

Solid-State Fermentation of Soybean Meal with Edible Mushroom Mycelium to Improve Its Nutritional, Antioxidant Capacities and Physicochemical Properties

 , , , ,
, , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Strain Culture
2.3. Solid-State Fermentation of Soybean Meal
2.4. Macronutrients Content Analysis
2.5. Determination of Amino Acid Content
2.6. Determination of Soybean Isoflavones
2.7. Determination of Total Polyphenol Content
2.8. Determination of Antioxidant Capacity
2.8.1. Determination of DPPH Radical Scavenging Activity
2.8.2. Determination of Hydroxyl Radical Scavenging Activity
2.8.3. Determination of ABTS+ Radical Scavenging Activity
2.9. Color
2.10. Particle Size
2.11. Scanning Electron Microscope (SEM)
2.12. Statistical Analysis
3. Results
3.1. Macronutrients Content
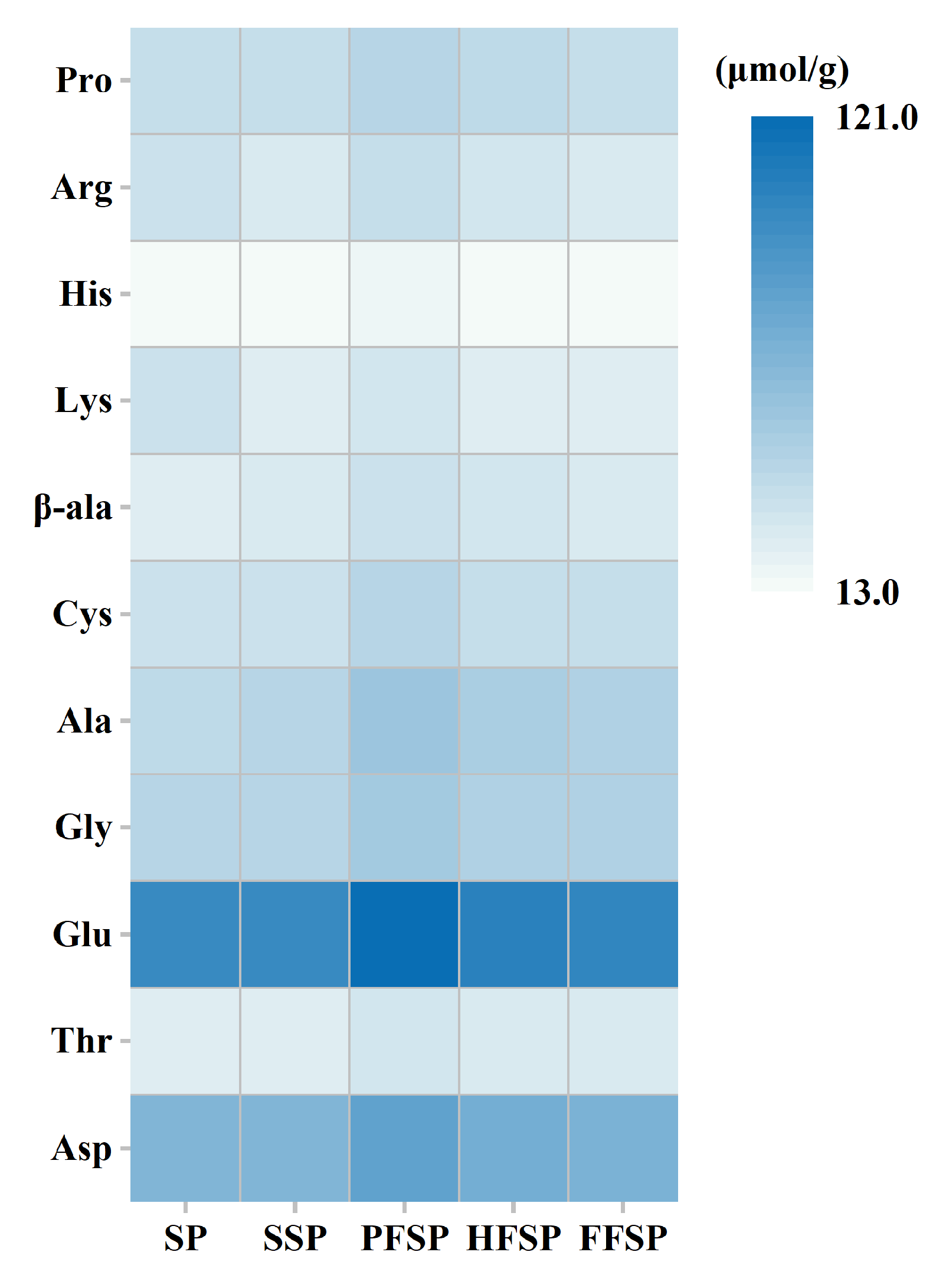
3.2. Amino Acid Content
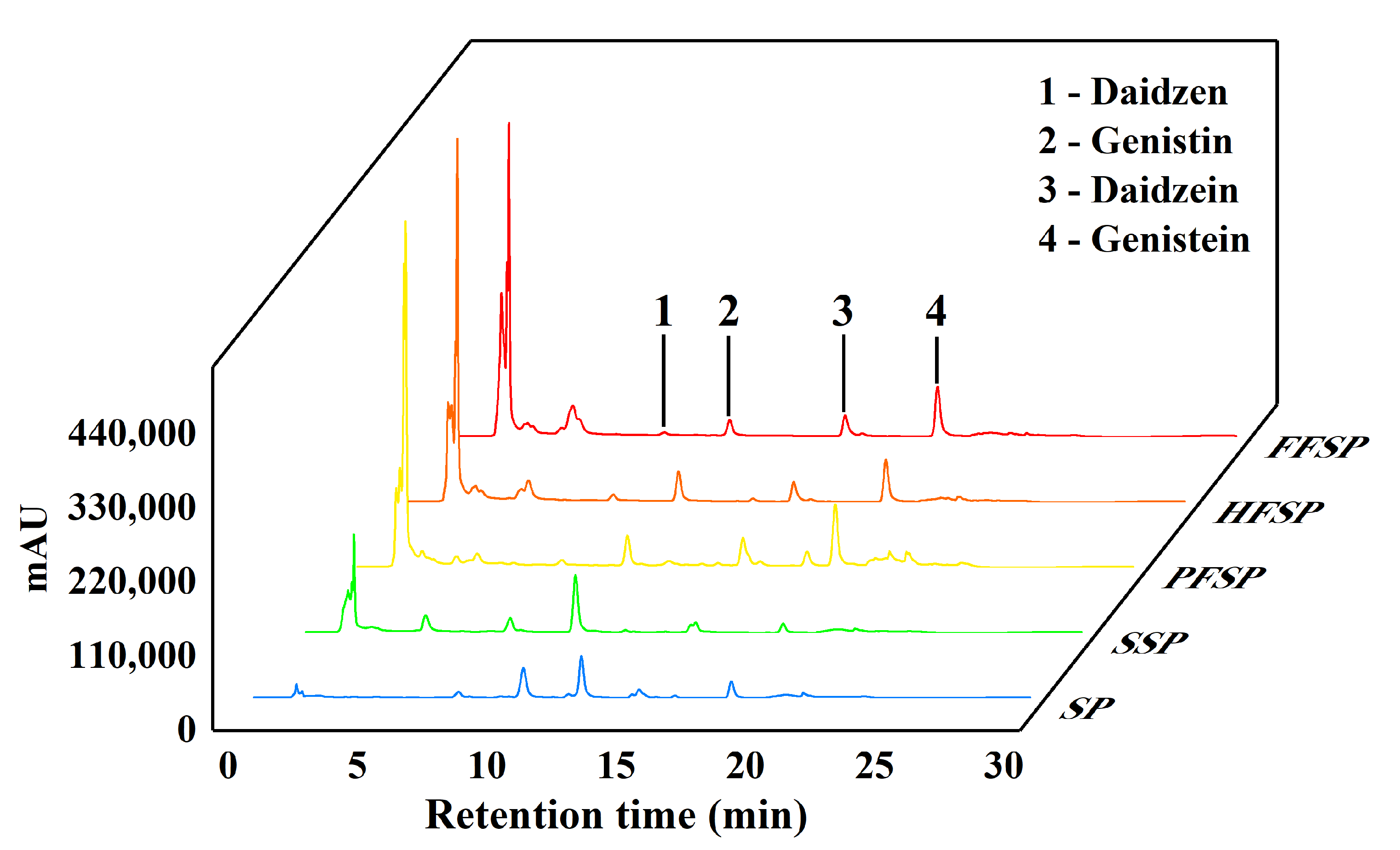
3.3. Isoflavone Content
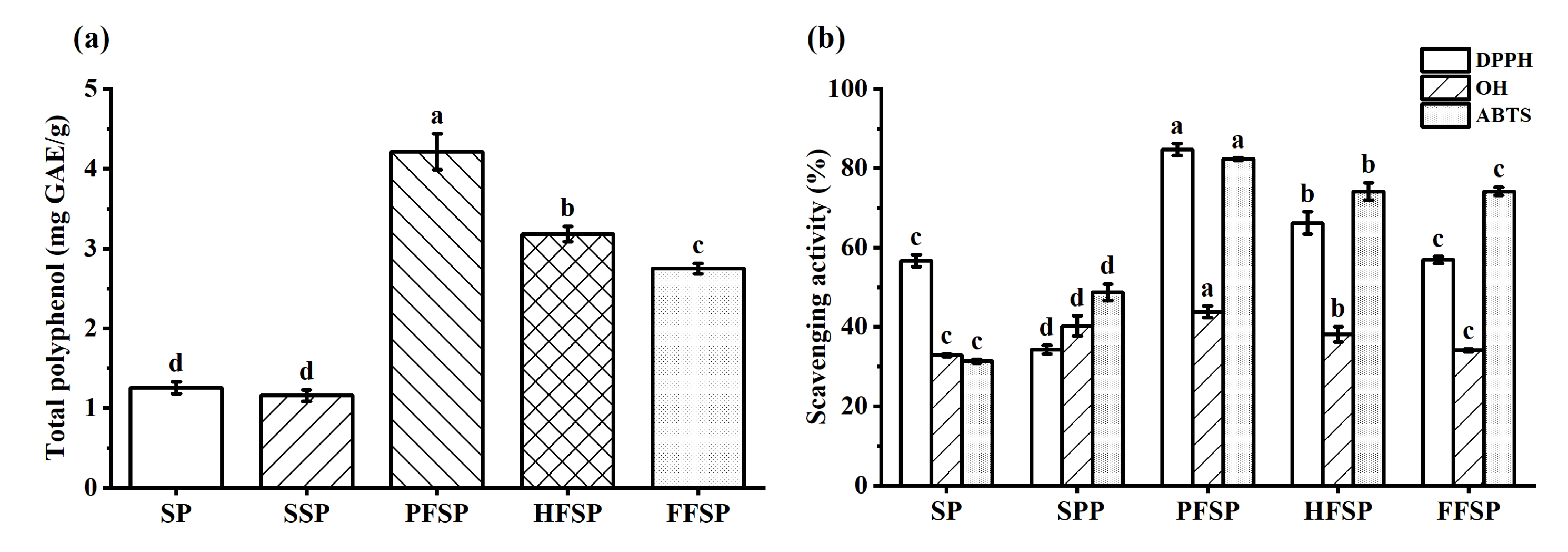
3.4. Total Polyphenol Content
3.5. Antioxidant Capacity
3.6. Color
3.7. Particle Size
3.8. SEM
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Teresa, B. Nutritional Value of Soybean Meal. In Soybean and Nutrition; Teresa, B., Ed.; IntechOpen: Rijeka, Croatia, 2011; p. 1. [Google Scholar]
- Silva, F.d.O.; Perrone, D. Characterization and stability of bioactive compounds from soybean meal. LWT Food Sci. Technol. 2015, 63, 992–1000. [Google Scholar] [CrossRef]
- Singh, P.; Krishnaswamy, K. Sustainable zero-waste processing system for soybeans and soy by-product valorization. Trends Food Sci. Technol. 2022, 128, 331–344. [Google Scholar] [CrossRef]
- Suruga, K.; Tomita, T.; Kadokura, K. Soybean fermentation with basidiomycetes (medicinal mushroom mycelia). Chem. Biol. Technol. Agric. 2020, 7, 23. [Google Scholar] [CrossRef]
- Hati, S.; Vij, S.; Singh, B.P.; Mandal, S. β-Glucosidase activity and bioconversion of isoflavones during fermentation of soymilk. J. Sci. Food Agric. 2015, 95, 216–220. [Google Scholar] [CrossRef]
- Mollakhalili-Meybodi, N.; Arab, M.; Zare, L. Harmful compounds of soy milk: Characterization and reduction strategies. J. Food Sci. Technol. 2022, 59, 3723–3732. [Google Scholar] [CrossRef]
- Jayachandran, M.; Xu, B. An insight into the health benefits of fermented soy products. Food Chem. 2019, 271, 362–371. [Google Scholar] [CrossRef]
- Toor, B.S.; Kaur, A.; Sahota, P.P.; Kaur, J. Antioxidant Potential, Antinutrients, Mineral Composition and FTIR Spectra of Legumes Fermented with Rhizopus oligosporus. Food Technol. Biotechnol. 2021, 59, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Du, B.; Xu, B. A systematic, comparative study on the beneficial health components and antioxidant activities of commercially fermented soy products marketed in China. Food Chem. 2015, 174, 202–213. [Google Scholar] [CrossRef]
- Roasa, J.; De Villa, R.; Mine, Y.; Tsao, R. Phenolics of cereal, pulse and oilseed processing by-products and potential effects of solid-state fermentation on their bioaccessibility, bioavailability and health benefits: A review. Trends Food Sci. Technol. 2021, 116, 954–974. [Google Scholar] [CrossRef]
- Olukomaiya, O.O.; Adiamo, O.Q.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour. Food Chem. 2020, 315, 126238. [Google Scholar] [CrossRef]
- Rathore, H.; Prasad, S.; Sharma, S. Mushroom nutraceuticals for improved nutrition and better human health: A review. PharmaNutrition 2017, 5, 35–46. [Google Scholar] [CrossRef]
- Xia, Y.D.; Wang, D.X.; Li, J.Q.; Chen, M.Q.; Wang, D.; Jiang, Z.P.; Liu, B. Compounds purified from edible fungi fight against chronic inflammation through oxidative stress regulation. Front. Pharmacol. 2022, 13, 13. [Google Scholar] [CrossRef]
- Gopal, J.; Sivanesan, I.; Muthu, M.; Oh, J.-W. Scrutinizing the Nutritional Aspects of Asian Mushrooms, Its Commercialization and Scope for Value-Added Products. Nutrients 2022, 14, 3700. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Li, H.-B.; Gunaratne, A.; Sui, Z.-Q.; Corke, H. Effects of Fermented Edible Seeds and Their Products on Human Health: Bioactive Components and Bioactivities. Compr. Rev. Food Sci. Food Saf. 2017, 16, 489–531. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Páez, E.; Alanis-Guzmán, M.G.; Hernández-Luna, C.E.; Báez-González, J.G.; Amaya-Guerra, C.A.; Andrés-Grau, A.M. Increasing Antioxidant Activity and Protein Digestibility in Phaseolus vulgaris and Avena sativa by Fermentation with the Pleurotus ostreatus Fungus. Molecules 2017, 22, 2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Yang, Y.; Guan, D.; Zhang, Y.; Zhang, Z. Bioactivity of the crude polysaccharides from fermented soybean curd residue by Flammulina velutipes. Carbohydr. Polym. 2012, 89, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of Fermentation in Improving Nutritional Quality of Soybean Meal—A Review. Asian-Australas J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asensio-Grau, A.; Calvo-Lerma, J.; Heredia, A.; Andrés, A. Enhancing the nutritional profile and digestibility of lentil flour by solid state fermentation with Pleurotus ostreatus. Food Funct. 2020, 11, 7905–7912. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Thakur, K.; Feng, J.Y.; Zhu, Y.Y.; Zhang, F.; Russo, P.; Spano, G.; Zhang, J.G.; Wei, Z.J. Functionalization of soy residue (okara) by enzymatic hydrolysis and LAB fermentation for B-2 bio-enrichment and improved in vitro digestion. Food Chem. 2022, 387, 10. [Google Scholar] [CrossRef]
- Ali, N.M.; Yeap, S.-K.; Yusof, H.M.; Beh, B.-K.; Ho, W.-Y.; Koh, S.-P.; Abdullah, M.P.; Alitheen, N.B.; Long, K. Comparison of free amino acids, antioxidants, soluble phenolic acids, cytotoxicity and immunomodulation of fermented mung bean and soybean. J. Sci. Food Agric. 2016, 96, 1648–1658. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-García, J.; Asensio-Grau, A.; García-Hernández, J.; Heredia, A.; Andrés, A. Nutritional and antioxidant changes in lentils and quinoa through fungal solid-state fermentation with Pleurotus ostreatus. Bioresour. Bioprocess. 2022, 9, 51. [Google Scholar] [CrossRef]
- Lee, H.Y.; Cho, D.Y.; Jang, K.J.; Lee, J.H.; Jung, J.G.; Kim, M.J.; Jeong, J.B.; Haque, M.A.; Cho, K.M. Changes of γ-Aminobutyric Acid, Phytoestrogens, and Biofunctional Properties of the Isoflavone-Enriched Soybean (Glycine max) Leaves during Solid Lactic Acid Fermentation. Fermentation 2022, 8, 525. [Google Scholar] [CrossRef]
- Li, S.; Chen, G.; Qiang, S.; Tang, D.; Chen, Y.; Zhang, Z.; Lei, Z.; Chen, Y. Intensifying soluble dietary fiber production and properties of soybean curd residue via autoclaving treatment. Bioresour. Technol. Rep. 2019, 7, 100203. [Google Scholar] [CrossRef]
- Salazar-Villanea, S.; Bruininx, E.M.A.M.; Butré, C.I.; van der Poel, A.F.B. Processing temperature and sugar type affect the rate and the extent of proteolysis of a model soy protein isolate system. Anim. Feed Sci. Technol. 2020, 269, 114680. [Google Scholar] [CrossRef]
- Lee, D.P.S.; Gan, A.X.; Kim, J.E. Incorporation of biovalorised okara in biscuits: Improvements of nutritional, antioxidant, physical, and sensory properties. LWT 2020, 134, 109902. [Google Scholar] [CrossRef]
- Nasehi, M.; Torbatinejad, N.M.; Zerehdaran, S.; Safaie, A.R. Effect of solid-state fermentation by oyster mushroom (Pleurotus florida) on nutritive value of some agro by-products. J. Appl. Anim. Res. 2017, 45, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Karimi, S.; Mahboobi Soofiani, N.; Lundh, T.; Mahboubi, A.; Kiessling, A.; Taherzadeh, M.J. Evaluation of Filamentous Fungal Biomass Cultivated on Vinasse as an Alternative Nutrient Source of Fish Feed: Protein, Lipid, and Mineral Composition. Fermentation 2019, 5, 99. [Google Scholar] [CrossRef] [Green Version]
- Vong, W.C.; Au Yang, K.L.C.; Liu, S.-Q. Okara (soybean residue) biotransformation by yeast Yarrowia lipolytica. Int. J. Food Microbiol. 2016, 235, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.; Patel, A.S.; Kar, A.; Deshpande, S.S.; Tripathi, M.K. Effect of different processing conditions on proximate composition, anti-oxidants, anti-nutrients and amino acid profile of grain sorghum. Food Chem. 2019, 271, 129–135. [Google Scholar] [CrossRef]
- Pranoto, Y.; Anggrahini, S.; Efendi, Z. Effect of natural and Lactobacillus plantarum fermentation on in-vitro protein and starch digestibilities of sorghum flour. Food Biosci. 2013, 2, 46–52. [Google Scholar] [CrossRef]
- Zhao, Z.; Yang, R.; Gu, Z. Enzymes produced by liquid state fermentation of edible fungi and their effects on the main nutrients of soybean okara. Food Ferment. Ind. 2016, 42, 108–113. [Google Scholar] [CrossRef]
- Qu, S.; Kwon, S.J.; Duan, S.; Lim, Y.J.; Eom, S.H. Isoflavone Changes in Immature and Mature Soybeans by Thermal Processing. Molecules 2021, 26, 7471. [Google Scholar] [CrossRef]
- Leksono, B.Y.; Cahyanto, M.N.; Rahayu, E.S.; Yanti, R.; Utami, T. Enhancement of Antioxidant Activities in Black Soy Milk through Isoflavone Aglycone Production during Indigenous Lactic Acid Bacteria Fermentation. Fermentation 2022, 8, 326. [Google Scholar] [CrossRef]
- Yoon, L.W.; Ang, T.N.; Ngoh, G.C.; Chua, A.S.M. Fungal solid-state fermentation and various methods of enhancement in cellulase production. Biomass Bioenergy 2014, 67, 319–338. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, Y.; Dong, L.; Jia, X.; Liu, L.; Huang, F.; Chi, J.; Xiao, J.; Zhang, M.; Zhang, R. Extrusion and fungal fermentation change the profile and antioxidant activity of free and bound phenolics in rice bran together with the phenolic bioaccessibility. LWT 2019, 115, 108461. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Torino, M.I.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Dhull, S.B.; Punia, S.; Kidwai, M.K.; Kaur, M.; Chawla, P.; Purewal, S.S.; Sangwan, M.; Palthania, S. Solid-state fermentation of lentil (Lens culinaris L.) with Aspergillus awamori: Effect on phenolic compounds, mineral content, and their bioavailability. Legume Sci. 2020, 2, e37. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Hayat, I.; Arif, S.; Masud, T.; Khalid, N.; Ahmed, A. Mechanisms Involved in the Therapeutic Effects of Soybean (Glycine Max). Int. J. Food Prop. 2014, 17, 1332–1354. [Google Scholar] [CrossRef] [Green Version]
- Verni, M.; De Mastro, G.; De Cillis, F.; Gobbetti, M.; Rizzello, C.G. Lactic acid bacteria fermentation to exploit the nutritional potential of Mediterranean faba bean local biotypes. Food Res. Int. 2019, 125, 108571. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Páez, E.; Hernández-Luna, C.E.; Longoria-García, S.; Martínez-Silva, P.A.; Ortiz-Rodríguez, I.; Villarreal-Vera, M.T.; Cantú-Saldaña, C.M. Pleurotus ostreatus: A potential concurrent biotransformation agent/ingredient on development of functional foods (cookies). LWT 2021, 148, 111727. [Google Scholar] [CrossRef]
- Toor, B.S.; Kaur, A.; Kaur, J. Fermentation of legumes with Rhizopus oligosporus: Effect on physicochemical, functional and microstructural properties. Int. J. Food Sci. Technol. 2022, 57, 1763–1772. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Crude Protein (%) | Sugars (%) | Moisture (%) | Ashes (%) |
|---|---|---|---|---|
| SP | 48.39 ± 3.46 b | 27.15 ± 2.07 a | 11.41 ± 0.42 a | 6.39 ± 0.28 a |
| SSP | 38.42 ± 3.29 c | 21.92 ± 1.86 b | 7.38 ± 0.35 c | 5.87 ± 0.31 b |
| PFSP | 52.98 ± 3.87 a | 17.98 ± 1.73 c | 7.30 ± 0.38 c | 5.44 ± 0.24 c |
| HFSP | 51.95 ± 3.14 a | 20.67 ± 1.47 b | 7.55 ± 0.29 b | 5.74 ± 0.43 b |
| FFSP | 51.18 ± 3.55 a | 18.86 ± 1.62 c | 7.46 ± 0.31 b | 5.49 ± 0.37 c |
| Color Coordinates | L* | a* | b* | ∆E |
|---|---|---|---|---|
| SP | 84.33 ± 2.27 a | 0.02 ± 0.20 e | 14.32 ± 0.40 c | — |
| SSP | 50.98 ± 1.54 c | 10.45 ± 0.74 b | 16.83 ± 0.92 b | 35.06 ± 1.18 a |
| PFSP | 57.68 ± 0.45 b | 11.36 ± 0.35 a | 21.08 ± 0.60 a | 29.75 ± 0.36 b |
| HFSP | 51.72 ± 1.01 c | 8.45 ± 0.01 c | 15.13 ± 0.67 c | 33.70 ± 0.96 a |
| FFSP | 57.85 ± 1.03 b | 7.37 ± 0.55 d | 15.17 ± 0.66 c | 27.51 ± 0.87 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Jiang, Q.; Huang, Z.; Wang, Y.; Roubik, H.; Yang, K.; Cai, M.; Sun, P. Solid-State Fermentation of Soybean Meal with Edible Mushroom Mycelium to Improve Its Nutritional, Antioxidant Capacities and Physicochemical Properties. Fermentation 2023, 9, 322. https://doi.org/10.3390/fermentation9040322
Wang J, Jiang Q, Huang Z, Wang Y, Roubik H, Yang K, Cai M, Sun P. Solid-State Fermentation of Soybean Meal with Edible Mushroom Mycelium to Improve Its Nutritional, Antioxidant Capacities and Physicochemical Properties. Fermentation. 2023; 9(4):322. https://doi.org/10.3390/fermentation9040322
Chicago/Turabian StyleWang, Jian, Quanjin Jiang, Zhenyu Huang, Yan Wang, Hynek Roubik, Kai Yang, Ming Cai, and Peilong Sun. 2023. "Solid-State Fermentation of Soybean Meal with Edible Mushroom Mycelium to Improve Its Nutritional, Antioxidant Capacities and Physicochemical Properties" Fermentation 9, no. 4: 322. https://doi.org/10.3390/fermentation9040322
APA StyleWang, J., Jiang, Q., Huang, Z., Wang, Y., Roubik, H., Yang, K., Cai, M., & Sun, P. (2023). Solid-State Fermentation of Soybean Meal with Edible Mushroom Mycelium to Improve Its Nutritional, Antioxidant Capacities and Physicochemical Properties. Fermentation, 9(4), 322. https://doi.org/10.3390/fermentation9040322