
Fermentation of Corn By-Products: From Agrifood Waste to Higher Value Antioxidant Products

 , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Corn By-Products Properties
2.2. Corn By-Products Fermentation
2.3. Corn By-Products Hydroalcoholic Extract
2.4. Determination of Antioxidant Activity
2.4.1. Determination of Total Phenol Content
2.4.2. 2-2 Azinobis(3-Ethylbenzothiazoline-6-sulfonic acid) (ABTS) Scavenging Assay
2.4.3. 2,2-difenil-1-picrylidrazyl (DPPH) Scavenging Assay
2.4.4. Lipid Peroxidation Assay
2.5. LC-DAD-ESI/MS Measurements
2.6. Statistical Analysis
3. Results and Discussion
3.1. Fermentation
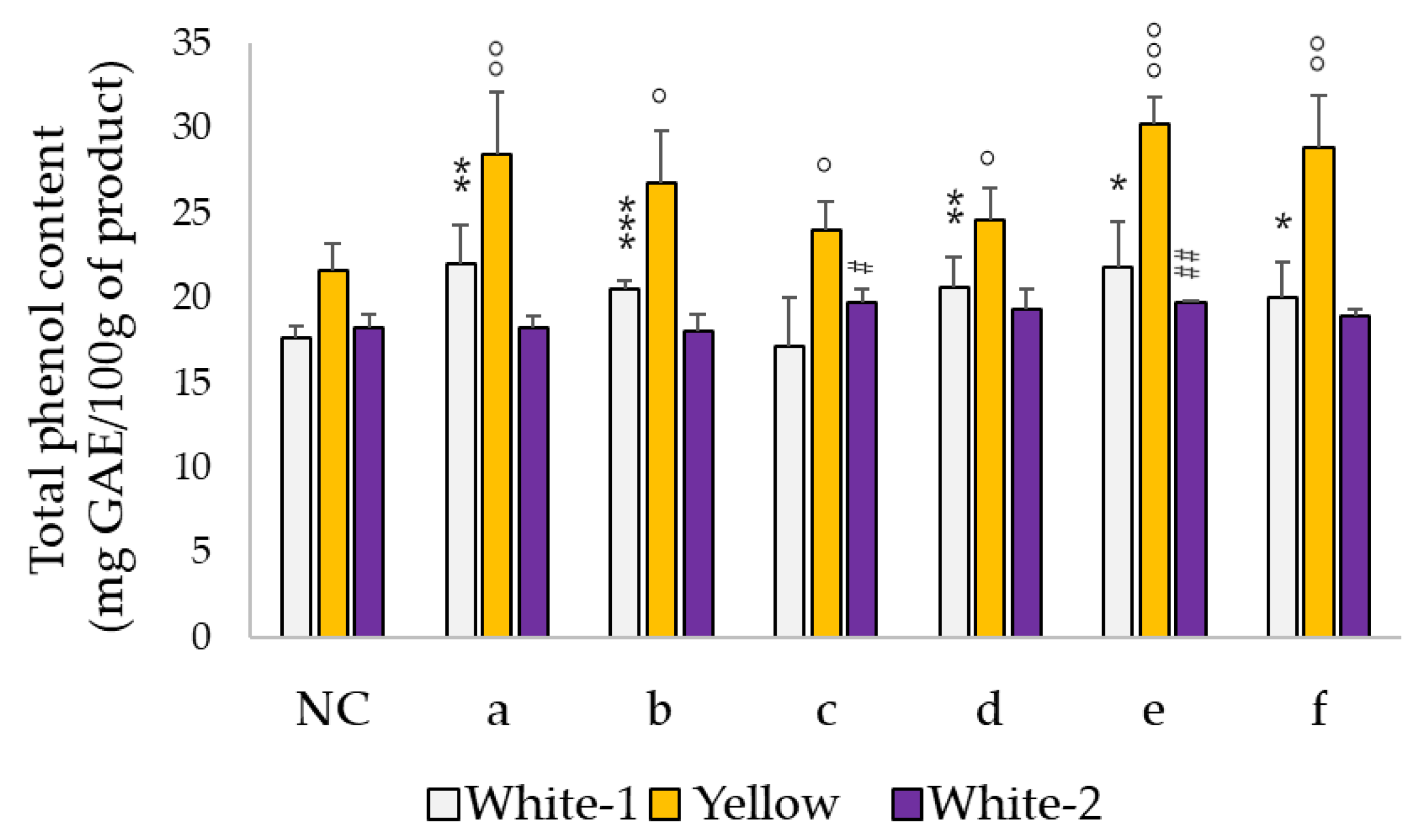
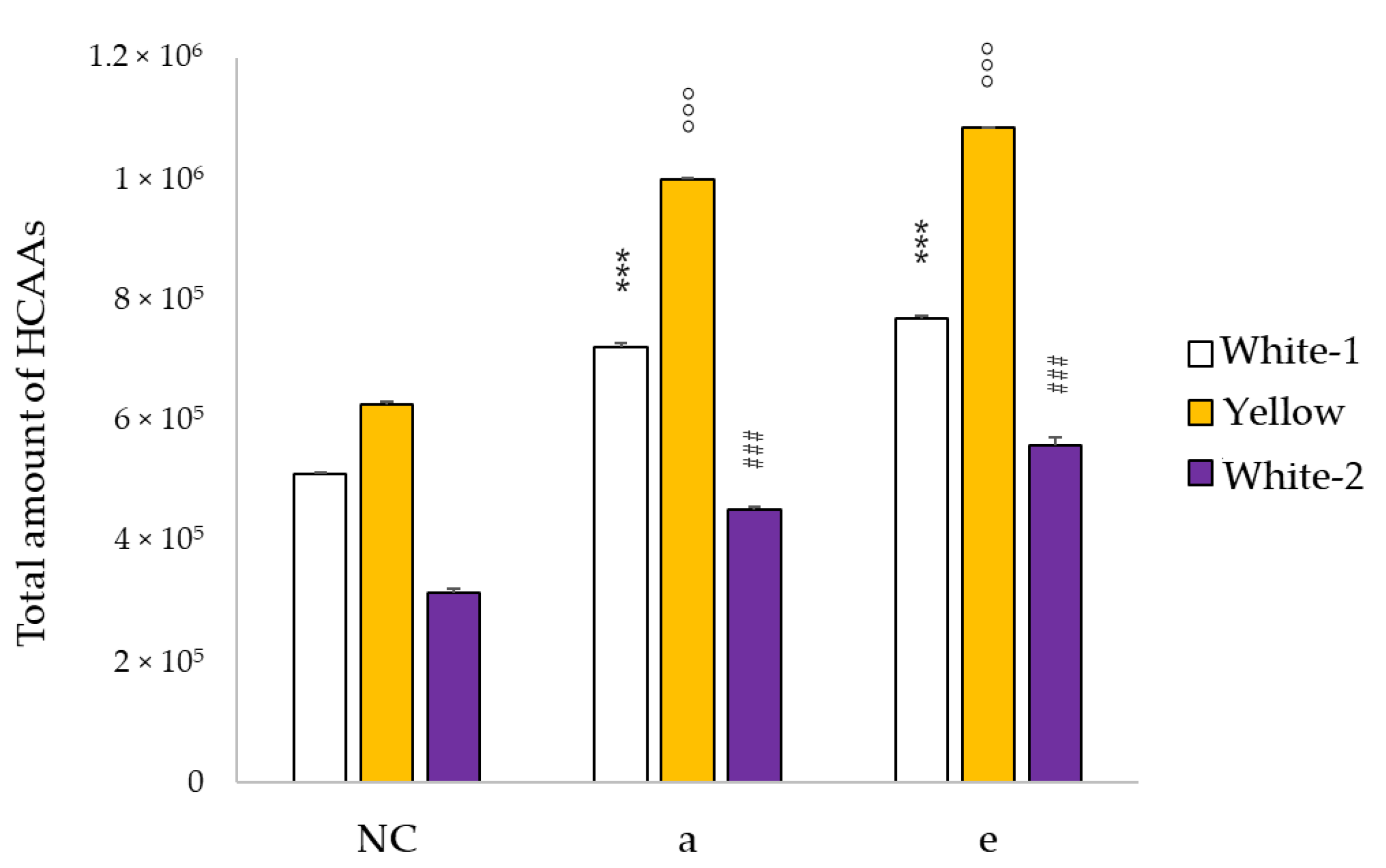
3.2. Total Phenol Content Increases after Fermentation
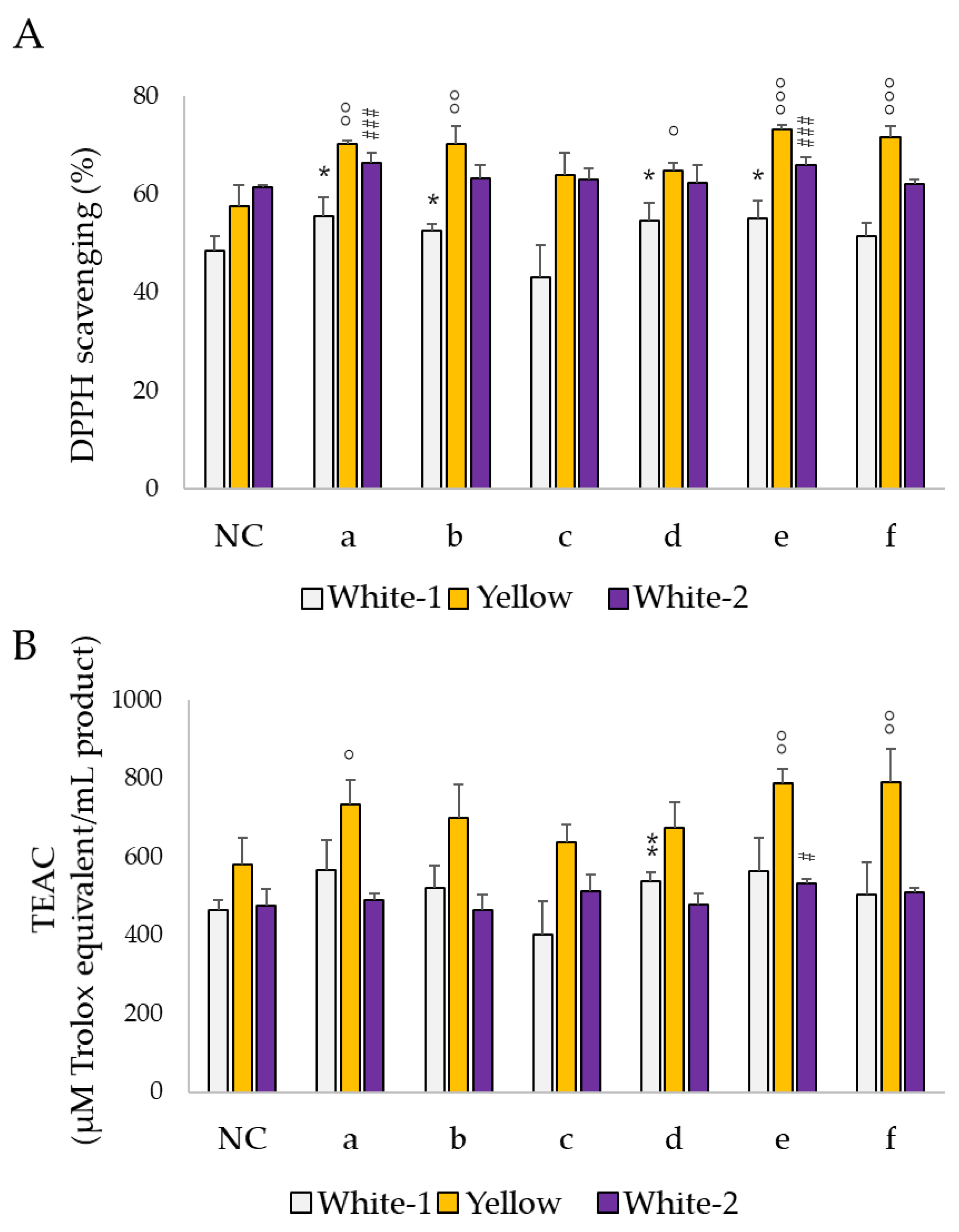
3.3. ABTS and DPPH
3.4. TBARS Determination in Soy Beverage in the Presence of Different Corn by-Product Extracts
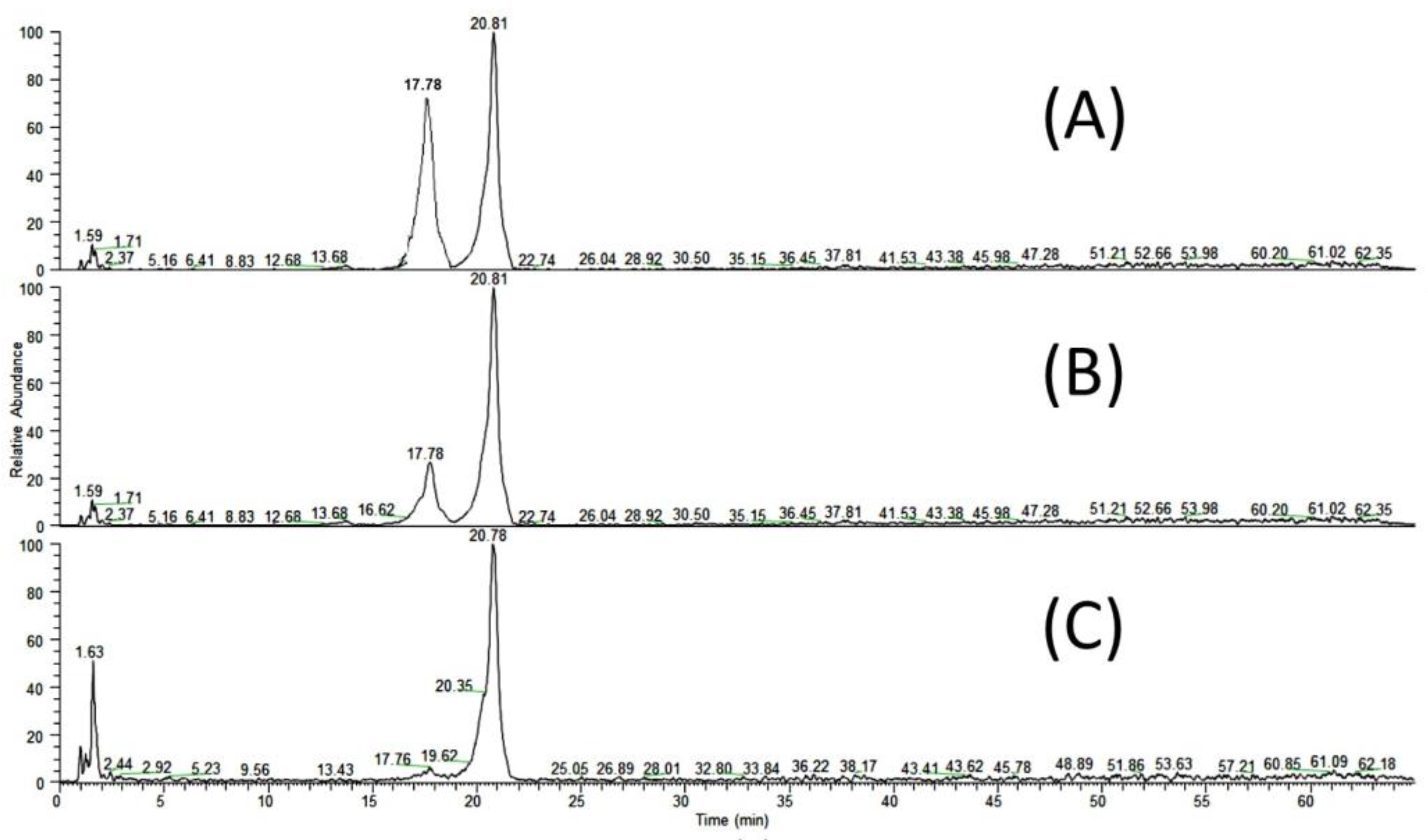
3.5. LC-DAD-ESI/MS Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamal, H.; Le, C.F.; Salter, A.M.; Ali, A. Extraction of Protein from Food Waste: An Overview of Current Status and Opportunities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2455–2475. [Google Scholar] [CrossRef]
- Transforming Our World: The 2030 Agenda for Sustainable Development|Department of Economic and Social Affairs. Available online: https://sdgs.un.org/2030agenda (accessed on 8 March 2023).
- Sancho, S.d.O.; da Silva, A.R.A.; Dantas, A.N.d.S.; Magalhães, T.A.; Lopes, G.S.; Rodrigues, S.; da Costa, J.M.C.; Fernandes, F.A.N.; Silva, M.G.d.V. Characterization of the Industrial Residues of Seven Fruits and Prospection of Their Potential Application as Food Supplements. J. Chem. 2015, 2015, e264284. [Google Scholar] [CrossRef] [Green Version]
- Torres-León, C.; Ramírez-Guzman, N.; Londoño-Hernandez, L.; Martinez-Medina, G.A.; Díaz-Herrera, R.; Navarro-Macias, V.; Alvarez-Pérez, O.B.; Picazo, B.; Villarreal-Vázquez, M.; Ascacio-Valdes, J.; et al. Food Waste and Byproducts: An Opportunity to Minimize Malnutrition and Hunger in Developing Countries. Front. Sustain. Food Syst. 2018, 2, 52. [Google Scholar] [CrossRef]
- Zhang, R.; Ma, S.; Li, L.; Zhang, M.; Tian, S.; Wang, D.; Liu, K.; Liu, H.; Zhu, W.; Wang, X. Comprehensive Utilization of Corn Starch Processing By-Products: A Review. Grain Oil Sci. Technol. 2021, 4, 89–107. [Google Scholar] [CrossRef]
- Pontonio, E.; Dingeo, C.; Gobbetti, M.; Rizzello, C.G. Maize Milling By-Products: From Food Wastes to Functional Ingredients Through Lactic Acid Bacteria Fermentation. Front. Microbiol. 2019, 10, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwirtz, J.A.; Garcia-Casal, M.N. Processing Maize Flour and Corn Meal Food Products. Ann. N. Y. Acad. Sci. 2014, 1312, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Siyuan, S.; Tong, L.; Liu, R. Corn Phytochemicals and Their Health Benefits. Food Sci. Hum. Wellness 2018, 7, 185–195. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented Foods as a Dietary Source of Live Organisms. Front Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health Benefits of Fermented Foods: Microbiota and Beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Sáez, G.D.; Sabater, C.; Fara, A.; Zárate, G. Fermentation of Chickpea Flour with Selected Lactic Acid Bacteria for Improving Its Nutritional and Functional Properties. J. Appl. Microbiol. 2022, 133, 181–199. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Variation of Microbial Community and Fermentation Quality in Corn Silage Treated with Lactic Acid Bacteria and Artemisia Argyi during Aerobic Exposure. Toxins 2022, 14, 349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Miao, F.; Wang, X.; Lu, W.; Ma, C. Effects of Homo- and Hetero-Fermentative Lactic Acid Bacteria on the Quality and Aerobic Stability of Corn Silage. Can. J. Anim. Sci. 2021, 101, 761–770. [Google Scholar] [CrossRef]
- Andrighetto, C.; Borney, F.; Barmaz, A.; Stefanon, B.; Lombardi, A. Genetic Diversity of Streptococcus Thermophilus Strains Isolated from Italian Traditional Cheeses. Int. Dairy J. 2002, 12, 141–144. [Google Scholar] [CrossRef]
- Tonolo, F.; Moretto, L.; Folda, A.; Scalcon, V.; Bindoli, A.; Bellamio, M.; Feller, E.; Rigobello, M.P. Antioxidant Properties of Fermented Soy during Shelf Life. Plant Foods Hum. Nutr. 2019, 74, 287–292. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. [30] Microsomal Lipid Peroxidation. In Methods in Enzymology; Fleischer, S., Packer, L., Eds.; Biomembranes—Part C: Biological Oxidations; Academic Press: Cambridge, MA, USA, 1978; Volume 52, pp. 302–310. [Google Scholar]
- Gänzle, M.; Ripari, V. Composition and Function of Sourdough Microbiota: From Ecological Theory to Bread Quality. Int. J. Food Microbiol. 2016, 239, 19–25. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic Acid Bacteria from Fermented Table Olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef]
- Beganović, J.; Kos, B.; Leboš Pavunc, A.; Uroić, K.; Jokić, M.; Šušković, J. Traditionally Produced Sauerkraut as Source of Autochthonous Functional Starter Cultures. Microbiol. Res. 2014, 169, 623–632. [Google Scholar] [CrossRef]
- Bento-Silva, A.; Duarte, N.; Mecha, E.; Belo, M.; Vaz Patto, M.C.; Bronze, M. do R. Hydroxycinnamic Acids and Their Derivatives in Broa, a Traditional Ethnic Maize Bread. Foods 2020, 9, 1471. [Google Scholar] [CrossRef]
- del Río, J.C.; Rencoret, J.; Gutiérrez, A.; Kim, H.; Ralph, J. Structural Characterization of Lignin from Maize (Zea Mays L.) Fibers: Evidence for Diferuloylputrescine Incorporated into the Lignin Polymer in Maize Kernels. J. Agric. Food Chem. 2018, 66, 4402–4413. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.W.; Lee, S.K.; Kim, E.O.; Oh, J.H.; Yoon, K.S.; Parris, N.; Hicks, K.B.; Moreau, R.A. Antioxidant and Antimelanogenic Activities of Polyamine Conjugates from Corn Bran and Related Hydroxycinnamic Acids. J. Agric. Food Chem. 2007, 55, 3920–3925. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Serna-Saldívar, S.O.; Gutiérrez-Uribe, J.A. Chemopreventive Effects of Feruloyl Putrescines from Wastewater (Nejayote) of Lime-Cooked White Maize (Zea Mays). J. Cereal Sci. 2015, 64, 23–28. [Google Scholar] [CrossRef]
- Kim, E.-O.; Kwon, T.-K.; Choi, S.-W. Diferuloylputrescine, a Predominant Phenolic Amide in Corn Bran, Potently Induces Apoptosis in Human Leukemia U937 Cells. J. Med. Food 2014, 17, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Burt, A.J.; Arnason, J.T.; García-Lara, S. Natural Variation of Hydroxycinnamic Acid Amides in Maize Landraces. J. Cereal Sci. 2019, 88, 145–149. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Fermenting Strains |
|---|---|
| a | Lactiplantibacillus plantarum TH925 |
| b | Lacticaseibacillus rhamnosus TH414 |
| c | Lactobacillus amylovorus TH875 |
| d | Lactococcus lactis TH928 |
| e | Pediococcus pentosaceus TH967/968 * |
| f | Leuconostoc citreum TH931 |
| Corn By-Product | Name | Fermenting Strains | pH at the End | Min at pH 4.60 | Colony Count (Log CFU/g) | % Colonization |
|---|---|---|---|---|---|---|
| White-1 | a | Lactiplantibacillus plantarum | 3.47 | 304 | 9.59 | 100% |
| b | Lacticaseibacillus rhamnosus | 3.91 | 753 | 9.23 | 78% | |
| c | Lactobacillus amylovorus | 4.05 | 48 | 8.74 | 45% | |
| d | Lactococcus lactis | 3.86 | 165 | 8.82 | 100% | |
| e | Pediococcus pentosaceus | 3.66 | 480 | 8.57 | 87% | |
| f | Leuconostoc citreum | 3.87 | 236 | 8.93 | 86% | |
| Yellow | a | Lactiplantibacillus plantarum | 3.79 | 462 | 9.57 | 100% |
| b | Lacticaseibacillus rhamnosus | 4.05 | 787 | 9.41 | 100% | |
| c | Lactobacillus amylovorus | 4.04 | 229 | 8.97 | 29% | |
| d | Lactococcus lactis | 4.12 | 274 | 9.11 | 100% | |
| e | Pediococcus pentosaceus | 3.96 | 481 | 9.20 | 100% | |
| f | Leuconostoc citreum | 4.12 | 370 | 9.15 | 94% | |
| White-2 | a | Lactiplantibacillus plantarum | 3.75 | 555 | 10.23 | 100% |
| b | Lacticaseibacillus rhamnosus | 3.98 | 771 | 9.62 | 100% | |
| c | Lactobacillus amylovorus | 3.84 | 216 | 9.08 | 33% | |
| d | Lactococcus lactis | 4.09 | 242 | 9.23 | 100% | |
| e | Pediococcus pentosaceus | 3.84 | 463 | 9.41 | 100% | |
| f | Leuconostoc citreum | 4.05 | 344 | 9.60 | 100% |
| Sample | White-1 | Yellow | White-2 |
|---|---|---|---|
| NC | 79.66 ± 2.94 | 62.61 ± 6.59 | 79.66 ± 2.90 |
| a | 80.04 ± 3.32 | 59.99 ± 8.49 | 80.70 ± 6.54 |
| b | 80.31 ± 1.28 | 64.62 ± 5.89 | 64.46 ± 7.84 |
| c | 80.42 ± 1.88 | 69.09 ± 3.80 | 86.25 ± 2.78 |
| d | 80.10 ± 3.06 | 77.86 ± 7.36 | 73.18 ± 6.56 |
| e | 81.13 ± 3.74 | 63.80 ± 4.36 | 63.37 ± 5.56 |
| f | 80.53 ± 1.16 | 69.53 ± 6.64 | 59.77 ± 6.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonolo, F.; Folda, A.; Ferro, S.; Seraglia, R.; Lombardi, A.; Andrighetto, C.; Giannoni, A.; Marin, O.; Rigobello, M.P. Fermentation of Corn By-Products: From Agrifood Waste to Higher Value Antioxidant Products. Fermentation 2023, 9, 373. https://doi.org/10.3390/fermentation9040373
Tonolo F, Folda A, Ferro S, Seraglia R, Lombardi A, Andrighetto C, Giannoni A, Marin O, Rigobello MP. Fermentation of Corn By-Products: From Agrifood Waste to Higher Value Antioxidant Products. Fermentation. 2023; 9(4):373. https://doi.org/10.3390/fermentation9040373
Chicago/Turabian StyleTonolo, Federica, Alessandra Folda, Stefania Ferro, Roberta Seraglia, Angiolella Lombardi, Christian Andrighetto, Alessia Giannoni, Oriano Marin, and Maria Pia Rigobello. 2023. "Fermentation of Corn By-Products: From Agrifood Waste to Higher Value Antioxidant Products" Fermentation 9, no. 4: 373. https://doi.org/10.3390/fermentation9040373
APA StyleTonolo, F., Folda, A., Ferro, S., Seraglia, R., Lombardi, A., Andrighetto, C., Giannoni, A., Marin, O., & Rigobello, M. P. (2023). Fermentation of Corn By-Products: From Agrifood Waste to Higher Value Antioxidant Products. Fermentation, 9(4), 373. https://doi.org/10.3390/fermentation9040373