
Effects of Temperature Shifts on Microbial Communities and Biogas Production: An In-Depth Comparison

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum and Substrates
2.2. Experimental Setup
2.3. Data Collection and Analysis
2.4. NGS and Microbial Diversity Analysis
3. Results
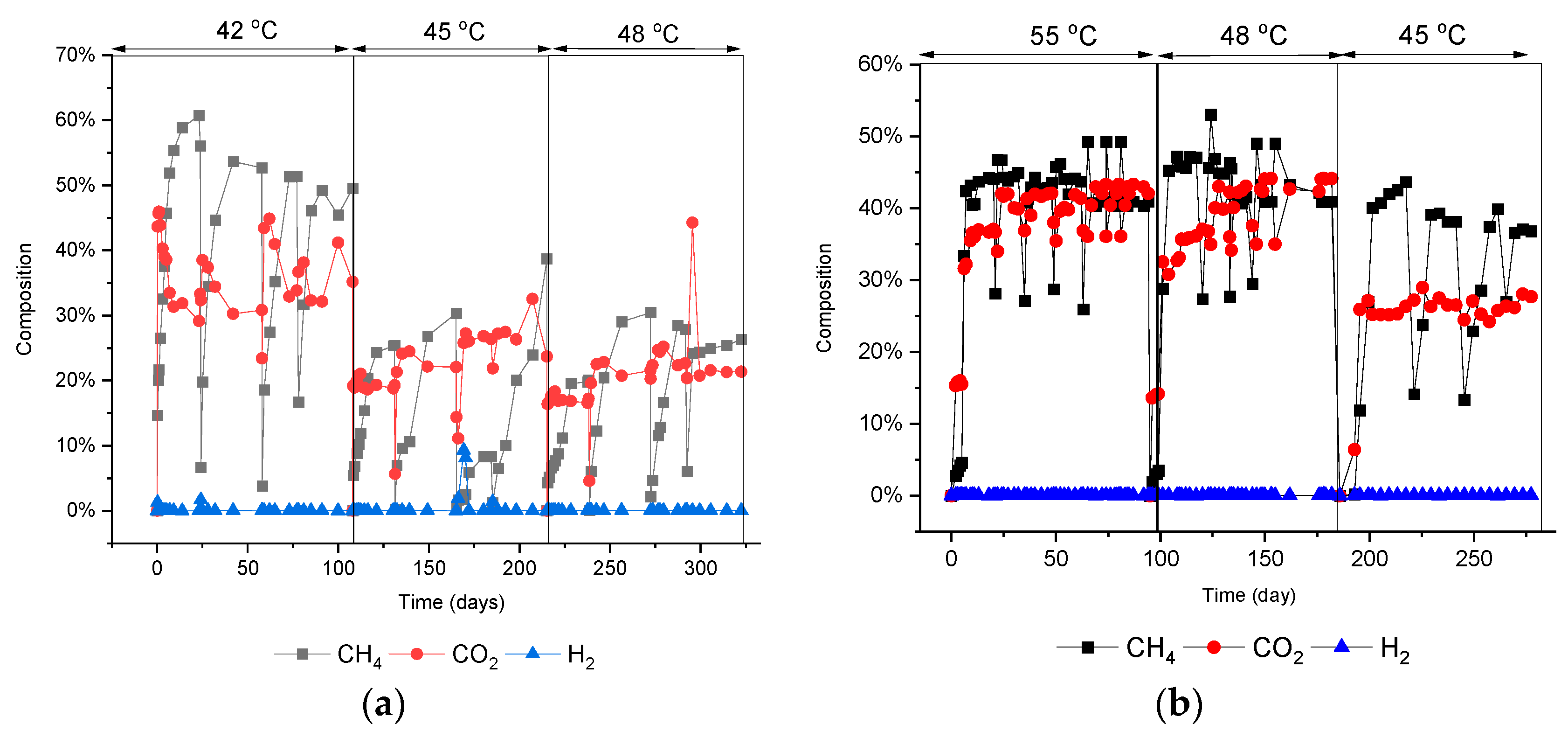
3.1. Biogas Production during Temperature Shifts
3.2. Microbial Response on the Temperature Shift Conditions
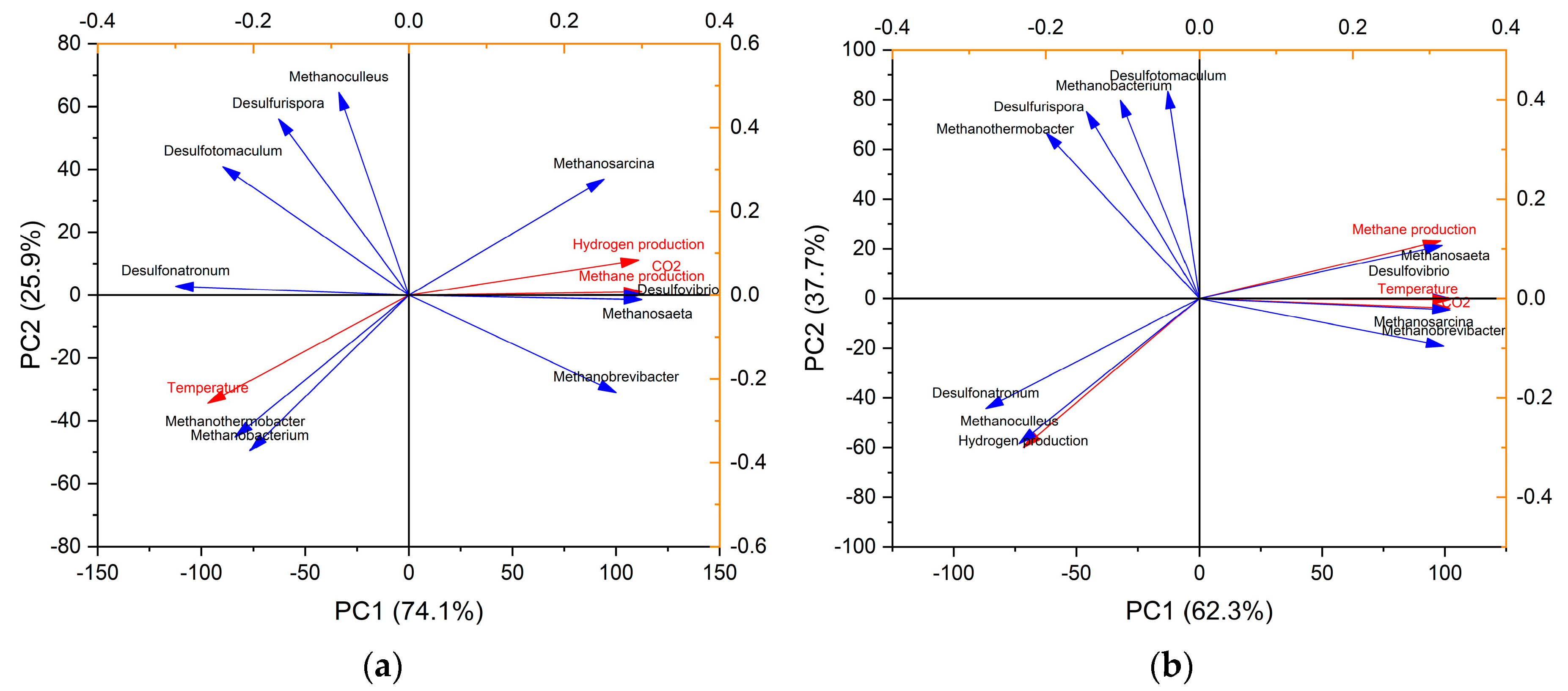
3.3. Correlation Analysis between Methanogen and SRB Populations on Biogas Production
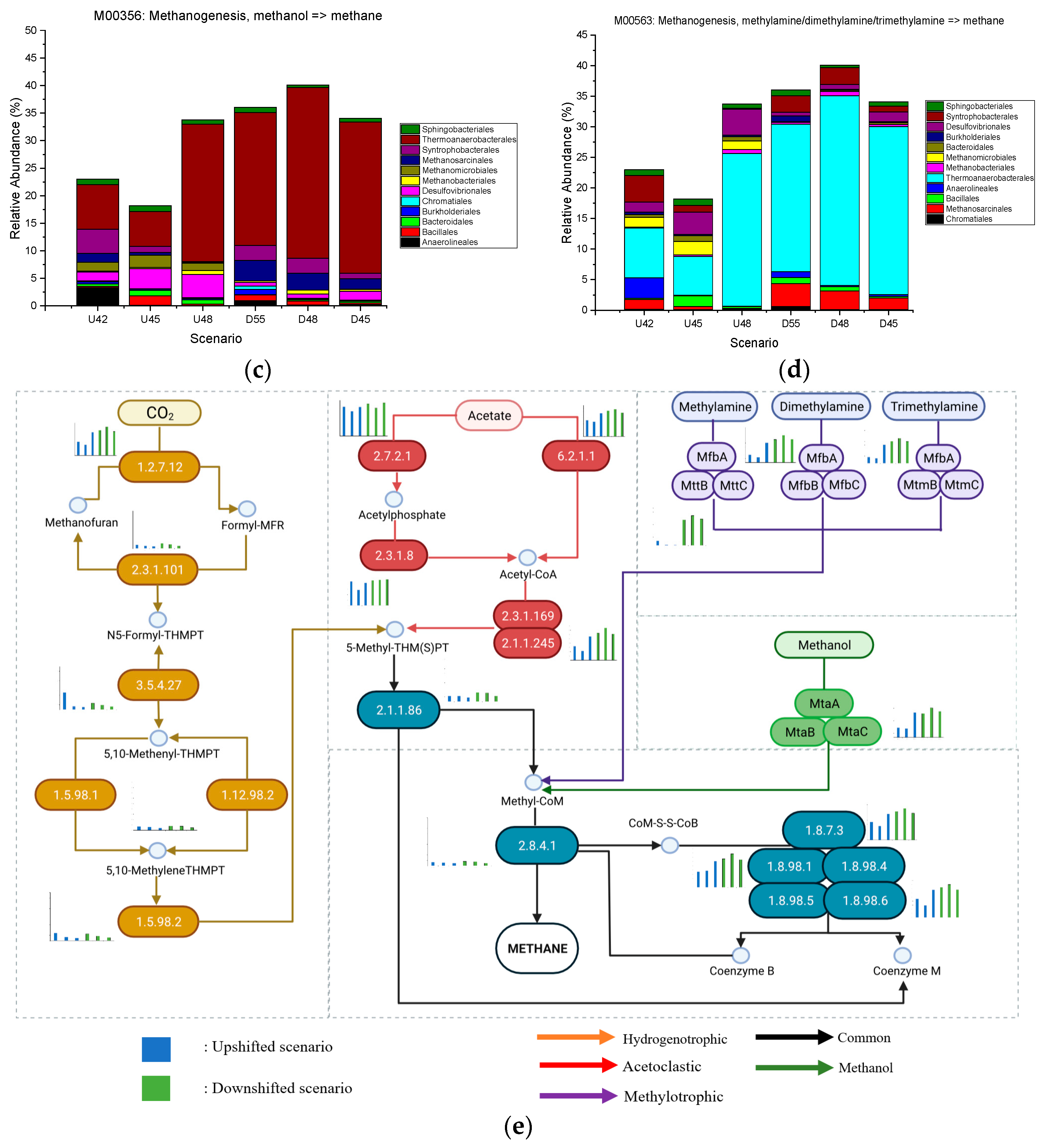
3.4. Effect of Upshifted and Downshifted Temperatures on CH4 Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Porté, H.; Kougias, P.G.; Alfaro, N.; Treu, L.; Campanaro, S.; Angelidaki, I. Process Performance and Microbial Community Structure in Thermophilic Trickling Biofilter Reactors for Biogas Upgrading. Sci. Total Environ. 2019, 655, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, Y. Turning Food Waste to Energy and Resources towards a Great Environmental and Economic Sustainability: An Innovative Integrated Biological Approach. Biotechnol. Adv. 2019, 37, 107414. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.Z.; Fang, S.; Zhang, L.; Huang, W.; Shao, Q.; Fang, F.; Feng, Q.; Cao, J.; Luo, J. Distribution Patterns of Functional Microbial Community in Anaerobic Digesters under Different Operational Circumstances: A Review. Bioresour. Technol. 2021, 341, 125823. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-L.; Zhang, Q.; Xia, Z.-Y.; Gou, M.; Sun, Z.-Y.; Tang, Y.-Q. The Responses of Mesophilic and Thermophilic Anaerobic Digestion of Municipal Sludge to Periodic Fluctuation Disturbance of Organic Loading Rate. Environ. Res. 2022, 218, 114783. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Fotidis, I.A.; Mancini, E.; Treu, L.; Mahdy, A.; Ballesteros, M.; González-Fernández, C.; Angelidaki, I. Acclimation to Extremely High Ammonia Levels in Continuous Biomethanation Process and the Associated Microbial Community Dynamics. Bioresour. Technol. 2018, 247, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Kurade, M.B.; Saha, S.; Salama, E.; Patil, S.M.; Govindwar, S.P. Acetoclastic Methanogenesis Led by Methanosarcina in Anaerobic Co-Digestion of Fats, Oil and Grease for Enhanced Production of Methane. Bioresour. Technol. 2019, 272, 351–359. [Google Scholar] [CrossRef]
- Sudiartha, G.A.W.; Imai, T.; Hung, Y.-T. Effects of Stepwise Temperature Shifts in Anaerobic Digestion for Treating Municipal Wastewater Sludge: A Genomic Study. Int. J. Environ. Res. Public Health 2022, 19, 5728. [Google Scholar] [CrossRef]
- Sudiartha, G.A.W.; Imai, T. An Investigation of Temperature Downshift Influences on Anaerobic Digestion in the Treatment of Municipal Wastewater Sludge. J. Water Environ. Technol. 2022, 20, 154–167. [Google Scholar] [CrossRef]
- Nie, E.; He, P.; Zhang, H.; Hao, L.; Shao, L.; Lü, F. How Does Temperature Regulate Anaerobic Digestion? Renew. Sustain. Energy Rev. 2021, 150, 111453. [Google Scholar] [CrossRef]
- Madigou, C.; Lê Cao, K.A.; Bureau, C.; Mazéas, L.; Déjean, S.; Chapleur, O. Ecological Consequences of Abrupt Temperature Changes in Anaerobic Digesters. Chem. Eng. J. 2019, 361, 266–277. [Google Scholar] [CrossRef]
- Kovalovszki, A.; Treu, L.; Ellegaard, L.; Luo, G.; Angelidaki, I. Modeling Temperature Response in Bioenergy Production: Novel Solution to a Common Challenge of Anaerobic Digestion. Appl. Energy 2020, 263, 114646. [Google Scholar] [CrossRef]
- Hupfauf, S.; Plattner, P.; Wagner, A.O.; Kaufmann, R.; Insam, H.; Podmirseg, S.M. Temperature Shapes the Microbiota in Anaerobic Digestion and Drives Efficiency to a Maximum at 45 °C. Bioresour. Technol. 2018, 269, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Beale, D.J.; Karpe, A.V.; McLeod, J.D.; Gondalia, S.V.; Muster, T.H.; Othman, M.Z.; Palombo, E.A.; Joshi, D. An “omics” Approach towards the Characterisation of Laboratory Scale Anaerobic Digesters Treating Municipal Sewage Sludge. Water Res. 2016, 88, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chang, S. Bioresource Technology Dissecting Methanogenesis for Temperature-Phased Anaerobic Digestion: Impact of Temperature on Community Structure, Correlation, and Fate of Methanogens. Bioresour. Technol. 2020, 306, 123104. [Google Scholar] [CrossRef]
- Fernández-Rodríguez, J.; Pérez, M.; Romero, L.I. Semicontinuous Temperature-Phased Anaerobic Digestion (TPAD) of Organic Fraction of Municipal Solid Waste (OFMSW). Comparison with Single-Stage Processes. Chem. Eng. J. 2016, 285, 409–416. [Google Scholar] [CrossRef]
- Wang, S.; Ruan, Y.; Zhou, W.; Li, Z.; Wu, J.; Liu, D. Net Energy Analysis of Small-Scale Biogas Self-Supply Anaerobic Digestion System Operated at Psychrophilic to Thermophilic Conditions. J. Clean. Prod. 2018, 174, 226–236. [Google Scholar] [CrossRef]
- Kasinski, S. Mesophilic and Thermophilic Anaerobic Digestion of Organic Fraction Separated during Mechanical Heat Treatment of Municipal Waste. Appl. Sci. 2020, 10, 2412. [Google Scholar] [CrossRef] [Green Version]
- Hidaka, T.; Nakamura, M.; Oritate, F.; Nishimura, F. Comparative Anaerobic Digestion of Sewage Sludge at Different Temperatures with and without Heat Pre-Treatment. Chemosphere 2022, 307, 135808. [Google Scholar] [CrossRef]
- Roopnarain, A.; Rama, H.; Ndaba, B.; Bello-Akinosho, M.; Bamuza-Pemu, E.; Adeleke, R. Unravelling the Anaerobic Digestion ‘Black Box’: Biotechnological Approaches for Process Optimization. Renew. Sustain. Energy Rev. 2021, 152, 111717. [Google Scholar] [CrossRef]
- Westerholm, M.; Isaksson, S.; Karlsson Lindsjö, O.; Schnürer, A. Microbial Community Adaptability to Altered Temperature Conditions Determines the Potential for Process Optimisation in Biogas Production. Appl. Energy 2018, 226, 838–848. [Google Scholar] [CrossRef]
- Zhou, W.; Imai, T.; Ukita, M.; Sekine, M.; Higuchi, T. Triggering Forces for Anaerobic Granulation in UASB Reactors. Process Biochem. 2006, 41, 36–43. [Google Scholar] [CrossRef]
- Sakaveli, F.; Petala, M.; Tsiridis, V.; Darakas, E. Enhanced Mesophilic Anaerobic Digestion of Primary Sewage Sludge. Water 2021, 13, 348. [Google Scholar] [CrossRef]
- García-Depraect, O.; Martínez-Mendoza, L.J.; Diaz, I.; Muñoz, R. Two-Stage Anaerobic Digestion of Food Waste: Enhanced Bioenergy Production Rate by Steering Lactate-Type Fermentation during Hydrolysis-Acidogenesis. Bioresour. Technol. 2022, 358, 127358. [Google Scholar] [CrossRef] [PubMed]
- Sittijunda, S.; Reungsang, A.; O-thong, S. Biohydrogen Production from Dual Digestion Pretreatment of Poultry Slaughterhouse Sludge by Anaerobic Self-Fermentation. Int. J. Hydrogen Energy 2010, 35, 13427–13434. [Google Scholar] [CrossRef]
- Kobayashi, T.; Yasuda, D.; Li, Y.-Y.; Kubota, K.; Harada, H.; Okaniwa, Y. Start-up Method of Anaerobic Digestion by Anaerobic Self-Degradation of Waste Activated Sludge and Microbial Community Shift during Self-Degradation. J. Jpn. Soc. Water Environ. 2008, 31, 525–532. [Google Scholar] [CrossRef]
- Signorini, A.; Lembo, G.; Giulia, M.; Pizzichini, D.; Fenice, M. Self-Fermentation: Innovative Pretreatment for Use High Concentration of Olive Mill Wastewater in Anaerobic Digestion Plant. In Proceedings of the 24th European Biomass Conference and Exhibition, Amsterdam, The Netherlands, 6–9 June 2016. [Google Scholar]
- Tetteh, E.K.; Rathilal, S. Biophotocatalytic Reduction of CO2 in Anaerobic Biogas Produced from Wastewater Treatment Using an Integrated System. Catalysts 2022, 12, 76. [Google Scholar] [CrossRef]
- Ángeles, R.; Vega-Quiel, M.J.; Batista, A.; Fernández-Ramos, O.; Lebrero, R.; Muñoz, R. Influence of Biogas Supply Regime on Photosynthetic Biogas Upgrading Performance in an Enclosed Algal-Bacterial Photobioreactor. Algal Res. 2021, 21, 102350. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y.; Dong, R. Recent Progress towards In-Situ Biogas Upgrading Technologies. Sci. Total Environ. 2021, 800, 149667. [Google Scholar] [CrossRef]
- Moestedt, J.; Nordell, E.; Schnürer, A. Comparison of Operating Strategies for Increased Biogas Production from Thin Stillage. J. Biotechnol. 2014, 175, 22–30. [Google Scholar] [CrossRef]
- Sun, L.; Pope, P.B.; Eijsink, V.G.H.; Schnürer, A. Characterization of Microbial Community Structure during Continuous Anaerobic Digestion of Straw and Cow Manure. Microb. Biotechnol. 2015, 8, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Westerholm, M.; Müller, B.; Isaksson, S.; Schnürer, A. Trace Element and Temperature Effects on Microbial Communities and Links to Biogas Digester Performance at High Ammonia Levels. Biotechnol. Biofuels 2015, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, G.T.W.; Weng, C.Y.; Chen, C.Y.; Weng, F.C.H.; Wang, D. A Systematic Approach Re-Analyzing the Effects of Temperature Disturbance on the Microbial Community of Mesophilic Anaerobic Digestion. Sci. Rep. 2019, 9, 6560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arelli, V.; Mamindlapelli, N.K.; Begum, S.; Juntupally, S.; Anupoju, G.R. Solid State Anaerobic Digestion of Food Waste and Sewage Sludge: Impact of Mixing Ratios and Temperature on Microbial Diversity, Reactor Stability and Methane Yield. Sci. Total Environ. 2021, 793, 148586. [Google Scholar] [CrossRef] [PubMed]
- Hochheimer, A.; Schmitz, R.A.; Thauer, R.K.; Hedderich, R. The Tungsten Formylmethanofuran Dehydrogenase from Methanobacterium Thermoautotrophicum Contains Sequence Motifs Characteristic for Enzymes Containing Molybdopterin Dinucleotide. Eur. J. Biochem. 1995, 234, 910–920. [Google Scholar] [CrossRef]
- Vorholt, J.A.; Thauer, R.K. The Active Species of “CO2” Utilized by Formylmethanofuran Dehydrogenase from Methanogenic Archaea. Eur. J. Biochem. 1997, 248, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Barnhart, E.P.; Mcclure, M.A.; Johnson, K.; Cleveland, S.; Hunt, K.A.; Fields, M.W. Potential Role of Acetyl-CoA Synthetase (Acs) and Malate Dehydrogenase (Mae) in the Evolution of the Acetate Switch in Bacteria and Archaea. Sci. Rep. 2015, 5, 12498. [Google Scholar] [CrossRef] [Green Version]
- Grahame, D.A. Methods for Analysis of Acetyl-CoA Synthase: Applications to Bacterial and Archaeal Systems, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 494, ISBN 9780123851123. [Google Scholar]
- Zhang, S.; Xiao, M.; Liang, C.; Chui, C.; Wang, N.; Shi, J.; Liu, L. Multivariate Insights into Enhanced Biogas Production in Thermophilic Dry Anaerobic Co-Digestion of Food Waste with Kitchen Waste or Garden Waste: Process Properties, Microbial Communities and Metagenomic Analyses. Bioresour. Technol. 2022, 361, 127684. [Google Scholar] [CrossRef]
- Ziembińska-Buczyńska, A.; Banach, A.; Bacza, T.; Pieczykolan, M. Diversity and Variability of Methanogens during the Shift from Mesophilic to Thermohilic Conditions While Biogas Production. World J. Microbiol. Biotechnol. 2014, 30, 3047–3053. [Google Scholar] [CrossRef]
- Wang, S.; Ma, F.; Ma, W.; Wang, P.; Zhao, G.; Lu, X. Influence of Temperature on Biogas Production Efficiency and Microbial Community in a Two-Phase Anaerobic Digestion System. Water 2019, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Babaei, A.; Shayegan, J. Effects of Temperature and Mixing Modes on the Performance of Municipal Solid Waste Anaerobic Slurry Digester 09 Engineering 0907 Environmental Engineering 09 Engineering 0904 Chemical Engineering. J. Environ. Health Sci. Eng. 2019, 17, 1077–1084. [Google Scholar] [CrossRef]
- Conrad, R. Contribution of Hydrogen to Methane Production and Control of Hydrogen Concentrations in Methanogenic Soils and Sediments. FEMS Microbiol. Ecol. 1999, 28, 193–202. [Google Scholar] [CrossRef]
- Metje, M.; Frenzel, P. Methanogenesis and Methanogenic Pathways in a Peat from Subarctic Permafrost. Environ. Microbiol. 2007, 9, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Liebner, S.; Ganzert, L.; Kiss, A.; Yang, S.; Wagner, D.; Svenning, M.M. Shifts in Methanogenic Community Composition and Methane Fluxes along the Degradation of Discontinuous Permafrost. Front. Microbiol. 2015, 6, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, R.; Xu, C.; Shi, X.; Yang, S.; Li, L.; Peng, X.; Song, L. Acetoclastic Methanogenesis Pathway Stability despite the High Microbial Taxonomic Variability in the Transition from Acidogenesis to Methanogenesis during Food Waste Anaerobic Digestion. J. Clean. Prod. 2022, 372, 133758. [Google Scholar] [CrossRef]
- Ho, D.P.; Jensen, P.D.; Batstone, D.J. Methanosarcinaceae and Acetate-Oxidizing Pathways Dominate in High-Rate Thermophilic Anaerobic Digestion of Waste-Activated Sludge. Appl. Environ. Microbiol. 2013, 79, 6491–6500. [Google Scholar] [CrossRef] [Green Version]
- Krohn, C.; Dias, D.A.; Den Akker, V.; Rees, C.A.; Crosbie, N.D.; Carroll, D.M.O.; Stuetz, R.M.; Batstone, D.J.; Ball, A.S. The Role of Microbial Ecology in Improving the Performance of Anaerobic Digestion of Sewage Sludge. Front. Microbiol. 2022, 13, 1079136. [Google Scholar] [CrossRef]
- De Vrieze, J.; Hennebel, T.; Boon, N.; Verstraete, W. Bioresource Technology Methanosarcina: The Rediscovered Methanogen for Heavy Duty Biomethanation. Bioresour. Technol. 2012, 112, 1–9. [Google Scholar] [CrossRef]
- Tian, H.; Fotidis, I.A.; Kissas, K.; Angelidaki, I. Bioresource Technology E Ff Ect of Di Ff Erent Ammonia Sources on Aceticlastic and Hydrogenotrophic Methanogens. Bioresour. Technol. 2018, 250, 390–397. [Google Scholar] [CrossRef] [Green Version]
- Demirel, B.; Yenigu, O. Two-Phase Anaerobic Digestion Processes: A Review. J. Chem. Technol. Biotechnol. 2002, 77, 743–755. [Google Scholar] [CrossRef]
- Harirchi, S.; Wainaina, S.; Sar, T.; Nojoumi, S.A.; Parchami, M.; Varjani, S.; Khanal, S.K.; Awasthi, M.K.; Taherzadeh, M.J.; Harirchi, S.; et al. Microbiological Insights into Anaerobic Digestion for Biogas, Hydrogen or Volatile Fatty Acids (VFAs): A Review. Bioengineered 2022, 13, 6521–6557. [Google Scholar] [CrossRef]
- St-pierre, B. Implications from Distinct Sulfate-Reducing Bacteria Populations between Cattle Manure and Digestate in the Elucidation of H2S Production during Anaerobic Digestion of Animal Slurry. Appl. Microbiol. Biotechnol. 2017, 101, 5543–5556. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.T.; Kannoorpatti, K.; Padovan, A.; Thennadil, S. Sulphate-Reducing Bacteria’s Response to Extreme pH Environments and the Effect of Their Activities on Microbial Corrosion. Appl. Sci. 2021, 11, 2201. [Google Scholar] [CrossRef]
- Wang, F.; Peng, S.; Fan, L.; Li, Y. Improved Sulfate Reduction Efficiency of Sulfate- Reducing Bacteria in Sulfate-Rich Systems by Acclimatization and Multiple-Grouting. Alexandria Eng. J. 2022, 61, 9993–10005. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Anaerobic Sludge | Units |
|---|---|---|
| pH | 8.17 | - |
| Total Solid (TS) | 8 | % w/v |
| Volatile Solid (VS) | 3 | % w/v |
| Fixed Solid (FS) | 5 | % w/v |
| VS/TS ratio | 0.6 | - |
| Conditions | Labels | Scenario |
|---|---|---|
| 42 °C | U42 | Upshifted |
| 42 °C → 45 °C | U45 | Upshifted |
| 45 °C → 48 °C | U48 | Upshifted |
| 55 °C | D55 | Downshifted |
| 55 °C→48 °C | D48 | Downshifted |
| 48 °C → 45 °C | D45 | Downshifted |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudiartha, G.A.W.; Imai, T.; Mamimin, C.; Reungsang, A. Effects of Temperature Shifts on Microbial Communities and Biogas Production: An In-Depth Comparison. Fermentation 2023, 9, 642. https://doi.org/10.3390/fermentation9070642
Sudiartha GAW, Imai T, Mamimin C, Reungsang A. Effects of Temperature Shifts on Microbial Communities and Biogas Production: An In-Depth Comparison. Fermentation. 2023; 9(7):642. https://doi.org/10.3390/fermentation9070642
Chicago/Turabian StyleSudiartha, Gede Adi Wiguna, Tsuyoshi Imai, Chonticha Mamimin, and Alissara Reungsang. 2023. "Effects of Temperature Shifts on Microbial Communities and Biogas Production: An In-Depth Comparison" Fermentation 9, no. 7: 642. https://doi.org/10.3390/fermentation9070642
APA StyleSudiartha, G. A. W., Imai, T., Mamimin, C., & Reungsang, A. (2023). Effects of Temperature Shifts on Microbial Communities and Biogas Production: An In-Depth Comparison. Fermentation, 9(7), 642. https://doi.org/10.3390/fermentation9070642