVNYGKGSSLSKGGAKCGLGIVGGLATIPSGPLGWLAGAAGVINSCMK | Carnobacterium piscicola | LV17A | [99] | |
| Bacteriocin | Number of Amino Acids | PIa | Net Charge | Ratio of Amino Acid Group | Most Enriched Amino Acids | Amino Acids Absent | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Polar | Non-Polar | Aromatic | Acidic | Basic | |||||||
| Class IIa | |||||||||||
| Acidocin A | 58 | 10.8 | 10.1 | 41% (24) | 24% (14) | 10% (6) | 2% (1) | 22% (13) | K(8)G(7)L(7) T(6) | ME | |
| Bacteriocin 31 | 43 | 9.7 | 4 | 40% (17) | 23% (10) | 16% (7) | 5% (2) | 16% (7) | G(5)N(4)W(4)K(4) | FM | |
| Bacteriocin BM1 | 43 | 8.9 | 2 | 47% (20) | 28% (12) | 9% (4) | 5% (2) | 12% (5) | G(7)A(5)I(4)K(4)I(4) | PFDR | |
| Bacteriocin B2 | 48 | 10 | 3.9 | 42% (20) | 38% (18) | 8% (4) | 2% (1) | 10% (5) | G(8)S(7)V(5)N(5) | LMWDH | |
| Bacteriocin T8 | 44 | 9.4 | 3 | 41% (18) | 20% (9) | 16% (7) | 7% (3) | 16% (7) | G(6)N(5)K(4)W(4) | FMS | |
| Carnobacteriocin B2 | 48 | 10 | 3.9 | 42% (20) | 35% (17) | 10% (5) | 2% (1) | 10% (5) | G(8)S(7)V(5)N(4) | LMDH | |
| Carnobacteriocin BM1 | 43 | 8.9 | 2 | 47% (20) | 28% (12) | 9% (4) | 5% (2) | 12% (5) | G(7)A(5)I(4)N(4)K(4) | PFDR | |
| Coagulin | 44 | 8.7 | 3.1 | 36% (16) | 36% (16) | 9% (4) | 2% (1) | 16% (7) | G(8)T(5)A(4)C(4)K(4) | LPFER | |
| Curvacin A | 41 | 9.6 | 2.9 | 44% (18) | 34% (14) | 10% (4) | 2% (1) | 10% (4) | G(8)A(4)S(4)N(4) | PFDH | |
| Divercin V41 | 43 | 8.7 | 2.8 | 47% (20) | 28% (12) | 14% (6) | 2% (1) | 9% (4) | G(10)V(4)C(4)K(4) | MFERH | |
| Enterocin A | 47 | 9.1 | 3.9 | 43% (20) | 32% (15) | 9% (4) | 2% (1) | 15% (7) | G(9)T(6)K(6)A(4)C(4) | QWER | |
| Enterocin CRL35 | 43 | 9.8 | 3.9 | 49% (21) | 28% (12) | 9% (4) | 2% (1) | 12% (5) | G(9)A(6)K(5)N(5) | EFHMPQR | |
| Enterocin HF | 43 | 10 | 4.9 | 49% (21) | 26% (11) | 9% (4) | 2% (1) | 14% (6) | G(10)K(6)A(5)N(5) | PMQFERH | |
| Enterocin P | 44 | 8.3 | 1 | 45% (20) | 30% (13) | 11% (5) | 5% (2) | 9% (4) | G(8)A(5)Q(5)S(4) | PQFD | |
| Enterocin SE-K4 | 43 | 9.7 | 3.9 | 37% (16) | 23% (10) | 16% (7) | 7% (3) | 16% (7) | G(5)V(5)Y(4)K(4) | HMP | |
| Leucocin A | 37 | 9 | 2.1 | 41% (15) | 27% (10) | 16% (6) | 3% (1) | 14% (5) | G(8)N(4)A(3)V(3) | IPMQD | |
| Mesentericin Y105 | 37 | 9 | 2.1 | 43% (16) | 27% (10) | 14% (5) | 3% (1) | 14% (5) | G(8)A(4)N(4)S(3) | PMQD | |
| Mundticin KS | 43 | 9.8 | 3.9 | 49% (21) | 28% (12) | 9% (4) | 2% (1) | 12% (5) | G(9)A(6)N(5)K(5) | PMQFERH | |
| Pediocin PA-1 | 44 | 8.7 | 3.1 | 36% (16) | 36% (16) | 9% (4) | 2% (1) | 16% (7) | G(8)A(4)C(4)N(4)T(4)K(4) | LPFER | |
| Piscicolin 126 | 44 | 9.5 | 2.9 | 48% (21) | 32% (14) | 9% (4) | 2% (1) | 9% (4) | G(9)N(7)A(6)K(4) | PMQFERH | |
| Plantaricin 423 | 37 | 8.7 | 3.1 | 32% (12) | 38% (14) | 14% (5) | 0% (0) | 16% (6) | G(6)S(5)C(4)K(3) | IPMDER | |
| Sakacin A | 41 | 9.6 | 2.9 | 44% (18) | 34% (14) | 10% (4) | 2% (1) | 10% (4) | G(8)A(4)S(4)N(4) | PFDH | |
| Sakacin P | 43 | 9 | 2.1 | 44% (19) | 30% (13) | 12% (5) | 2% (1) | 12% (5) | G(9)N(7)A(6)K(3) | LPMQFER | |
| Sakacin G | 38 | 7.9 | 1.1 | 42% (16) | 37%(14) | 11% (4) | 0% (0) | 11% (4) | G(9)N(5)C(4)V(4) | IPMFDER | |
| Unbericin A | 49 | 9.5 | 3 | 37% (18) | 41% (20) | 10% (5) | 2% (1) | 10% (5) | N(8)G(8)T(5)K(3) | DFP | |
| Bacteriocin L-1077 | 37 | 10.2 | 3 | 51% (19) | 22% (8) | 14% (5) | 3% (1) | 11% (4) | G(6)I(6)N(4)K(3)F(3) | CEHSW | |
| Bavaricin MN | 42 | 9.3 | 3 | 51% (21) | 22% (9) | 15% (6) | 2% (1) | 10% (4) | G(10)K(4)V(4)W(3)A(3) | EFHMR | |
| Bavaricin A | 39 | 8.3 | 1.1 | 49% (19) | 31% (12) | 8% (3) | 3% (1) | 10% (4) | G(9)N(6)A(6)T(3) | EFLMPQR | |
| Lactococcin MMFII | 37 | 7 | 0 | 32% (12) | 32% (12) | 11% (4) | 11% (4) | 14% (5) | S(4)K(4)T(3)G(3)N(3) | FMQR | |
| Leucocin C | 43 | 9 | 2.1 | 37% (16) | 37% (16) | 12% (5) | 2% (1) | 12% (5) | N(8)G(8)K(3)V(3) | EFPQR | |
| Mundticin | 41 | 9.5 | 2.9 | 49% (21) | 28% (12) | 9% (4) | 2% (1) | 12% (5) | G(9)A(6)K(5)N(5) | EFHMPQR | |
| Piscicocin VIa | 44 | 9.5 | 2.9 | 48% (21) | 32% (14) | 9% (4) | 2% (1) | 9% (4) | G(9)N(7)A(6)K(4) | EFHMPQR | |
| Plantaricin C19 | 36 | 9.9 | 5 | 39% (14) | 33% (12) | 11% (4) | 0% (0) | 17% (6) | G(7)K(4)N(3)A(3) | DEIMP | |
| plantaricin LPL-1 | 41 | 7.1 | 0.1 | 37% (15) | 29% (12) | 12% (5) | 7% (3) | 15% (6) | G(6)V(4)C(4)H(3) | MPQR | |
| Class IIb | |||||||||||
| ABP-118 | 118α | 45 | 9.1 | 1.9 | 76% (34) | 16% (7) | 4% (2) | 0% (0) | 4% (2) | G(13)A(7)L(6)V(4) | SQYWDEH |
| 118β | 46 | 9.8 | 3 | 63% (29) | 20% (9) | 9% (4) | 0% (0) | 9% (4) | G(15)A(5)S(4)I(4) | MQDE | |
| Acidocin LF221B | LF221B | 53 | 9.7 | 3.9 | 47% (25) | 32% (17) | 9% (5) | 2% (1) | 9% (5) | G(8)N(6)A(6)S(4) | DH |
| LF221β | 48 | 10 | 3.9 | 56% (27) | 23% (11) | 13% (6) | 0% (0) | 8% (4) | G(12)A(8)W(3)T(3) | DEHQ | |
| Brevicin 174A | 174A-β | 55 | 10.6 | 7.9 | 58% (32) | 22% (12) | 5% (3) | 0% (0) | 15% (8) | G(16)K(7)V(5)S(4) | QWDEH |
| 174A-γ | 53 | 10.7 | 7.1 | 55% (29) | 25% (13) | 4% (2) | 0% (0) | 17% (9) | A(11)G(9)K(7)V(3)T(3) | FYDER | |
| Carnobacteriocin XY | CbnX | 33 | 10.3 | 2 | 52% (17) | 24% (8) | 12% (4) | 3% (1) | 9% (3) | G(6)V(4)K(3)Q(3) | CYDRH |
| CbnY | 29 | 10.9 | 4 | 52% (15) | 21% (6) | 7% (2) | 3% (1) | 17% (5) | A(4)I(4)G(4)K(3) | PCWEH | |
| Enterocin X | EntXα | 40 | 9 | 2.1 | 43% (17) | 33% (13) | 10% (4) | 3% (1) | 13% (5) | G(5)I(4)L(3)K(3)S(3)N(3) | ER |
| EntXβ | 37 | 10.3 | 3.1 | 51% (19) | 11% (4) | 11% (4) | 8% (3) | 19% (7) | G(6)I(5)K(4)A(3) | CMTE | |
| Enterocin 1071 | Entα | 39 | 10.3 | 3.1 | 44% (17) | 23% (9) | 8% (3) | 8% (3) | 18% (7) | G(4)A(4)N(4)K(4) | CT |
| Entβ | 35 | 10.4 | 4 | 49% (17) | 14% (5) | 14% (5) | 6% (2) | 17% (6) | G(6)K(6)L(3)P(3)N(3)W(3) | SCMRH | |
| Gassericin S | GasA | 64 | 11.4 | 4.9 | 64% (41) | 20% (13) | 8% (5) | 0% (0) | 8% (5) | G(17)A(11)V(5)I(5) | DEHMY |
| GasX | 53 | 10 | 4.9 | 47% (25) | 34% (18) | 9% (5) | 0% (0) | 9% (5) | G(9)N(6)A(5)K(4) | DEH | |
| Gassericin T | GatA | 57 | 10.2 | 4 | 65% (37) | 16% (9) | 11% (6) | 0% (0) | 9% (5) | G(13)A(12)V(6)N(4) | DEMQS |
| GatX | 48 | 10 | 3.9 | 56%(27) | 23% (11) | 13% (6) | 0% (0) | 8% (4) | G(13)A(7)N(3)W(3) | DEHQ | |
| Mutacin IV | nlm A | 44 | 8.3 | 1 | 66% (29) | 20% (9) | 5% (2) | 2% (1) | 7% (3) | A(9)G(8)I(4)T(4) | DFMNQ |
| nlm B | 49 | 6 | −0.9 | 57% (28) | 20% (10) | 12% (6) | 4% (2) | 6% (3) | A(9)G(8)V(6)F(3) | EMR | |
| Lactococcin G | LcnGα | 39 | 10.6 | 4.1 | 38% (15) | 26% (10) | 8% (3) | 8% (3) | 21% (8) | G(6)K(4)A(3)V(3)I(3)N(3) | LPCFE |
| LcnGβ | 35 | 10.2 | 4 | 40% (14) | 6% (2) | 20% (7) | 11% (4) | 23% (8) | K(8)G(5)W(4)A(3)I(3) | SCMQTRH | |
| Lactococcin Q | LcnQα | 39 | 10.8 | 5.1 | 38% (15) | 28% (11) | 8% (3) | 5% (2) | 21% (8) | G(5)K(5)A(4)V(3)I(3)S(3)Q(3)T(3) | LPCTFE |
| LcnQβ | 35 | 10.2 | 4 | 43% (15) | 6% (2) | 17% (6) | 11% (4) | 23% (8) | K(8)G(6)W(4)A(3)E(3) | SCMQTYRH0 | |
| Lactocin 705 | 705α | 33 | 9.6 | 1.3 | 58% (19) | 18% (6) | 12% (4) | 0% (0) | 12% (4) | G(8)A(6)H(3)N(3) | CMTDEK |
| 705β | 33 | 10.3 | 4.2 | 48% (16) | 15% (5) | 12% (4) | 3% (1) | 21% (7) | G(7)K(4)Y(3)I(3)L(3) | VCTWE | |
| Plantaricin NC8 | PlnC8α | 29 | 10.3 | 4.1 | 31% (9) | 21% (6) | 24% (7) | 3% (1) | 21% (6) | L(4)K(4)Y(3)W(3) | ICME |
| PlnC8β | 34 | 10.4 | 5.2 | 32% (11) | 24% (8) | 18% (6) | 3% (1) | 24% (8) | K(5)S(4)I(3)Y(3) | CMQD | |
| Plantaricin S | Plsα | 27 | 10.4 | 3.1 | 56% (15) | 15% (4) | 15% (4) | 0% (0) | 15% (4) | A(7)L(4)G(3)K(2) | VPSCQDE |
| Plsβ | 26 | 10 | 2 | 42% (11) | 15% (4) | 19% (5) | 8% (2) | 15% (4) | A(5)K(4)G(3)S(2) | PCMTRH | |
| Plantaricin EF | PlnE | 33 | 12 | 5.1 | 52% (17) | 15% (5) | 9% (3) | 3% (1) | 21% (7) | G(6)R(4)V(4)I(4) | CEMPQTW |
| PlnF | 34 | 10.6 | 3.2 | 50% (17) | 15% (5) | 15% (5) | 3% (1) | 18% (6) | A(5)V(5)G(4)S(3)R(3) | CELMQT | |
| Plantaricin JK | PlnJ | 25 | 11.4 | 4 | 36% (9) | 12% (3) | 20% (5) | 8% (2) | 24% (6) | G(4)R(4)A(3)W(2)K(2) | CHMPQTV |
| PlnK | 32 | 10.9 | 5 | 47% (15) | 13% (4) | 13% (4) | 6% (2) | 22% (7) | G(6)R(5)A(4)Y(3) | CHMPQTV | |
| Salivaricin P | SalPα | 45 | 9.1 | 1.9 | 76% (34) | 16% (7) | 4% (2) | 0% (0) | 4% (2) | G(13) A(7)L(6)V(4) | DEHQSWY |
| SalPβ | 46 | 9.1 | 2.1 | 65% (30) | 17% (8) | 9% (4) | 0% (0) | 9% (4) | G(15)A(6)S(4)I(4) | DEMQT | |
| Thermophilin 13 | ThmA | 62 | 8.3 | 1 | 66% (41) | 15% (9) | 10% (6) | 3% (2) | 6% (4) | G(18)A(12)V(3)F(3) | EQRT |
| ThmB | 43 | 8.3 | 1 | 72% (31) | 19% (8) | 5% (2) | 0% (0) | 5% (2) | G(13)I(6)A(6)V(4) | DEMPRTY | |
| Uncharacterized | LSEI_2392 | 42 | 4.4 | −2.8 | 55% (23) | 17% (7) | 12% (5) | 10% (4) | 7% (3) | A(9)S(6)G(5)L(4) | CMNQRY |
| LSEI_2393 | 44 | 10.6 | 2 | 55% (24) | 23% (10) | 9% (4) | 5% (2) | 9% (4) | G(7)L(6)S(5)V(4) | CHMY | |
| Uncharacterized | LSEI_2405 | 44 | 12.2 | 5 | 64% (28) | 23% (10) | 2% (1) | 0% (0) | 11% (5) | A(8)G(7)L(4)T(3) | DEHMWY |
| LSEI_2406 | 58 | 9.1 | 1.9 | 64% (37) | 21% (12) | 5% (3) | 3% (2) | 7% (4) | G(16)A(7)I(6)T(5) | EHRWY | |
| Class IIc | |||||||||||
| Carnocyclin A | 60 | 10.5 | 4 | 67% (40) | 20% (12) | 3% (2) | 2% (1) | 8% (5) | A(9)G(9)V(8)I(8) | DCHMPRW | |
| Enterocin NKR-5-3B | 64 | 10.3 | 5 | 64% (41) | 19% (12) | 6% (4) | 2% (1) | 9% (6) | A(14)I(9)G(7)T(6)K(6)V(6) | PCMQFERH | |
| Enterocin AS-48 | 70 | 10.6 | 6 | 60% (42) | 14% (10) | 6% (4) | 6% (4) | 14% (10) | A(12)G(9)K(8)V(8) | DCHQ | |
| Garvicin ML | 60 | 10.6 | 5 | 60% (36) | 22% (13) | 7% (4) | 2% (1) | 10% (6) | A(15)L(6)G(6)K(6) | CEHPR | |
| Leucocyclicin Q | 61 | 10.2 | 2.1 | 66% (40) | 18% (11) | 5% (3) | 3% (2) | 8% (5) | A(14)L(9)G(6)I(5) | DCFMRY | |
| Lactocyclicin Q | 61 | 10.3 | 2.2 | 72% (44) | 8% (5) | 7% (4) | 3% (2) | 10% (6) | A(17)L(7)G(7)V(6) | CMFYE | |
| Plantaricyclin A | 58 | 9.9 | 1.1 | 67% (39) | 21% (12) | 5% (3) | 2% (1) | 5% (3) | A(14)L(9)G(7)V(6) | CEMNRY | |
| Plantacyclin B21AG | 58 | 10.6 | 2.1 | 67% (39) | 19% (11) | 5% (3) | 2% (1) | 7% (4) | A(12)L(9)G(8)V(6)T(6) | CEMNY | |
| Uberolysin | 70 | 10.1 | 3 | 60% (42) | 20% (14) | 7% (5) | 4% (3) | 9% (6) | A(14)I(11)S(7)V(7) | CEHMP | |
| Acidocin B | 58 | 7.7 | 0.1 | 69% (40) | 14% (8) | 9% (5) | 3% (2) | 5% (3) | A(18)L(8)G(7)T(5) | CEMN | |
| Butyrivibriocin AR10 | 58 | 3.7 | −2 | 62% (36) | 17% (10) | 14% (8) | 5% (3) | 2% (1) | A(13)I(8)G(6)V(5) | CHR | |
| Gassericin A | 58 | 7.7 | 0.1 | 67% (39) | 16% (9) | 9% (5) | 3% (2) | 5% (3) | A(18)L(8)G(7)T(5) | CEN | |
| Reutericin 6 | 58 | 7.7 | 0.1 | 67% (39) | 16% (9) | 9% (5) | 3% (2) | 5% (3) | A(18)L(8)G(7)T(5) | CEN | |
| Class IId (leaderless bacteriocins) | |||||||||||
| Bacteriocin LS2 | 41 | 9.4 | 2.9 | 56% (23) | 20% (8) | 17% (7) | 0% (0) | 7% (3) | G(10)A(6)K(3)Y(3)V(3) | DEHQR | |
| Enterocin EJ97 | 44 | 10.3 | 4 | 39% (17) | 20% (9) | 18% (8) | 5% (2) | 18% (8) | K(6)A(5)Y(5)P(4) | DCHS | |
| Epidermicin NI01 | 51 | 10.7 | 8.1 | 39% (20) | 22% (11) | 18% (9) | 2% (1) | 20% (10) | K(9)A(6)L(5)I(5) | DCPR | |
| Enterocin Q | 34 | 9.8 | 3.9 | 35% (12) | 26% (9) | 15% (5) | 6% (2) | 18% (6) | K(6)L(3)G(3)A(3) | DHRV | |
| LsbB | 30 | 10.6 | 5.1 | 40% (12) | 13% (4) | 13% (4) | 7% (2) | 27% (8) | K(5)G(4)A(3)Y(2) | CNQ | |
| Lacticin Q | 53 | 10.6 | 6 | 53% (28) | 17% (9) | 11% (6) | 4% (2) | 15% (8) | K(8)A(7)L(6)G(5)V(5)I(5) | CEHPRT | |
| Lacticin Z | 53 | 10.4 | 5 | 53% (28) | 15% (8) | 11% (6) | 6% (3) | 15% (8) | K(8)A(7)I(7)G(5)V(5) | CHPRT | |
| Lactolisterin BU | 43 | 10.6 | 4.1 | 44% (19) | 16% (7) | 19% (8) | 5% (2) | 16% (7) | W(5)G(5)I(4)A(4)R(3) | C | |
| Weisselicin Y | 42 | 10.3 | 4.1 | 45% (19) | 19% (8) | 19% (8) | 2% (1) | 14% (6) | I(5)G(5)N(4)V(4)K(4) | DCT | |
| Weisselicin M | 43 | 10.2 | 4.1 | 44% (19) | 16% (7) | 14% (6) | 7% (3) | 19% (8) | V(6)K(6)G(4)W(3) | CNP | |
| Enterocin L50 | L50A | 44 | 10.5 | 6.1 | 48% (21) | 11% (5) | 16% (7) | 5% (2) | 20% (9) | I(9)K(8)G(4)A(4) | DCRST |
| L50B | 43 | 10.7 | 6.1 | 40% (17) | 19% (8) | 16% (7) | 5% (2) | 21% (9) | K(7)I(6)G(4)Q(4) | CNS | |
| Enterocin MR10 | MR10A | 44 | 10.5 | 6.1 | 48% (21) | 11% (5) | 16% (7) | 5% (2) | 20% (9) | I(9)K(8)G(4)A(4) | CRST |
| MR10B | 43 | 10.7 | 6.1 | 40% (17) | 16% (7) | 19% (8) | 5% (2) | 21% (9) | K(7)I(6)G(4)F(4)Q(4) | CNS | |
| Garvicin KS | KS-A | 34 | 10.7 | 4 | 65% (22) | 9% (3) | 9% (3) | 3% (1) | 15% (5) | G(9)K(5)A(4)I(4) | DCFHPQRTY |
| KS-B | 34 | 10.8 | 5 | 74% (25) | 6% (2) | 6% (2) | 0% (0) | 15% (5) | G(11)A(6)I(5)K(5) | DCEHNPQRSVW | |
| KS-C | 32 | 10.7 | 4 | 69% (22) | 6% (2) | 6% (2) | 3% (1) | 16% (5) | G(9)A(5)K(5)I(3)L(3) | DCHNPQRSY | |
| Class IId (non-pediocin liner bacteriocins) | |||||||||||
| Bovicin 255 | 56 | 9.2 | 2.9 | 45% (25) | 23% (13) | 20% (11) | 4% (2) | 9% (5) | G(11)Y(7)V(5)A(5) | HM | |
| Enterocin 96 | 48 | 8.5 | 2.9 | 38% (18) | 33% (16) | 4% (2) | 8% (4) | 17% (8) | C(5)A(5)S(4)K(4)D(4) | EFW | |
| Garvieacin Q | 50 | 9.7 | 2.2 | 48% (24) | 22% (11) | 14% (7) | 4% (2) | 12% (6) | G(9)V(6)Y(4)N(4) | C | |
| Garvicin A | 43 | 10.2 | 3 | 49% (21) | 30% (13) | 14% (6) | 0% (0) | 7% (3) | G(9)N(7)A(4)L(3)Q(3)Y(3) | SCDEH | |
| Lactococcin 972 | 66 | 10 | 4.4 | 30% (20) | 33% (22) | 15% (10) | 5% (3) | 17% (11) | G(8)T(7)S(6)A(6)N(6) | CIMP | |
| Lactococcin A | 54 | 9.3 | 1.3 | 43% (23) | 31% (17) | 15% (8) | 2% (1) | 9% (5) | A(8)G(8)T(6)N(5) | CEPR | |
| Lactococcin B | 47 | 9.3 | 2 | 30% (14) | 34% (16) | 17% (8) | 6% (3) | 13% (6) | T(8)G(5)Y(4)K(4) | N | |
| Enterocin B | 53 | 9.6 | 3 | 53% (28) | 23% (12) | 6% (3) | 6% (3) | 13% (7) | G(8)A(8)N(7)L(5) | QT | |
| DivergicinA | 46 | 9.8 | 2.9 | 70% (32) | 22% (10) | 2% (1) | 0% (0) | 7% (3) | G(14)A(6)L(4)K(3)I(3) | SYWDERH | |
| Carnobacteriocin A | 53 | 9 | 1.9 | 58% (31) | 26% (14) | 4% (2) | 4% (2) | 8% (4) | G(13)S(6)L(5)A(5) | FERH | |
| Garvicins A | 43 | 10.2 | 3 | 49% (21) | 30% (13) | 14% (6) | 0% (0) | 7% (3) | G(9)N(7)A(4)L(3)Q(3) | SCDEH | |
| Species (Number of Strains Analyzed) | Percentage of Strains Harbor Class II Bcteriocin | Number of Class II Bacteriocins Identified | ||||||
|---|---|---|---|---|---|---|---|---|
| IIa | IIb | IIc | IId | IIa | IIb | IIc | IId | |
| Carnobacterium divergens (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Carnobacterium maltaromaticum (16) | 43.75% | 0 | 6.25% | 18.75% | 12 | 0 | 1 | 3 |
| Enterococcus faecalis (32) | 56.25% | 3.13% | 3.13% | 31.25% | 20 | 1 | 1 | 10 |
| Enterococcus faecium (30) | 83.33% | 46.66% | 13.33% | 76.66% | 33 | 16 | 4 | 31 |
| Enterococcus avium (5) | 0 | 0 | 0 | 20% | 0 | 0 | 0 | 1 |
| Enterococcus gallinarum (10) | 0 | 0 | 0 | 30% | 0 | 0 | 0 | 5 |
| Enterococcus durans (12) | 50% | 25% | 0 | 8.33% | 10 | 3 | 0 | 1 |
| Enterococcus casseliflavus (12) | 0 | 8.33% | 8.33% | 25% | 0 | 2 | 1 | 7 |
| Enterococcus mundtii (16) | 31.25% | 0 | 0 | 56.25% | 5 | 0 | 0 | 9 |
| Enterococcus sulfureus (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus acidophilus (18) | 94.44% | 100% | 0 | 100% | 17 | 20 | 53 | |
| Amylolactobacillus amylotrophicus (1) | 0.00% | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lentilactobacillus buchneri (5) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Levilactobacillus brevis (26) | 0 | 0 | 0 | 11.54% | 0 | 0 | 0 | 5 |
| Lacticaseibacillus casei (14) | 92.86% | 21.43% | 0 | 100% | 15 | 3 | 0 | 77 |
| Lactococcus chungangensis (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Secundilactobacillus collinoides (2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Loigolactobacillus coryniformis subsp. torquens (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Loigolactobacillus coryniformis subsp. coryniformis (3) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus crispatus (22) | 36.36% | 50% | 0 | 72.73% | 17 | 11 | 0 | 27 |
| Companilactobacillus crustorum (3) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus delbrueckii subsp. bulgaricus (14) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus delbrueckii subsp. indicus (2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus delbrueckii subsp. sunkii (1) | 100% | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Lactobacillus delbrueckii subsp. lactis (5) | 0 | 0 | 0 | 80% | 0 | 0 | 0 | 4 |
| Lactobacillus delbrueckii subsp. delbrueckii (5) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lapidilactobacillus dextrinicus (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus equicursoris (2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactococcus fujiensis (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Fructilactobacillus fructivorans (4) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus gasseri (23) | 39.13% | 34.78% | 39.13% | 52.17% | 9 | 8 | 12 | 26 |
| Lactobacillus helveticus (27) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus iners (17) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactobacillus johnsonii (22) | 72.73% | 54.55% | 0 | 31.82% | 27 | 12 | 0 | 7 |
| Apilactobacillus kunkeei (10) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lacticaseibacillus paracasei (20) | 100% | 35% | 0 | 100% | 21 | 7 | 0 | 94 |
| Schleiferilactobacillus perolens (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactiplantibacillus plantarum (50) | 0 | 80% | 4% | 80% | 0 | 72 | 2 | 73 |
| Limosilactobacillus reuteri (31) | 0 | 6.45% | 0 | 6.45% | 0 | 2 | 0 | 2 |
| Lacticaseibacillus rhamnosus (30) | 100% | 20% | 100% | 32 | 6 | 0 | 76 | |
| Furfurilactobacillus rossiae (1) | 100% | 0 | 0 | 100% | 1 | 0 | 0 | 1 |
| Latilactobacillus sakei (25) | 32% | 16% | 0 | 56% | 10 | 4 | 0 | 20 |
| Ligilactobacillus salivarius (30) | 0% | 50% | 3.33% | 56.67% | 0 | 16 | 1 | 31 |
| Paucilactobacillus vaccinostercus (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactococcus garvieae (19) | 0 | 0 | 5.26% | 26.32% | 0 | 0 | 1 | 7 |
| Lactococcus lactis subsp. cremoris (21) | 0 | 0 | 0 | 85.71% | 0 | 0 | 0 | 37 |
| Lactococcus lactis subsp. hordniae (2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Lactococcus lactis subsp. lactis (20) | 0 | 0 | 0 | 50% | 0 | 0 | 0 | 18 |
| Lactococcus lactis subsp. tructae (1) | 0 | 0 | 0 | 100% | 0 | 0 | 0 | 4 |
| Lactococcus piscium (4) | 25% | 0 | 0 | 50% | 1 | 0 | 0 | 3 |
| Lactococcus raffinolactis (4) | 50% | 0 | 25% | 100% | 2 | 0 | 1 | 5 |
| Leuconostoc carnosum (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leuconostoc citreum (9) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leuconostoc fallax (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leuconostoc gelidum (11) | 27.27% | 0 | 0 | 72.73% | 3 | 0 | 0 | 14 |
| Leuconostoc kimchi (1) | 0 | 0 | 0 | 100% | 0 | 0 | 0 | 1 |
| Leuconostoc lactis (7) | 0 | 0 | 0 | 71.43% | 0 | 0 | 0 | 5 |
| Leuconostoc mesenteroides (40) | 5% | 0 | 0 | 37.50% | 2 | 0 | 0 | 17 |
| Leuconostoc pseudomesenteroides (23) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Oenococcus oeni (40) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pediococcus pentosaceus (12) | 58.33% | 0 | 0 | 8.30% | 12 | 0 | 0 | 1 |
| Pediococcus acidilactici (22) | 4.50% | 0 | 0 | 4.50% | 2 | 0 | 0 | 1 |
| Pediococcus damnosus (11) | 100% | 0 | 0 | 18.18% | 12 | 0 | 0 | 2 |
| Pediococcus parvulus (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pediococcus inopinatus (2) | 0 | 0 | 0 | 100% | 0 | 0 | 0 | 2 |
| Streptococcus thermophilus (42) | 0 | 9.52% | 0 | 100% | 0 | 5 | 0 | 189 |
| Weissella confusa (4) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Weissella cibaria (15) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Weissella kandleri (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Weissella koreensis (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Weissella oryzae (1) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Weissella viridescens (2) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Zhang, Y.; Li, L.; Jiang, X.; Chen, Z.; Zhao, F.; Yi, Y. Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria. Fermentation 2022, 8, 217. https://doi.org/10.3390/fermentation8050217
Zhang T, Zhang Y, Li L, Jiang X, Chen Z, Zhao F, Yi Y. Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria. Fermentation. 2022; 8(5):217. https://doi.org/10.3390/fermentation8050217
Chicago/Turabian StyleZhang, Tingting, Yu Zhang, Lin Li, Xiuqi Jiang, Zhuo Chen, Fan Zhao, and Yanglei Yi. 2022. "Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria" Fermentation 8, no. 5: 217. https://doi.org/10.3390/fermentation8050217
APA StyleZhang, T., Zhang, Y., Li, L., Jiang, X., Chen, Z., Zhao, F., & Yi, Y. (2022). Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria. Fermentation, 8(5), 217. https://doi.org/10.3390/fermentation8050217