

Isolation, Purification, and Antitumor Activity of a Novel Active Protein from Antrodia cinnamomea Liquid Fermentation Mycelia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction of Mycelial Proteins (MP)
2.3. Isolation and Purification of AC Antitumor Active Protein (ACAP)
2.4. In Vitro Antitumor Activity Assay
2.5. Flow Cytometric Analysis of Apoptosis
2.6. Western Blotting Analysis
2.7. Statistical Analysis
3. Results
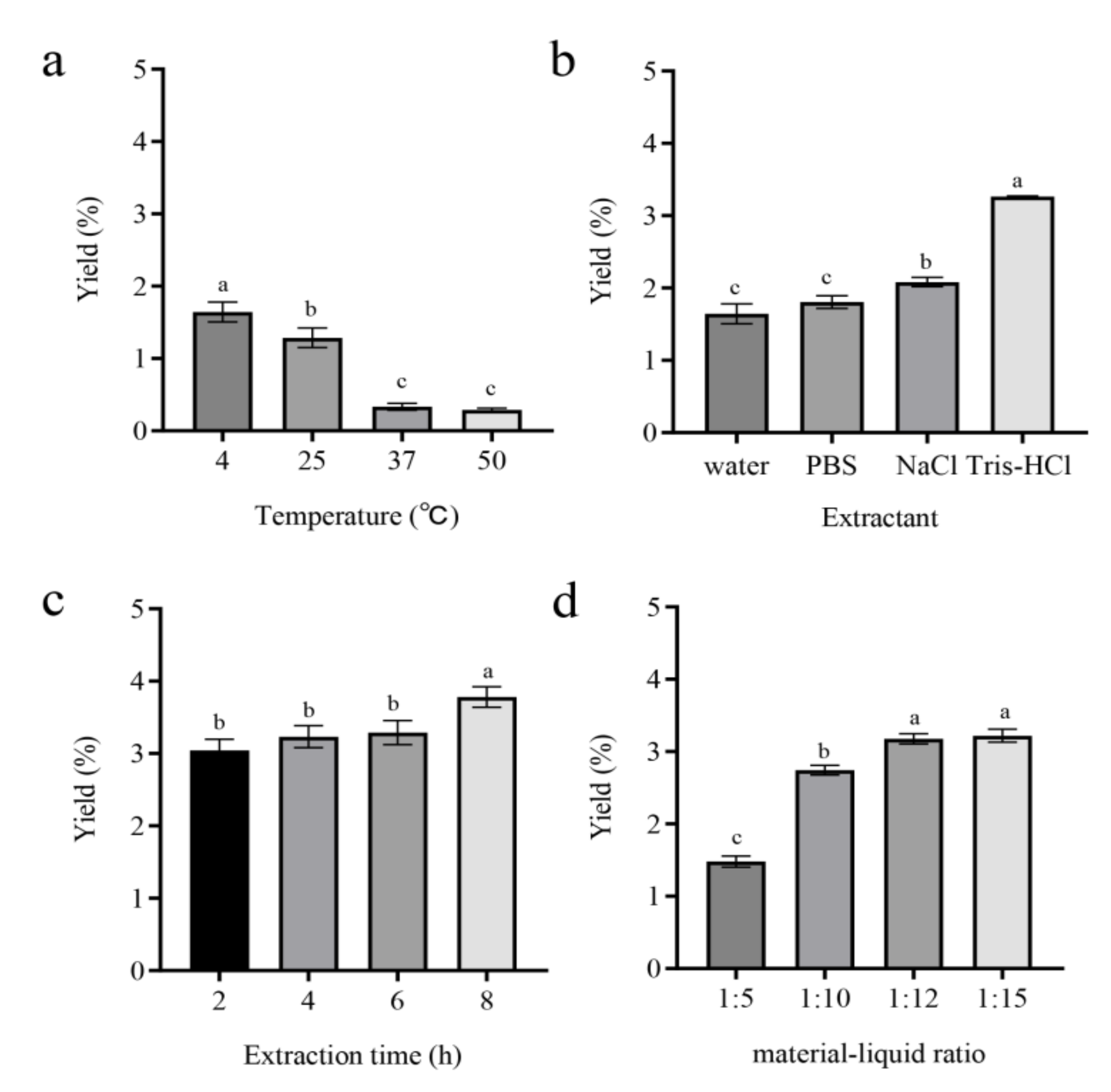
3.1. Effects of Various Factors on MP Extraction Yield
3.2. Orthogonal Test for MP Extraction
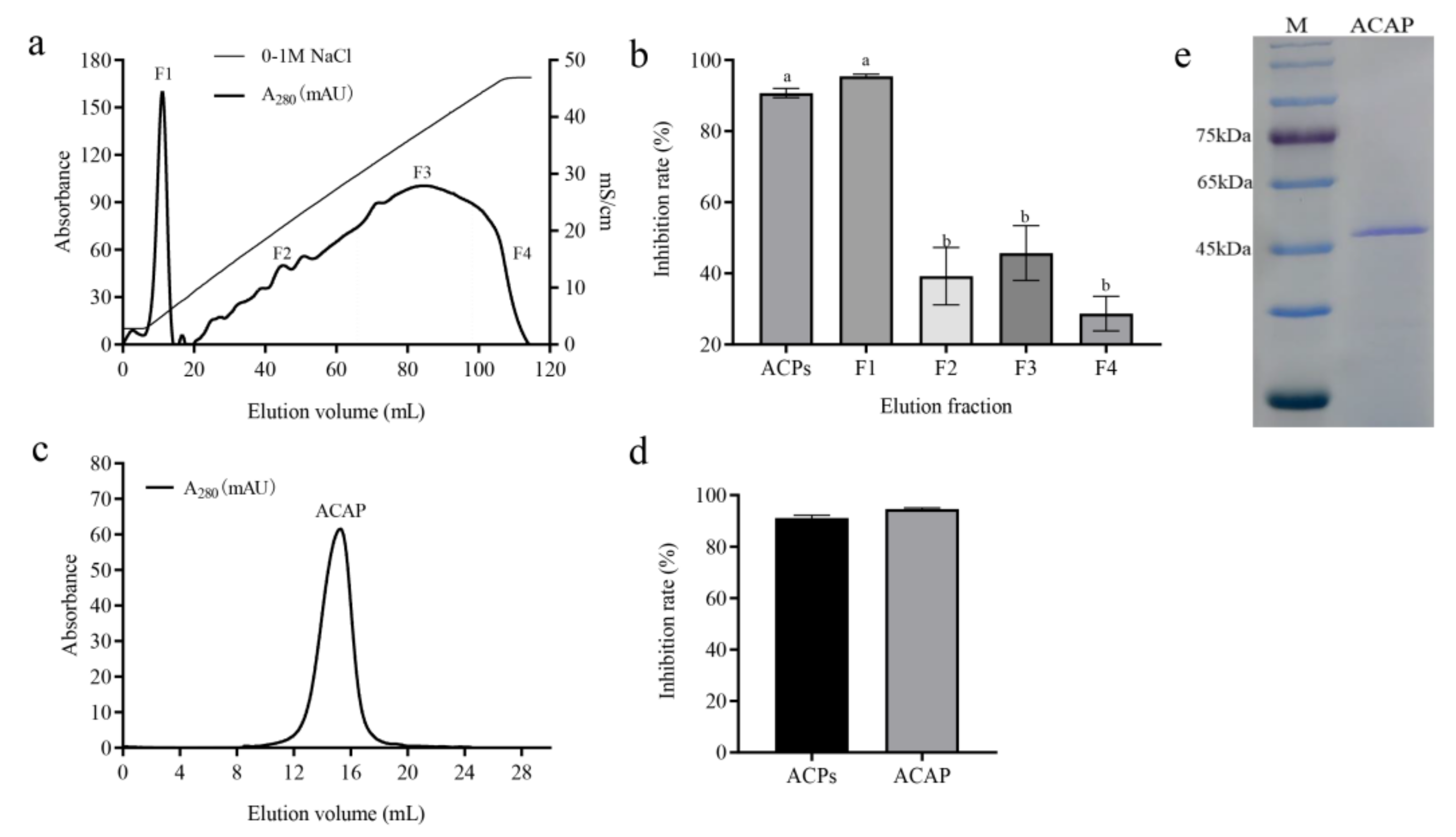
3.3. ACAP Purification by Ion Exchange Chromatography and Gel Filtration Chromatography
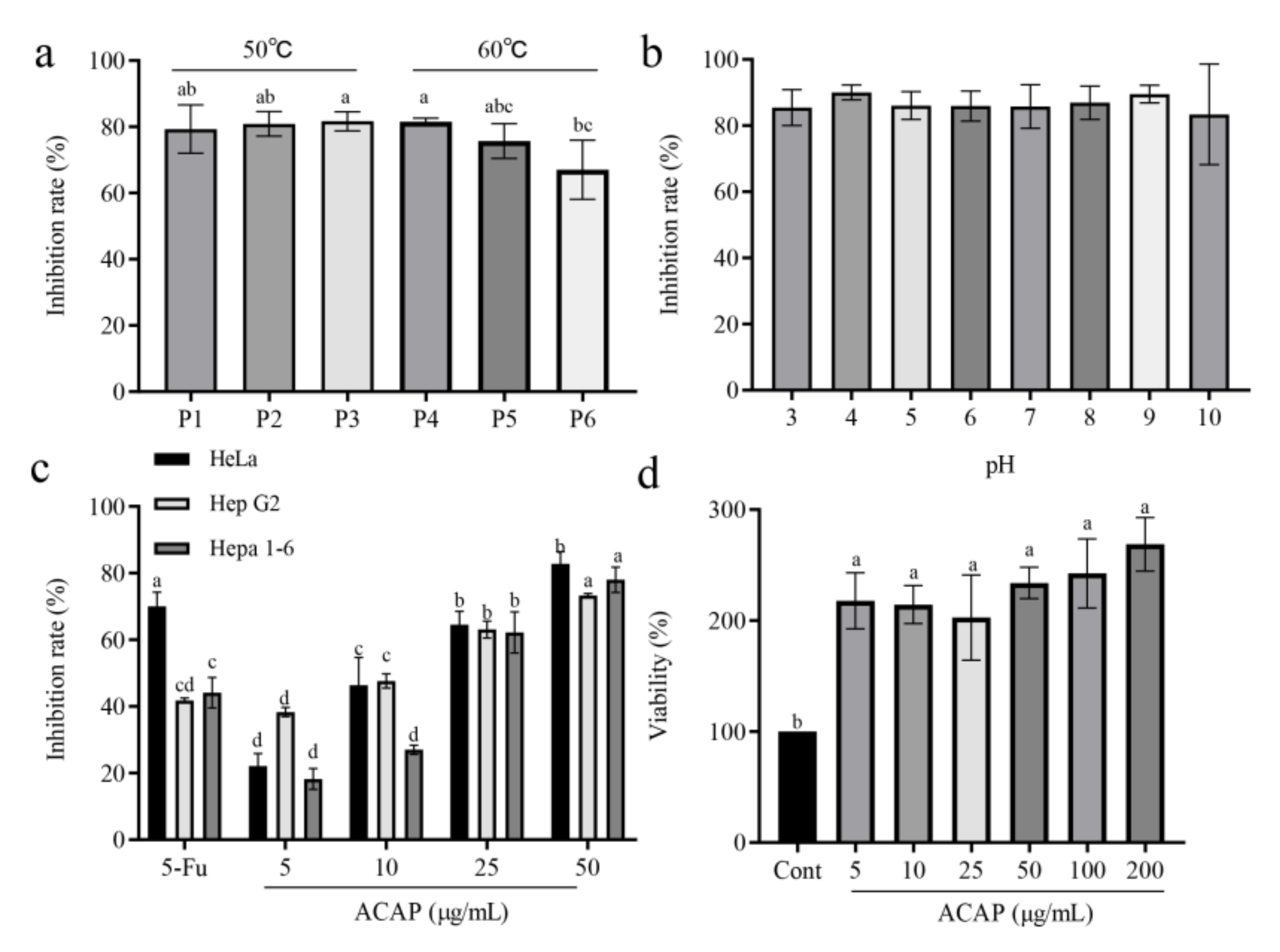
3.4. Effects of ACAP on Tumor Cell Proliferation
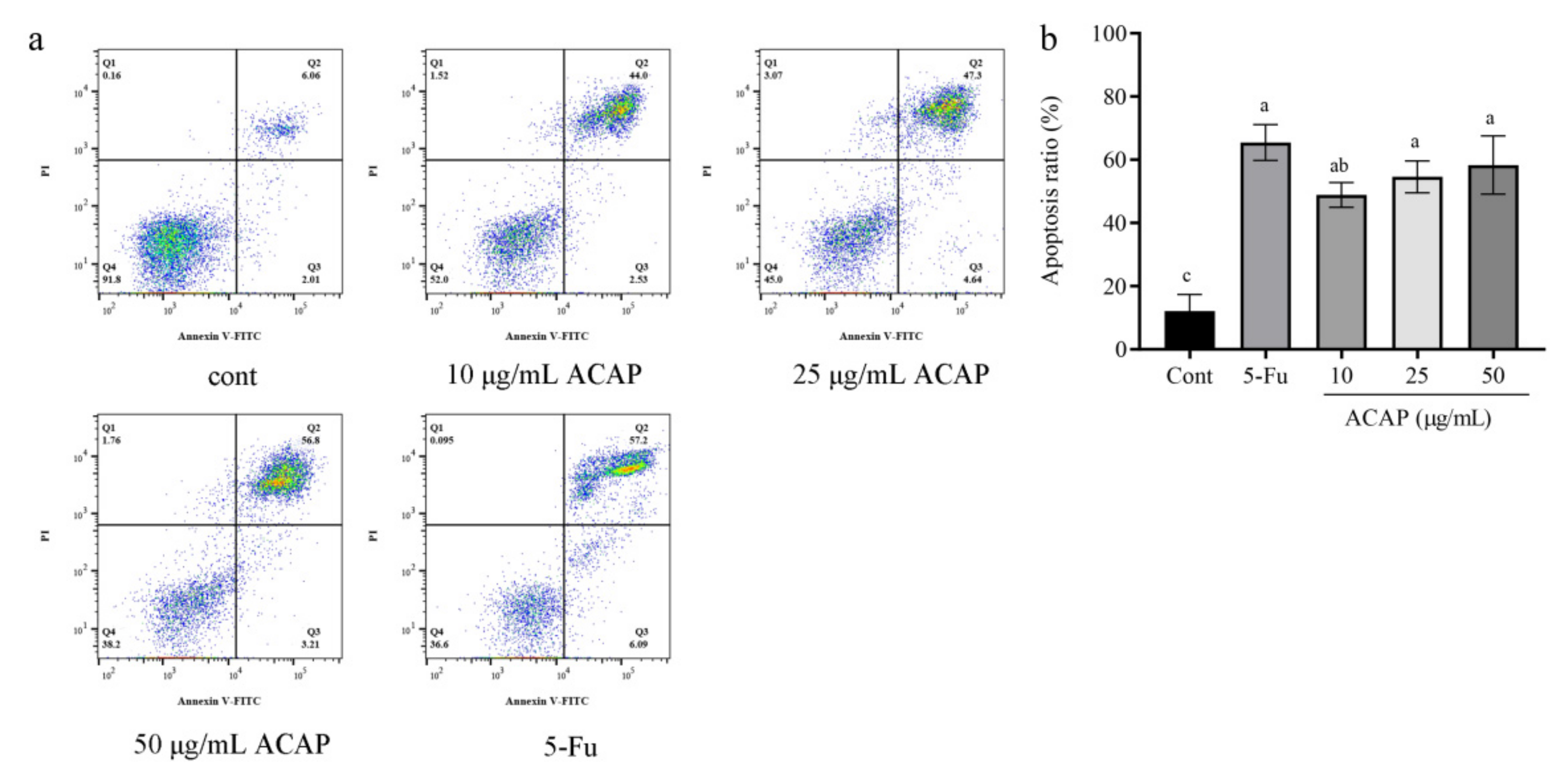
3.5. ACAP Induced Apoptosis of Hep G2 Cells
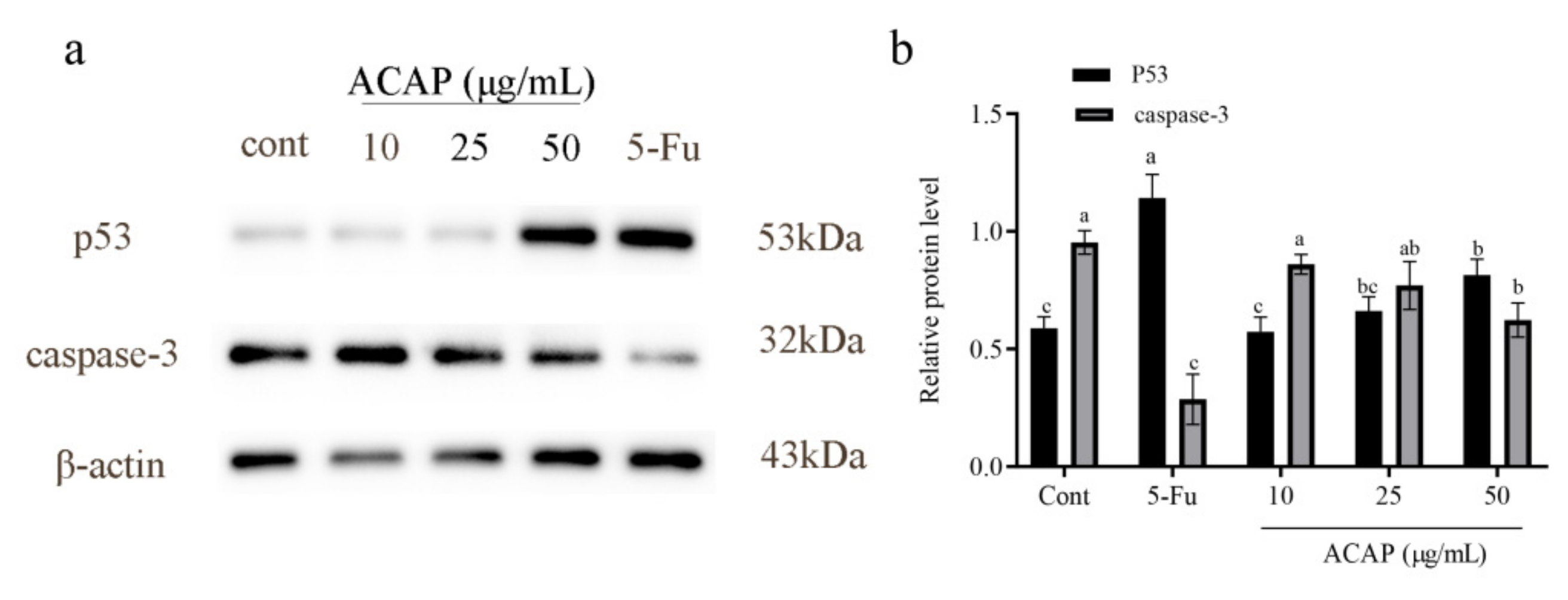
3.6. ACAP Affected Expression Levels of Apoptotic Pathway Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, M.; El-Shazly, M.; Wu, T.; Du, Y.; Chang, T.; Chen, C.; Hsu, Y.M.; Lai, K.; Chiu, C.P.; Chang, F.; et al. Recent research and development of Antrodia cinnamomea. Pharmacol. Ther. 2013, 139, 124–156. [Google Scholar] [CrossRef] [PubMed]
- Ao, Z.; Xu, Z.; Lu, Z.; Xu, H.; Zhang, X.; Dou, W. Niuchangchih (Antrodia camphorata) and its potential in treating liver diseases. J. Ethnopharmacol. 2009, 121, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Geethangili, M.; Tzeng, Y.M. Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evid.-based Complement Altern. 2011, 2011, 212641. [Google Scholar]
- Zhang, B.; Hu, P.; Huang, J.; Hu, Y.; Chen, L.; Xu, G. Current advances on the structure, bioactivity, synthesis, and metabolic regulation of novel ubiquinone derivatives in the edible and medicinal mushroom Antrodia cinnamomea. J. Agric. Food Chem. 2017, 65, 10395–10405. [Google Scholar] [CrossRef]
- Zhang, B.; Guan, Y.; Hu, P.; Chen, L.; Xu, G.; Liu, L.; Cheung, P.C.K. Production of bioactive metabolites by submerged fermentation of the medicinal mushroom Antrodia cinnamomea: Recent advances and future development. Crit. Rev. Biotechnol. 2019, 39, 541–554. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Li, D.; Zang, W.; Zhu, H.; Wu, P.; Mei, Y.; Liang, Y. A polysaccharide from Antrodia cinnamomea mycelia exerts antitumor activity through blocking of TOP1/TDP1-mediated DNA repair pathway. Int. J. Biol. 2018, 120, 1551–1560. [Google Scholar] [CrossRef]
- Lee, Y.P.; Tsai, W.C.; Ko, C.J.; Rao, Y.; Yang, C.; Chen, D.; Yang, M.; Yang, C.; Tzeng, Y.M. Anticancer effects of eleven triterpenoids derived from Antrodia camphorata. Anticancer Res. 2012, 32, 2727–2734. [Google Scholar]
- Wu, T.; Huang, T.; Martel, J.; Liau, J.C.; Chiu, C.Y.; Leu, Y.L.; Jian, W.T.; Chang, I.; Lu, C.; Ojcius, D.M.; et al. Pinicolol B from Antrodia cinnamomea induces apoptosis of nasopharyngeal carcinoma cells. J. Ethnopharmacol. 2017, 201, 117–122. [Google Scholar] [CrossRef]
- Yeh, C.T.; Rao, Y.; Yao, C.; Yeh, C.F.; Li, C.H.; Chuang, S.; Luong, J.H.T.; Lai, G.; Tzeng, Y.M. Cytotoxic triterpenes from Antrodia camphorata and their mode of action in HT-29 human colon cancer cells. Cancer Lett. 2009, 285, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Yen, I.C.; Shi, L.; Chung, M.; Ahmetaj-Shala, B.; Chang, T.; Lee, S.Y. Antrolone, a novel benzoid derived from Antrodia cinnamomea, inhibits the LPS-induced inflammatory response in RAW264.7 macrophage cells by balancing the NF-kappa B and Nrf2 pathways. Am. J. Chin. Med. 2018, 46, 1297–1313. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, A.; Qu, Y.; Wang, Z.; Zhang, Y.; Liu, Y.; Wang, N.; Teng, L.; Wang, D. Ameliorative effects of Antrodia cinnamomea polysaccharides against cyclophosphamide-induced immunosuppression related to Nrf2/HO-1 signaling in BALB/c mice. Int. J. Biol. Macromol. 2018, 116, 8–15. [Google Scholar] [CrossRef]
- Huang, T.; Lan, Y.; Ko, Y.; Chen, C.; Lai, H.; Ojcius, D.M.; Martel, J.; Young, J.D.; Chong, K. Antrodia cinnamomea produces anti-angiogenic effects by inhibiting the VEGFR2 signaling pathway. J. Ethnopharmacol. 2018, 220, 239–249. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, C.; Tong, X.; Qi, Y. A lectin with mycelia differentiation and antiphytovirus activities from the edible mushroom Agrocybe aegerita. J. Biochem. Mol. Biol. 2003, 36, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Lam, S.K.; Ng, T.B. First simultaneous isolation of a ribosome inactivating protein and an antifungal protein from a mushroom (Lyophyllum shimeji) together with evidence for synergism of their antifungal effects. Arch. Biochem. Biophys. 2001, 393, 271–280. [Google Scholar] [CrossRef]
- El-Maradny, Y.A.; El-Fakharany, E.M.; Abu-Serie, M.M.; Hashish, M.H.; Selim, H.S. Lectins purified from medicinal and edible mushrooms: Insights into their antiviral activity against pathogenic viruses. Int. J. Biol. Macromol. 2021, 179, 239–258. [Google Scholar] [CrossRef]
- Cui, F.; Li, Y.; Zan, X.; Yang, Y.; Sun, W.; Qian, J.; Zhou, Q.; Yu, S. Purification and partial characterization of a novel hemagglutinating glycoprotein from the cultured mycelia of Hericium erinaceus. Process Biochem. 2014, 49, 1362–1369. [Google Scholar] [CrossRef]
- Kino, K.; Yamashita, A.; Yamaoka, K.; Watanabe, J.; Tanaka, S.; Ko, K.; Shimizu, K.; Tsunoo, H. Isolation and characterization of a new immunomodulatory protein, ling zhi-8 (LZ-8), from Ganoderma-lucidium. J. Biol. Chem. 1989, 264, 472–478. [Google Scholar] [CrossRef]
- Dai, R.; Liu, M.; Nabil, W.N.N.; Xi, Z.; Xu, H. Mycomedicine: A unique class of natural products with potent anti-tumour bioactivities. Molecules 2021, 26, 1113. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, R.; Liu, F.; Ng, T.B. Purification and characterization of a novel protein with activity against non-small-cell lung cancer in vitro and in vivo from the edible mushroom Boletus edulis. Int. J. Biol. Macromol. 2021, 174, 77–88. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Gao, W.; Zhou, R.; Liu, F.; Ng, T.B. A novel antitumor protein from the mushroom Pholiota nameko induces apoptosis of human breast adenocarcinoma MCF-7 cells in vivo and modulates cytokine secretion in mice bearing MCF-7 xenografts. J. Biol. Macromol. 2020, 164, 3171–3178. [Google Scholar] [CrossRef]
- Sheu, F.; Chien, P.J.; Hsieh, K.Y.; Chin, K.L.; Huang, W.; Tsao, C.Y.; Chen, Y.; Cheng, H.; Chang, H. Purification, cloning, and functional characterization of a novel immunomodulatory protein from Antrodia camphorata (Bitter Mushroom) that exhibits TLR2-dependent NF-kappa B activation and m1 polarization within murine macrophages. J. Agric. Food Chem. 2009, 57, 4130–4141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, F.; Ng, T.B. Interrelationship among paraptosis, apoptosis and autophagy in lung cancer A549 cells induced by BEAP, an antitumor protein isolated from the edible porcini mushroom Boletus edulis. Int. J. Biol. 2021, 188, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Hsiao, Y.M.; Wu, M.; Ou, C.; Lin, Y.; Lue, K.; Ko, J.L. Interruption of lung cancer cell migration and proliferation by fungal immunomodulatory protein FIP-fve from Flammulina velutipes. J. Agric. Food Chem. 2013, 61, 12044–12052. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.; Mischi, M.; Scheepens, W.; De la Rosette, J.J.; Wijkstra, H. Angiogenesis in prostate cancer: Onset, progression and imaging. BJU Int. 2012, 110, E794–E808. [Google Scholar] [CrossRef]
- Tsai, P.F.; Ma, C.; Wu, J.S.B. A novel glycoprotein from mushroom Hypsizygus marmoreus (Peck) Bigelow with growth inhibitory effect against human leukaemic U937 cells. Food Chem. 2013, 141, 1252–1258. [Google Scholar] [CrossRef]
- Lin, C.; Sheu, G.T.; Lin, Y.; Yeh, C.S.; Huang, Y.; Lai, Y.; Chang, J.; Ko, J.L. A new immunomodulatory protein from Ganoderma microsporum inhibits epidermal growth factor mediated migration and invasion in A549 lung cancer cells. Process Biochem. 2010, 45, 1537–1542. [Google Scholar] [CrossRef]
- Cui, F.; Zan, X.; Li, Y.; Yang, Y.; Sun, W.; Zhou, Q.; Yu, S.; Dong, Y. Purification and partial characterization of a novel anti-tumor glycoprotein from cultured mycelia of Grifola frondosa. Int. J. Biol. Macromol. 2013, 62, 684–690. [Google Scholar] [CrossRef]
- Prateep, A.; Sumkhemthong, S.; Suksomtip, M.; Chanvorachote, P.; Chaotham, C. Peptides extracted from edible mushroom: Lentinus squarrosulus induces apoptosis in human lung cancer cells. Pharm. Biol. 2017, 55, 1792–1799. [Google Scholar] [CrossRef] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Wachter, F.; Grunert, M.; Blaj, C.; Weinstock, D.M.; Jeremias, I.; Ehrhardt, H. Impact of the p53 status of tumor cells on extrinsic and intrinsic apoptosis signaling. Cell Commun. Signal. 2013, 11, 27. [Google Scholar] [CrossRef] [Green Version]
- Fridman, J.S.; Lowe, S.W. Control of apoptosis by p53. Oncogene 2003, 22, 9030–9040. [Google Scholar] [CrossRef] [Green Version]
- Yuan, B.; Ma, N.; Zhao, L.; Zhao, E.; Gao, Z.; Wang, W.; Song, M.; Zhang, G.; Hu, Q.; Xiao, H. In vitro and in vivo inhibitory effects of a Pleurotus eryngii protein on colon cancer cells. Food Funct. 2017, 8, 3553–3562. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Hou, L.; Xin, F.; Fan, B.; Lu, C.; Zhang, L.; Wang, F.; Li, S. Immunomodulatory protein from Nectria haematococca induces apoptosis in lung cancer cells via the p53 pathway. Int. J. Mol. Sci. 2019, 20, 5348. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Factors | |||
|---|---|---|---|---|
| A 1 | B 2 | C 3 | D 4 | |
| 1 | 10% NaCl | 6 | 4 | 1:10 |
| 2 | Distilled water | 8 | 25 | 1:15 |
| 3 | Tris-HCl | 10 | 37 | 1:20 |
| Experimental Number | A | B | C | D | Protein Yield (%) |
|---|---|---|---|---|---|
| 1 | 1 | 1 | 1 | 1 | 0.609 |
| 2 | 1 | 2 | 2 | 2 | 0.461 |
| 3 | 1 | 3 | 3 | 3 | 0.221 |
| 4 | 2 | 1 | 2 | 3 | 0.574 |
| 5 | 2 | 2 | 3 | 1 | 0.165 |
| 6 | 2 | 3 | 1 | 2 | 0.972 |
| 7 | 3 | 1 | 3 | 2 | 0.624 |
| 8 | 3 | 2 | 1 | 3 | 4.390 |
| 9 | 3 | 3 | 2 | 1 | 2.177 |
| K1 1 | 1.291 | 1.807 | 5.971 | 2.951 | |
| K2 1 | 1.711 | 5.016 | 3.212 | 2.057 | |
| K3 1 | 7.191 | 3.370 | 1.010 | 5.185 | |
| K1 2 | 0.430 | 0.602 | 1.990 | 0.984 | |
| K2 2 | 0.570 | 1.672 | 1.071 | 0.686 | |
| K3 2 | 2.397 | 1.123 | 0.337 | 1.728 | |
| Range | 1.967 | 1.070 | 1.654 | 1.043 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Ge, J.; Li, Y.; Zheng, S.; Liu, Y.; Liang, Y.; Mei, Y. Isolation, Purification, and Antitumor Activity of a Novel Active Protein from Antrodia cinnamomea Liquid Fermentation Mycelia. Fermentation 2023, 9, 185. https://doi.org/10.3390/fermentation9020185
Li Y, Ge J, Li Y, Zheng S, Liu Y, Liang Y, Mei Y. Isolation, Purification, and Antitumor Activity of a Novel Active Protein from Antrodia cinnamomea Liquid Fermentation Mycelia. Fermentation. 2023; 9(2):185. https://doi.org/10.3390/fermentation9020185
Chicago/Turabian StyleLi, Yingying, Jiaxin Ge, Yanbin Li, Siqi Zheng, Yangyang Liu, Yunxiang Liang, and Yuxia Mei. 2023. "Isolation, Purification, and Antitumor Activity of a Novel Active Protein from Antrodia cinnamomea Liquid Fermentation Mycelia" Fermentation 9, no. 2: 185. https://doi.org/10.3390/fermentation9020185
APA StyleLi, Y., Ge, J., Li, Y., Zheng, S., Liu, Y., Liang, Y., & Mei, Y. (2023). Isolation, Purification, and Antitumor Activity of a Novel Active Protein from Antrodia cinnamomea Liquid Fermentation Mycelia. Fermentation, 9(2), 185. https://doi.org/10.3390/fermentation9020185