
Understanding the Essential Metabolic Nodes in the Synthesis of 4-Acetylantroquinol B (4-AAQB) by Antrodia cinnamomea Using Transcriptomic Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Medium
2.2. Culture Methods
2.3. Analysis Methods
2.3.1. Dry Cell Weight of A. cinnamomea and 4-AAQB Detection Method
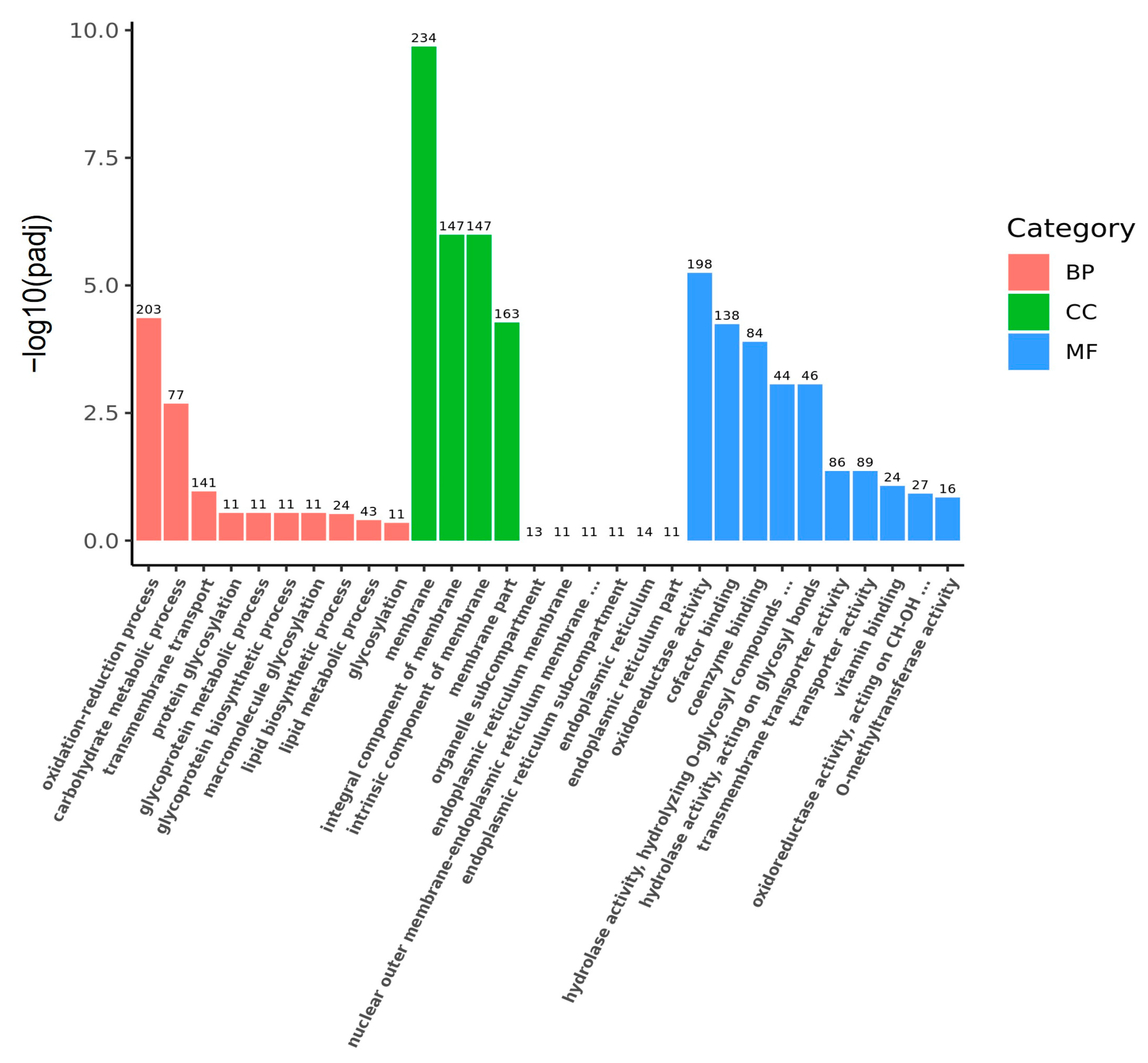
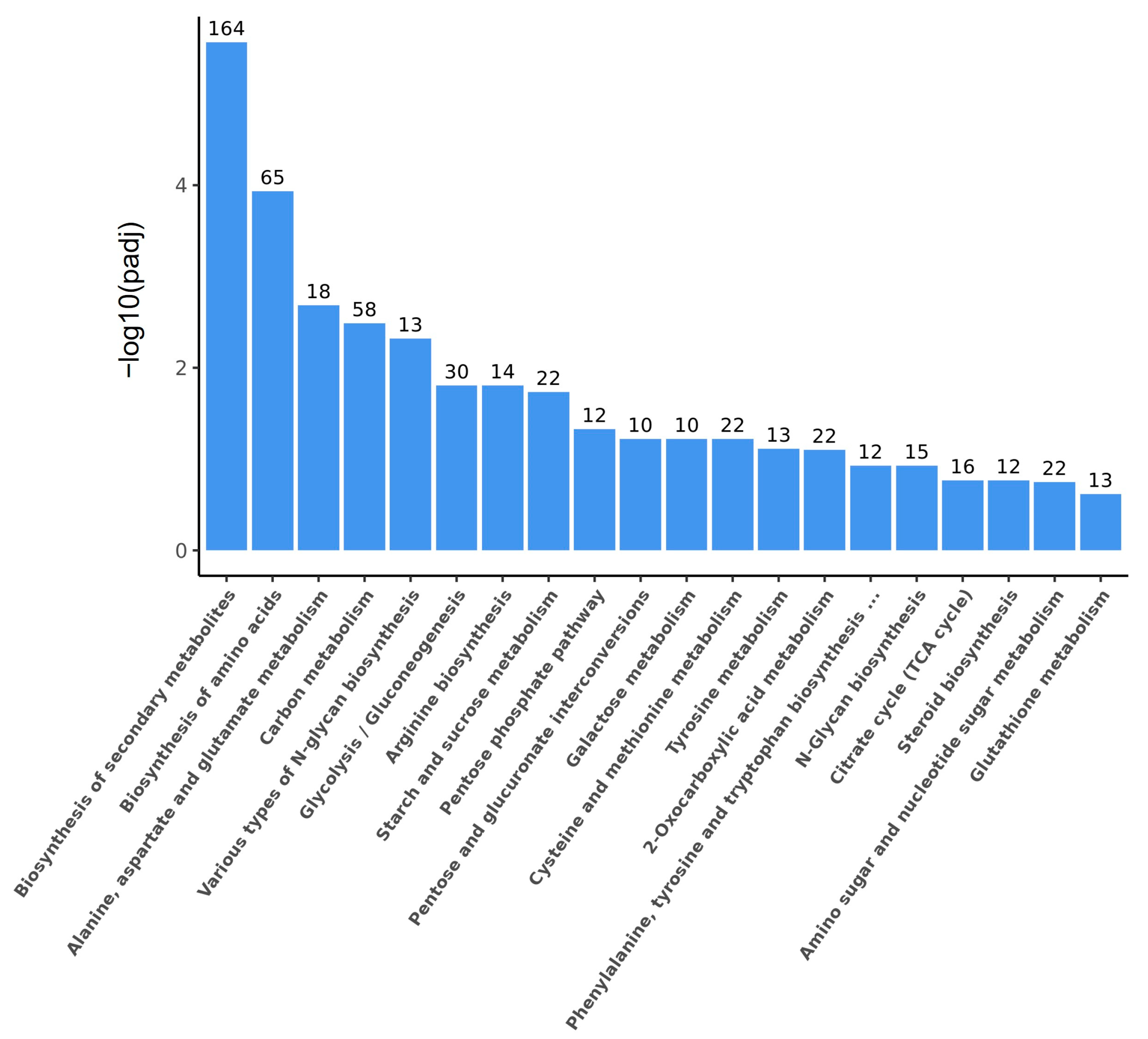
2.3.2. Transcriptome Analysis
2.3.3. Quantitative PCR (qPCR) Analysis
3. Results and Discussion
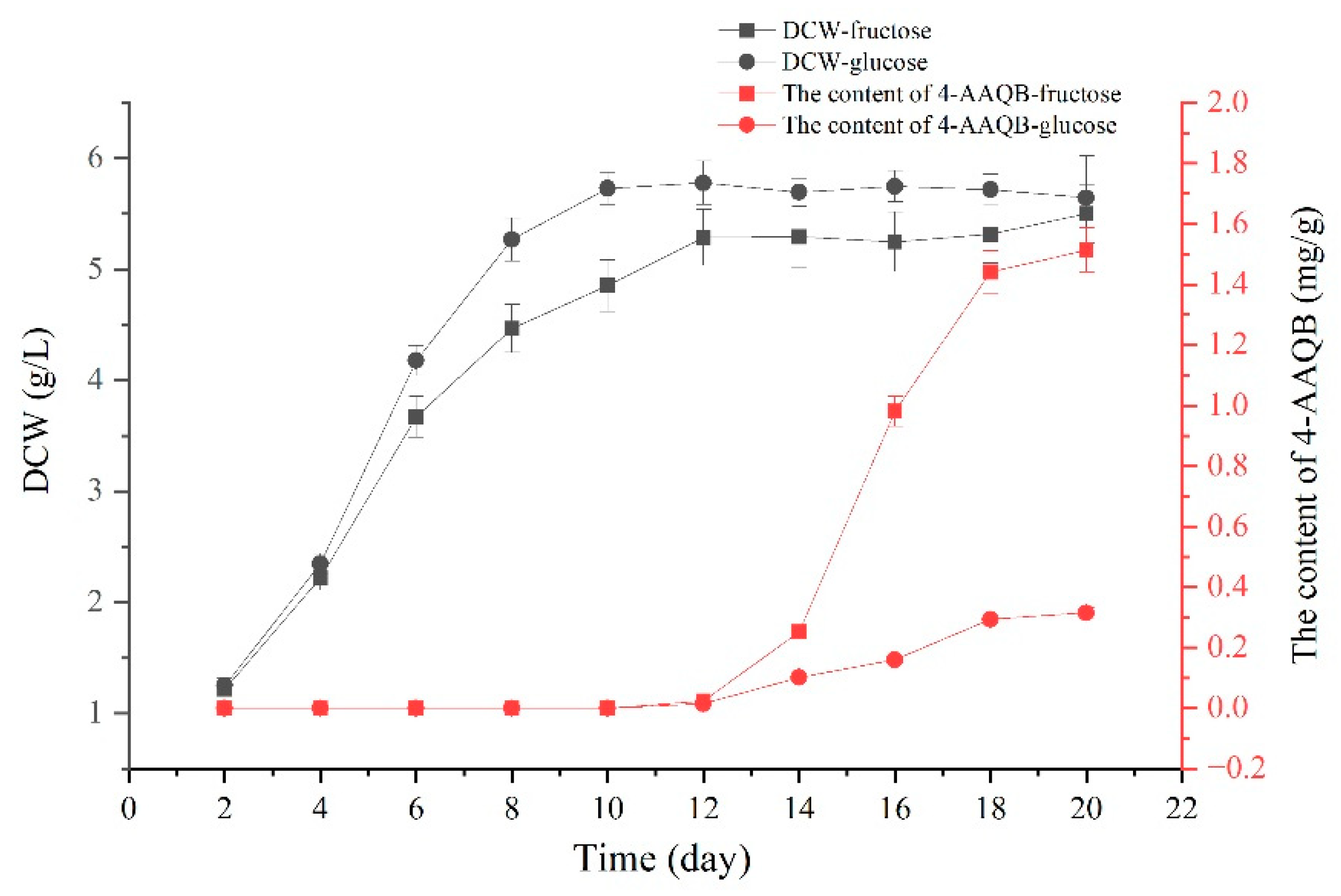
3.1. Comparison of 4-AAQB Production from Fructose and Glucose
3.2. Transcriptomic Analysis of 4-AAQB Synthesis Using Glucose and Fructose
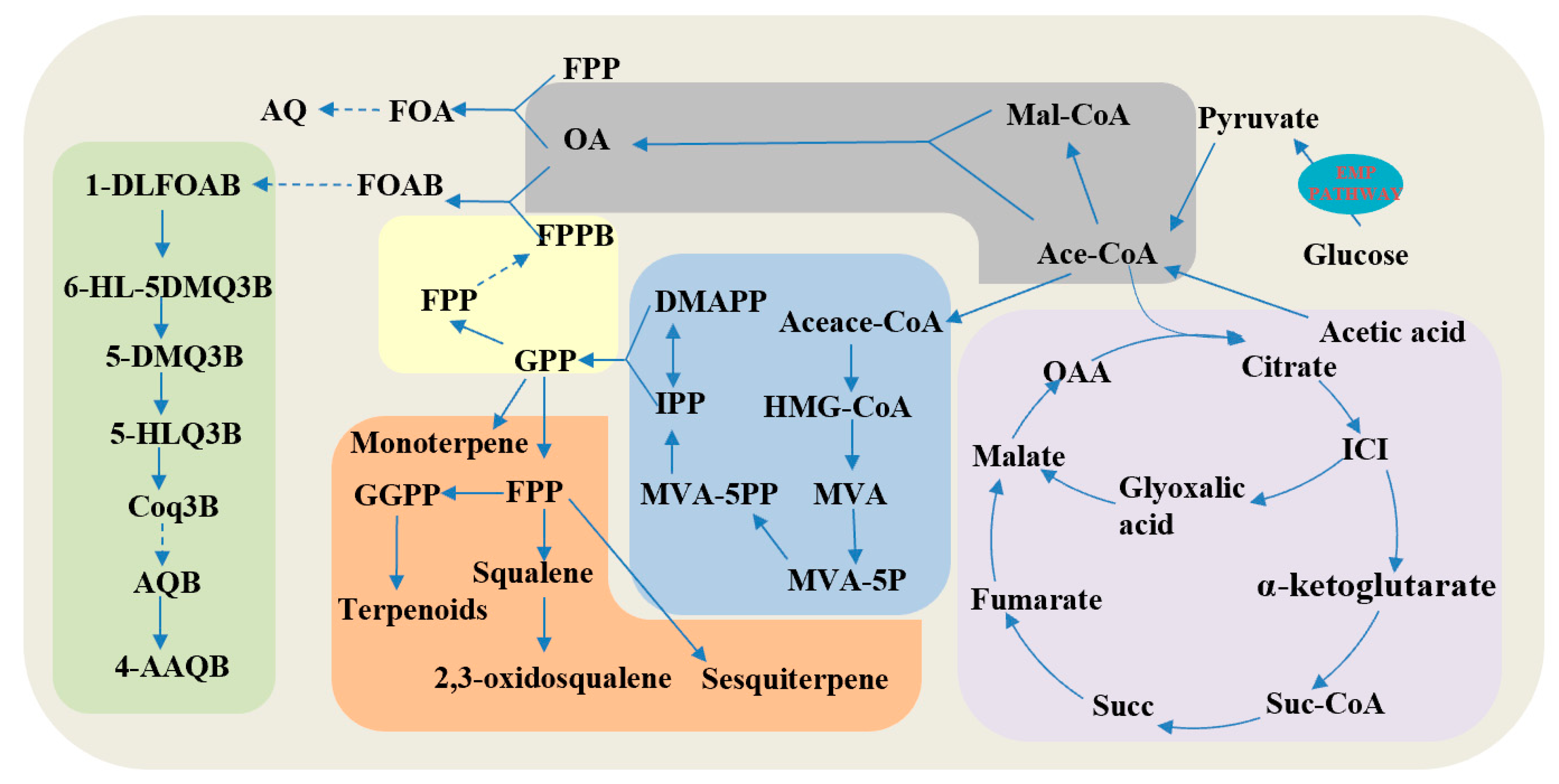
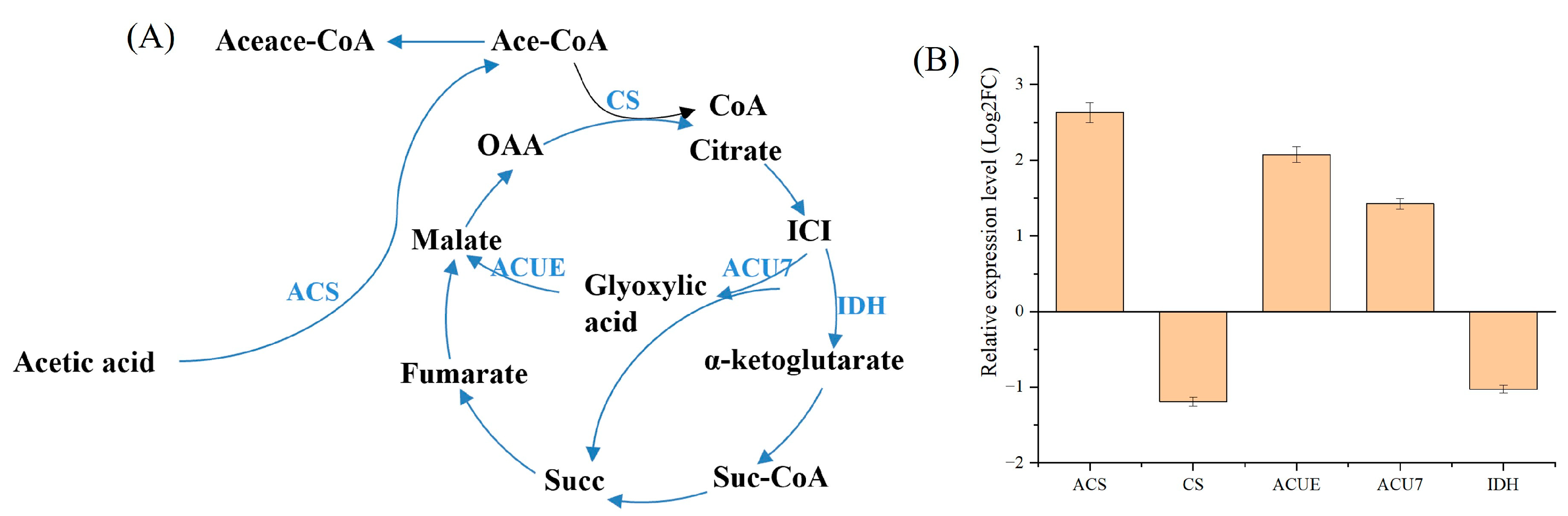
3.2.1. Analysis of the TCA Cycle and Glyoxylate Pathway Module
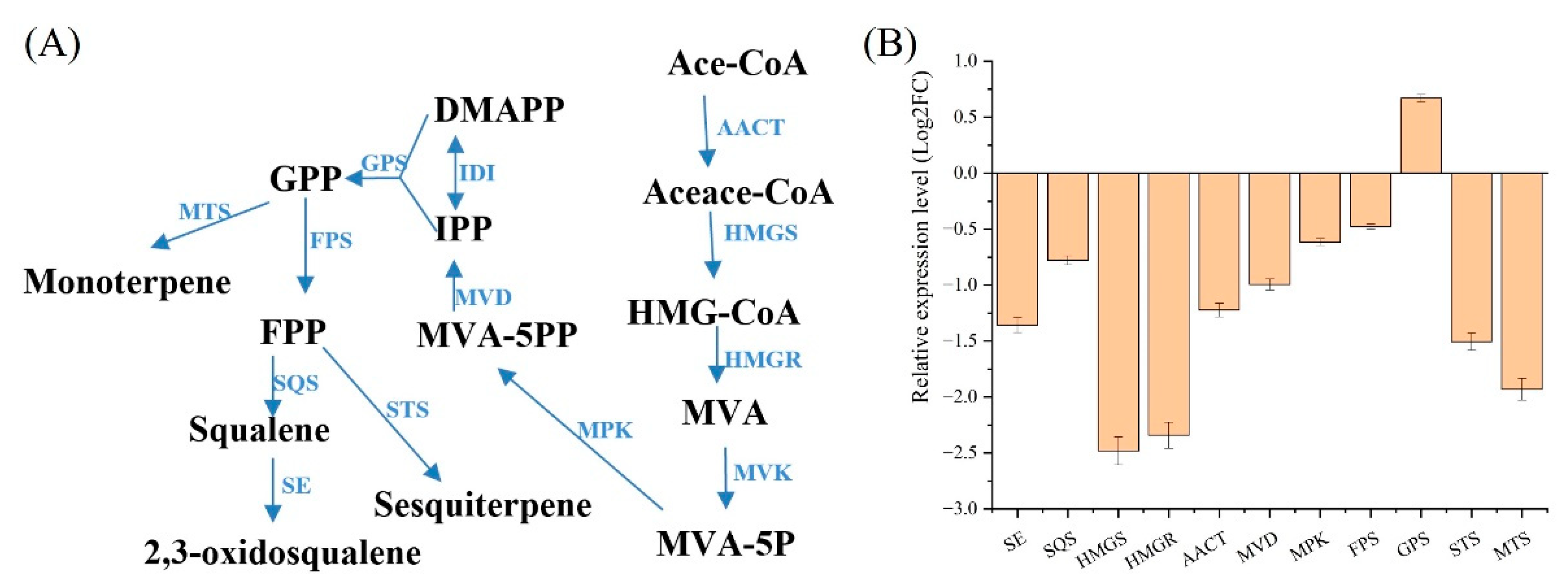
3.2.2. Analysis of Terpene Synthesis Module
3.2.3. Analysis of the Quinone Ring Modification Module
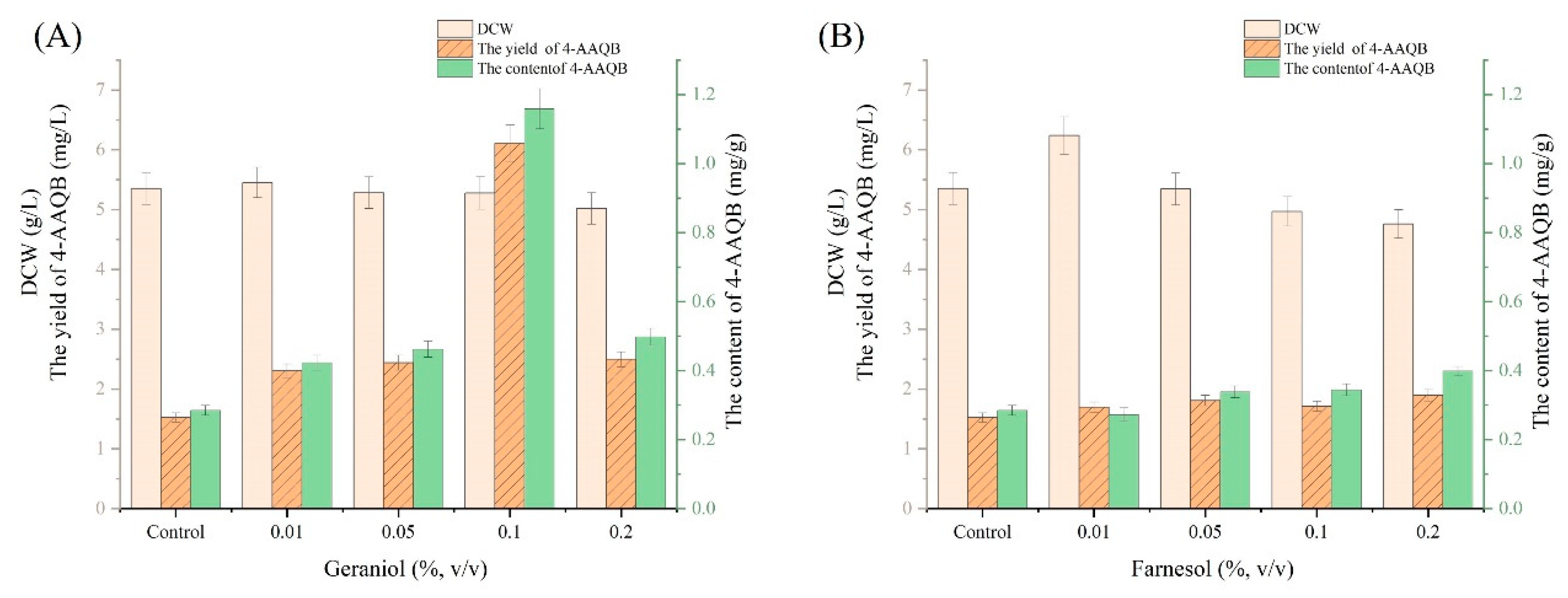
3.3. Effects of Geraniol and Farnesol on the Production of 4-AAQB
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, S.S.; Wang, G.J.; Wang, S.Y.; Lin, Y.Y.; Kuo, Y.H.; Lee, T.H. New Constituents with INOS Inhibitory Activity from Mycelium of Antrodia camphorata. Planta Med. 2009, 75, 512–516. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.W.; Pan, J.H.; Liu, R.H.; Kuo, Y.H.; Sheen, L.Y.; Chiang, B.H. The 4-Acetylantroquinonol B Isolated from Mycelium of Antrodia cinnamomea Inhibits Proliferation of Hepatoma Cells. J. Sci. Food Agric. 2010, 90, 1739–1744. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Ou, C.H.; Yen, I.C.; Lee, S.Y. 4-Acetylantroquinonol B Inhibits Osteoclastogenesis by Inhibiting the Autophagy Pathway in a Simulated Microgravity Model. Int. J. Mol. Sci. 2020, 21, 6971. [Google Scholar] [CrossRef]
- Chang, C.H.; Hsu, C.C.; Lee, A.S.; Wang, S.W.; Lin, K.T.; Chang, W.L.; Peng, H.C.; Huang, W.C.; Chung, C.H. 4-Acetylantroquinonol B Inhibits Lipopolysaccharide-Induced Cytokine Release and Alleviates Sepsis through of MAPK and NFκB Suppression. BMC Complem. Altern. Med. 2018, 18, 108. [Google Scholar] [CrossRef] [Green Version]
- Yen, I.C.; Tu, Q.W.; Chang, T.C.; Lin, P.H.; Li, Y.F.; Lee, S.Y. 4-Acetylantroquinonol B Ameliorates Nonalcoholic Steatohepatitis by Suppression of ER Stress and NLRP3 Inflammasome Activation. Biomed. Pharmacother. 2021, 138, 111504. [Google Scholar] [CrossRef]
- Lin, T.C.; Alison, G.; Liu, Z. Influences of Antroquinonol and 4-Acetylantroquinonol B on Inflammatory Tumorigenesis in the MCF-7 Breast Cancer Cell Line. Curr. Dev. Nutr. 2019, 3, nzz031.P06-039-19. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.C.; Yeh, C.T.; Adebayo, B.O.; Lin, Y.C.; Deng, L.; Rao, Y.K.; Huang, C.C.; Lee, W.H.; Wu, A.T.H.; Hsiao, M.; et al. 4-Acetylantroquinonol B Inhibits Colorectal Cancer Tumorigenesis and Suppresses Cancer Stem-like Phenotype. Toxicol. Appl. Pharm. 2015, 288, 258–268. [Google Scholar] [CrossRef]
- Liu, M.; Bamodu, O.A.; Huang, W.C.; Zucha, M.A.; Lin, Y.K.; Wu, A.T.H.; Huang, C.C.; Lee, W.H.; Yuan, C.C.; Hsiao, M.; et al. 4-Acetylantroquinonol B Suppresses Autophagic Flux and Improves Cisplatin Sensitivity in Highly Aggressive Epithelial Cancer through the PI3K/Akt/MTOR/P70S6K Signaling Pathway. Toxicol. Appl. Pharm. 2017, 325, 48–60. [Google Scholar] [CrossRef]
- Li, T.Y.; Chiang, B.H. 4-Acetylantroquinonol B from Antrodia cinnamomea Enhances Immune Function of Dendritic Cells against Liver Cancer Stem Cells. Biomed. Pharmacother. 2019, 109, 2262–2269. [Google Scholar] [CrossRef]
- Chang, C.H.; Huang, T.F.; Lin, K.T.; Hsu, C.C.; Chang, W.L.; Wang, S.W.; Ko, F.N.; Peng, H.C.; Chung, C.H. 4-Acetylantroquinonol B Suppresses Tumor Growth and Metastasis of Hepatoma Cells via Blockade of Translation-Dependent Signaling Pathway and VEGF Production. J. Agr. Food Chem. 2015, 63, 208–215. [Google Scholar] [CrossRef]
- Chang, T.; Chou, W. Antrodia cinnamomea Sp. Nov. on Cinnamomum kanehirai in Taiwan. Mycol. Res. 1995, 99, 756–758. [Google Scholar] [CrossRef]
- Yang, S.H.; Lin, Y.W.; Chiang, B.H. Biosynthesis of 4-Acetylantroquinonol B in Antrodia cinnamomea via a Pathway Related to Coenzyme Q Synthesis. Biochem. Eng. J. 2017, 117, 23–29. [Google Scholar] [CrossRef]
- Chou, K.C.C.; Yang, S.H.; Wu, H.L.; Lin, P.Y.; Chang, T.L.; Sheu, F.; Chen, K.-H.; Chiang, B.-H. Biosynthesis of Antroquinonol and 4-Acetylantroquinonol B via a Polyketide Pathway Using Orsellinic Acid as a Ring Precursor in Antrodia cinnamomea. J. Agric. Food Chem. 2017, 65, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.C.C.; Wu, H.L.; Lin, P.Y.; Yang, S.H.; Chang, T.L.; Sheu, F.; Chen, K.H.; Chiang, B.H. 4-Hydroxybenzoic acid serves as an endogenous ring precursor for antroquinonol biosynthesis in Antrodia cinnamomea. Phytochemistry 2019, 161, 97–106. [Google Scholar] [CrossRef]
- Yu, P.W.; Chang, Y.C.; Liou, R.F.; Lee, T.H.; Tzean, S.S. Pks63787, a Polyketide Synthase Gene Responsible for the Biosynthesis of Benzenoids in the Medicinal Mushroom Antrodia cinnamomea. J. Nat. Prod. 2016, 79, 1485–1491. [Google Scholar] [CrossRef]
- Yu, P.W.; Cho, T.Y.; Liou, R.F.; Tzean, S.S.; Lee, T.H. Identification of the Orsellinic Acid Synthase PKS63787 for the Biosynthesis of Antroquinonols in Antrodia cinnamomea. Appl. Microbiol. Biot. 2017, 101, 4701–4711. [Google Scholar] [CrossRef]
- Liu, H.; Xing, H.; Jin, Y.; Liu, J.; Tzeng, Y.M.; Deng, L.; Wang, F. Application of Multiple Strategies to Improve the Production of the Potential Cancer Drug 4-Acetylantroquinonol B (4-AAQB) by the Rare Fungus Antrodia cinnamomea. Appl. Biochem. Biotech. 2022, 194, 2720–2730. [Google Scholar] [CrossRef]
- Chiang, C.C.; Huang, T.N.; Lin, Y.W.; Chen, K.H.; Chiang, B.H. Enhancement of 4-Acetylantroquinonol B Production by Supplementation of Its Precursor during Submerged Fermentation of Antrodia Cinnamomea. J. Agr. Food Chem. 2013, 61, 9160–9165. [Google Scholar] [CrossRef]
- Englard, S.; Siegel, L. [18] Mitochondrial l-malate dehydrogenase of beef heart: [EC 1.1.1.37 l-Malate: NAD oxidoreductase]. Method. Enzymol. 1969, 13, 99–106. [Google Scholar]
- Weitzman, J.K.-P. Krebs Citric Acid Cycle: Half a Century and Still Turning. Biochem. Soc. Symposia 1987, 54, 1–198. [Google Scholar]
- Oh, Y.-K.; Palsson, B.O.; Park, S.M.; Schilling, C.H.; Mahadevan, R. Genome-Scale Reconstruction of Metabolic Network in Bacillus Subtilis Based on High-Throughput Phenotyping and Gene Essentiality Data. J. Biol. Chem. 2007, 282, 28791–28799. [Google Scholar] [CrossRef] [Green Version]
- Cooper, T.; Beevers, H. Mitochondria and Glyoxysomes from Castor Bean Endosperm: Enzyme Constituents and Catalytic Capacity. J. Biol. Chem. 1969, 244, 3507–3513. [Google Scholar] [CrossRef]
- Liu, H.; Marsafari, M.; Wang, F.; Deng, L.; Xu, P. Engineering Acetyl-CoA Metabolic Shortcut for Eco-Friendly Production of Polyketides Triacetic Acid Lactone in Yarrowia Lipolytica. Metab. Eng. 2019, 56, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.H.; Wu, H.J.; Ko, Y.H.; Lin, W.H.; Jaiswal, R. Effects of Red Light and Addition of Monoterpenes and Tangerine Oil on the Production of Biomass and Triterpenoids of Antrodia cinnamomea in Submerged Cultures. J. Taiwan Inst. Chem. E 2016, 67, 140–147. [Google Scholar] [CrossRef]
- Bates, A.R. Terpenoids and Squalene: Biosynthesis, Functions and Health Implications; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2015. [Google Scholar]
- Liu, X.; Xia, Y.; Zhang, Y.; Sang, K.; Xiong, Z.; Wang, G.; Liu, X.; Ai, L. RNA-Seq Transcriptomic Analyses of Antrodia camphorata to Determine Antroquinonol and Antrodin C Biosynthetic Mechanisms in the in Situ Extractive Fermentation. J. Sci. Food Agric. 2020, 100, 4252–4262. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, Y.; Zhang, Y.; Yang, C.; Xiong, Z.; Song, X.; Ai, L. Comprehensive Transcriptomic and Proteomic Analyses of Antroquinonol Biosynthetic Genes and Enzymes in Antrodia camphorata. AMB Express 2020, 10, 136. [Google Scholar] [CrossRef] [PubMed]
- Meganathan, R. Biosynthesis of Menaquinone (Vitamin K2) and Ubiquinone (Coenzyme Q): A Perspective on Enzymatic Mechanisms. Vitam. Horm. 2001, 61, 173–218. [Google Scholar] [PubMed]
- Thai, L.; Rush, J.S.; Maul, J.E.; Devarenne, T.; Rodgers, D.L.; Chappell, J.; Waechter, C.J. Farnesol Is Utilized for Isoprenoid Biosynthesis in Plant Cells via Farnesyl Pyrophosphate Formed by Successive Monophosphorylation Reactions. Proc. Natl. Acad. Sci. India B 1999, 96, 13080–13085. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Enzymes | Function |
|---|---|---|
| Glyoxylate and citrate cycle | ACS | Acetyl CoA synthase |
| CS | Citrate synthase | |
| ACUE | Malate synthase | |
| ACU7 | Isocitrate lyase | |
| IDH | Isocitrate dehydrogenase | |
| MVA pathway and terpenoid biosynthesis | SE | Squalene epoxidase |
| SQS | Squalene synthase | |
| HMGS | Hydroxymethylglutaryl-CoA | |
| HMGR | 3-hydroxy-3-methylglutaryl-coenzyme A reductase | |
| AACT | Acetyl CoA acetyltransferase A | |
| MVD | Diphosphomevalonate decarboxylase | |
| MPK | Phosphomevalonate kinase | |
| FPS | Farnesyl pyrophosphate synthase | |
| GPS | Geranyl pyrophosphate synthase | |
| Ubiquinone biosynthesis | PPT1/2 | 4-hydroxybenzoate polyprenyltransferase |
| P4501/2/3 | Monooxygenase | |
| GEDA1/2/3 | Oxygen methyltransferase | |
| COQ6 | Phenol hydroxylase | |
| NAHG | Salicylate hydroxylase | |
| YAT1 | Oxygen acetyltransferase | |
| CAT2 | Oxygen acetyltransferase | |
| METXA | Oxygen acetyltransferase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; Liu, H.; Ning, Y.; Tzeng, Y.-M.; Deng, L.; Wang, F. Understanding the Essential Metabolic Nodes in the Synthesis of 4-Acetylantroquinol B (4-AAQB) by Antrodia cinnamomea Using Transcriptomic Analysis. Fermentation 2023, 9, 542. https://doi.org/10.3390/fermentation9060542
Jin Y, Liu H, Ning Y, Tzeng Y-M, Deng L, Wang F. Understanding the Essential Metabolic Nodes in the Synthesis of 4-Acetylantroquinol B (4-AAQB) by Antrodia cinnamomea Using Transcriptomic Analysis. Fermentation. 2023; 9(6):542. https://doi.org/10.3390/fermentation9060542
Chicago/Turabian StyleJin, Yuhan, Huan Liu, Yuchen Ning, Yew-Min Tzeng, Li Deng, and Fang Wang. 2023. "Understanding the Essential Metabolic Nodes in the Synthesis of 4-Acetylantroquinol B (4-AAQB) by Antrodia cinnamomea Using Transcriptomic Analysis" Fermentation 9, no. 6: 542. https://doi.org/10.3390/fermentation9060542
APA StyleJin, Y., Liu, H., Ning, Y., Tzeng, Y. -M., Deng, L., & Wang, F. (2023). Understanding the Essential Metabolic Nodes in the Synthesis of 4-Acetylantroquinol B (4-AAQB) by Antrodia cinnamomea Using Transcriptomic Analysis. Fermentation, 9(6), 542. https://doi.org/10.3390/fermentation9060542