
Examining the Impact of Substrate Composition on the Biochemical Properties and Antioxidant Activity of Pleurotus and Agaricus Mushrooms

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mushroom Production
2.2. Reagents and Chemicals
2.3. Total Intra-Cellular Polysaccharide (IPS) Determination
2.4. Quantitative Evaluation of a- and β-Glucan Content
2.5. Total Protein Determination
2.6. Determination of Total Lipids and Fatty Acids
2.7. Minerals
2.8. Determination of Tocopherols
2.9. Total Phenolic Compounds and Antioxidant Activity
2.9.1. Determination of Total Phenolic Compounds (TPC)
2.9.2. Antioxidant Activity: Ferric Reducing Antioxidant Power (FRAP)
2.9.3. Antioxidant Activity: Scavenging Activity of ABTS + Radical
2.9.4. Antioxidant Activity: Scavenging Ability on 1.1-Diphenyl-2-Picrylhydrazyl (DPPH) Radicals
2.9.5. Statistical Analysis
2.9.6. Abbreviations
3. Results and Discussion
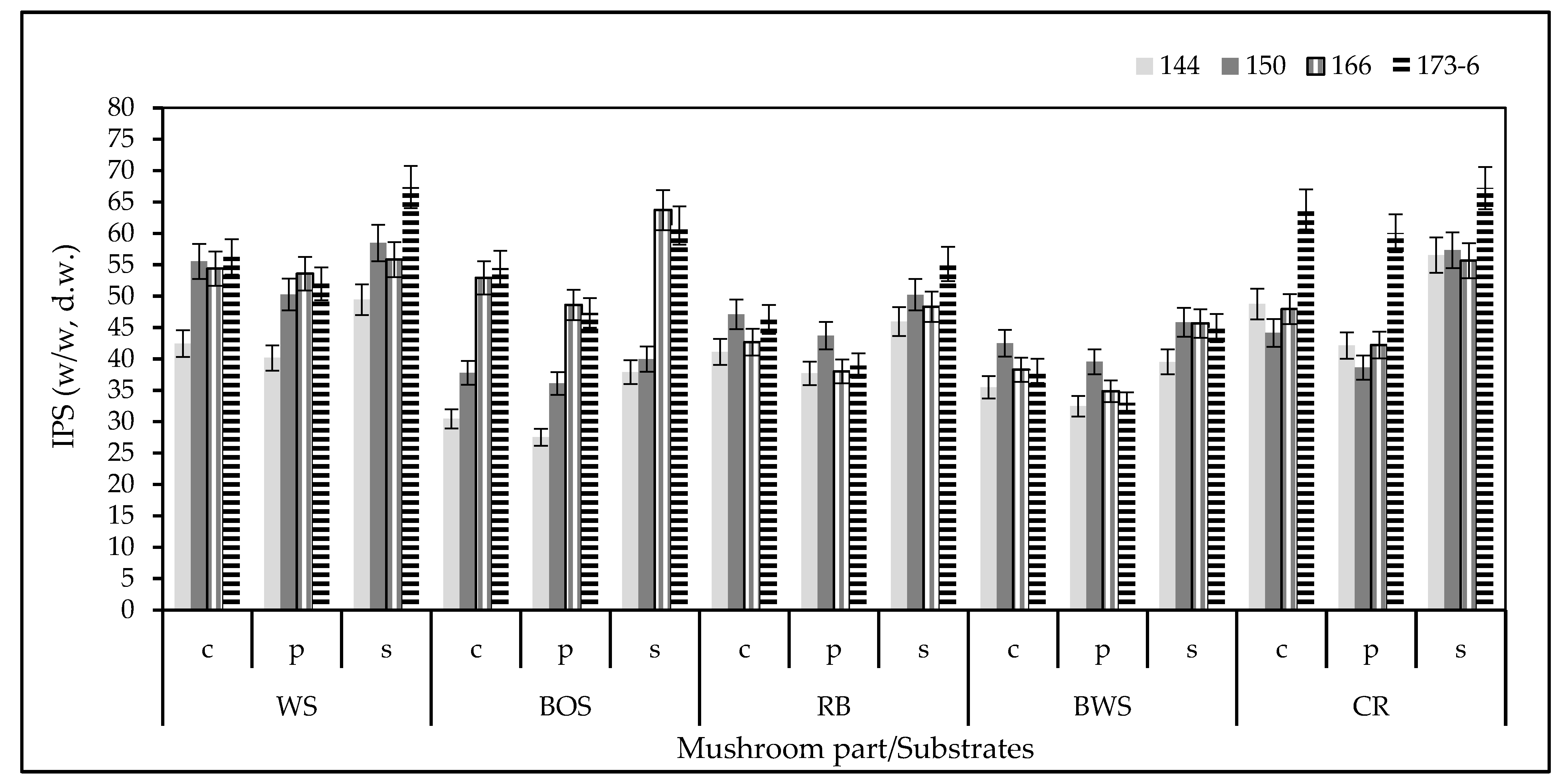
3.1. Intra-Cellular Polysaccharide (IPS) Content and Profile
3.2. Total Glucans, α-Glucans and β-Glucans
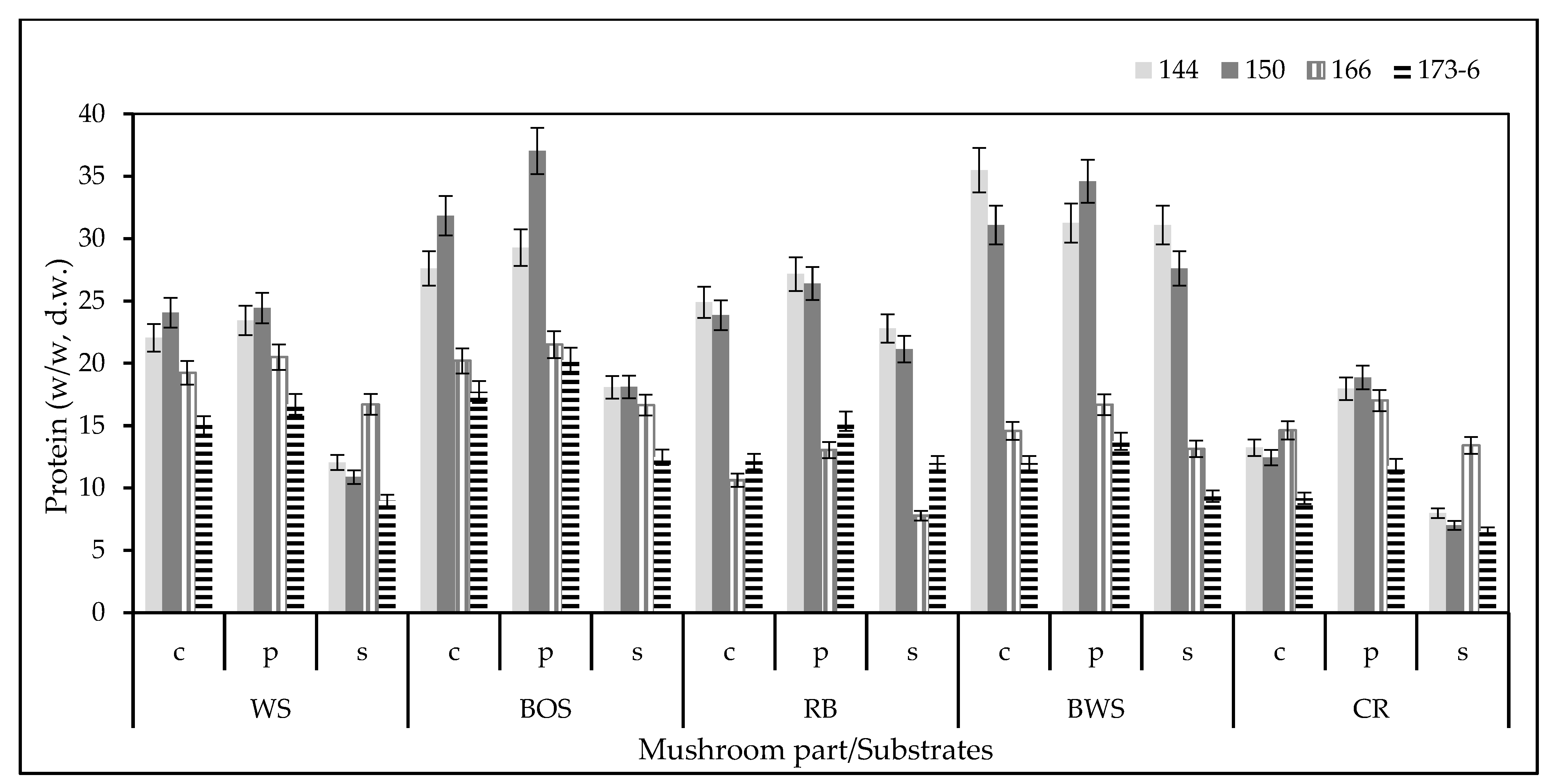
3.3. Total Protein
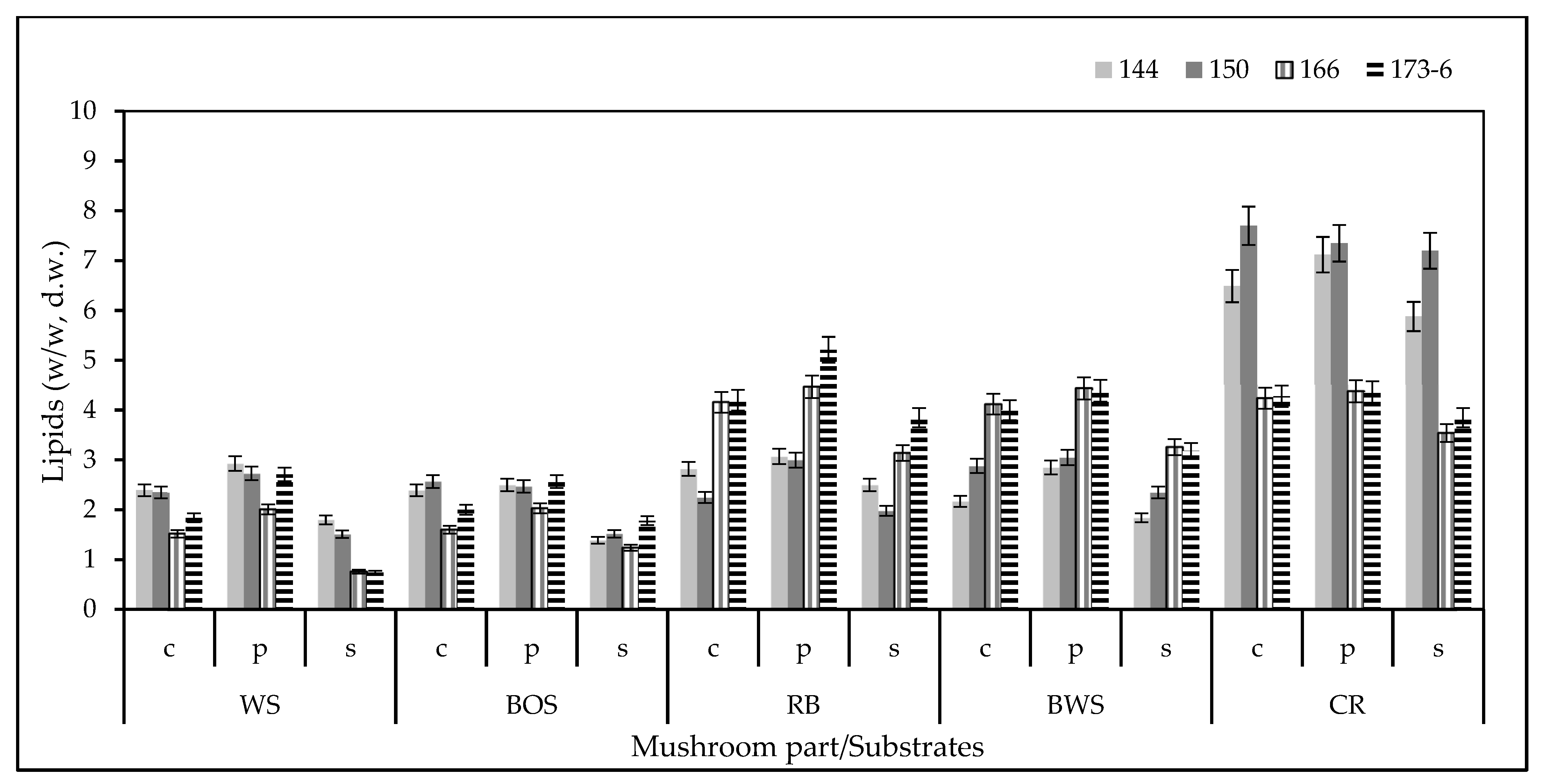
3.4. Total Lipid Determination and Fatty Acid (FA) Composition
3.5. Tocopherols
3.6. Total Phenolic Compounds (TPC), Antioxidant Studies and Total Reducing Power
3.7. Moisture, Ash Content and Mineral Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valverde, M.E.; Hernández-Pérez, T.; Paredes-López, O. Edible mushrooms: Improving human health and promoting quality life. Int. J. Microbiol. 2015, 2015, 376387. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-M.; Zhang, J.; Wu, L.-H.; Zhao, Y.-L.; Li, T.; Li, J.-Q.; Wang, Y.; Liu, H.-G. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014, 151, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Miles, P.G. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2004; p. 451. [Google Scholar]
- Hetland, G.; Johnson, E.; Bernardshaw, S.V.; Grinde, B. Can medicinal mushrooms have prophylactic or therapeutic effect against COVID-19 and its pneumonic superinfection and complicating inflammation? Scand. J. Immunol. 2021, 93, e12937. [Google Scholar] [CrossRef]
- Kalač, P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. Food Chem. 2009, 113, 9–16. [Google Scholar] [CrossRef]
- Manikandan, K. Nutritional and medicinal values of mushrooms. In Mushrooms Cultivation, Marketing and Consumption, 1st ed.; Singh, M., Vijay, B., Kamal, S., Wakchaure, G.C., Eds.; Directorate of Mushroom Research: Solan, India, 2011. [Google Scholar]
- Kalač, P. A review of chemical composition and nutritional value of wild-growing and cultivated mushrooms: Chemical composition of edible mushrooms. J. Sci. Food Agric. 2013, 93, 209–218. [Google Scholar] [CrossRef]
- Pandey, N.; Budhathoki, U. Protein determination through Bradford’s method of Nepalese mushroom. Sci. World 2010, 5, 85–88. [Google Scholar] [CrossRef]
- Oei, P. Manual on Mushroom Cultivation: Techniques Species and Opportunities for Commercial Application in Developing Countries; TOOL: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Singdevsachan, S.K.; Auroshree, P.; Mishra, J.; Baliyarsingh, B.; Tayung, K.; Thatoi, H. Mushroom polysaccharides as potential prebiotics with their antitumor and immunomodulating properties: A review. Bioact. Carbohydr. Diet. Fibre 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Afiati, F.; Firza, S.F.; Kusmiati; Aliya, L.S. The effectiveness β-glucan of shiitake mushrooms and Saccharomyces cerevisiae as antidiabetic and antioxidant in mice Sprague Dawley induced alloxan. AIP Conf. Proc. 2019, 2120, 070006. [Google Scholar] [CrossRef]
- Khan, A.; Gani, A.; Khanday, F.; Masoodi, F.A. Biological and pharmaceutical activities of mushroom β-glucan discussed as a potential functional food ingredient. Bioact. Carbohydr. Diet. Fibre 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Murphy, E.J.; Rezoagli, E.; Major, I.; Rowan, N.J.; Laffey, J.G. β-glucan metabolic and immunomodulatory properties and potential for clinical application. J. Fungi 2020, 6, 356. [Google Scholar] [CrossRef]
- Raman, A.P.; Anoma, M.A.; Zhu, L.; Rephaeli, E.; Fan, S. Passive radiative cooling below ambient air temperature under direct sunlight. Nature 2014, 515, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Ahn, W.-S.; Kim, D.-J.; Chae, G.-T.; Lee, J.-M.; Bae, S.-M.; Sin, J.-I.; Kim, Y.-W.; Namkoong, S.-E.; Lee, I.P. Natural killer cell activity and quality of life were improved by consumption of a mushroom extract, Agaricus blazei Murill Kyowa, in gynecological cancer patients undergoing chemotherapy. Int. J. Gynecol. Cancer 2004, 14, 589–594. [Google Scholar] [CrossRef]
- Ishihara, T. Polysaccharide of Agaricus blazei Murrill. In Handbook of Active Substances in Plant Resources, 2nd ed.; Tanimura, A., Ed.; Science Forum: Tokyo, Japan, 1999; pp. 348–350. [Google Scholar]
- He, J.-Z.; Ru, Q.-M.; Dong, D.-D.; Sun, P.-L. Chemical characteristics and antioxidant properties of crude water soluble polysaccharides from four common edible mushrooms. Molecules 2012, 17, 4373–4387. [Google Scholar] [CrossRef] [PubMed]
- Sande, D.; de Oliveira, G.P.; e Moura, M.A.F.; de Almeida Martins, B.; Lima, M.T.N.S.; Takahashi, J.A. Edible mushrooms as a ubiquitous source of essential fatty acids. Food Res. Int. 2019, 125, 108524. [Google Scholar] [CrossRef] [PubMed]
- Günç Ergönül, P.; Akata, I.; Kalyoncu, F.; Ergönül, B. Fatty acid compositions of six wild edible mushroom species. Sci. World J. 2013, 2013, 163964. [Google Scholar] [CrossRef] [PubMed]
- Holighaus, G.; Rohlfs, M. Volatile and non-volatile fungal oxylipins in fungus-invertebrate interactions. Fungal Ecol. 2019, 38, 28–36. [Google Scholar] [CrossRef]
- Kües, U.; Khonsuntia, W.; Subba, S.; Dörnte, B. Volatiles in communication of Agaricomycetes. In Physiology and Genetics; Anke, T., Schüffler, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 149–212. [Google Scholar]
- Von Arx, M.; Schmidt-Büsser, D.; Guerin, P.M. Plant volatiles enhance behavioral responses of grapevine moth males, Lobesia botrana to sex pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Gursoy, N.; Sarikurkcu, C.; Tepe, B.; Halil Solak, M. Evaluation of antioxidant activities of 3 edible mushrooms: Ramaria flava (Schaef.: Fr.) Quél., Rhizopogon roseolus (Corda) T.M. Fries., and Russula delica Fr. Food Sci. Biotechnol. 2010, 19, 691–696. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Anti-obesity effects of medicinal and edible mushrooms. Molecules 2018, 23, 2880. [Google Scholar] [CrossRef]
- Côté, J.; Caillet, S.; Doyon, G.; Sylvain, J.-F.; Lacroix, M. Bioactive compounds in cranberries and their biological properties. Crit. Rev. Food Sci. Nutr. 2010, 50, 666–679. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Calhelha, R.C.; Vaz, J.A.; Ferreira, I.C.F.R.; Baptista, P.; Estevinho, L.M. Antimicrobial activity and bioactive compounds of Portuguese wild edible mushrooms methanolic extracts. Eur. Food Res. Technol. 2007, 225, 151–156. [Google Scholar] [CrossRef]
- Glamočlija, J.; Ćirić, A.; Nikolić, M.; Fernandes, Â.; Barros, L.; Calhelha, R.C.; Ferreira, I.C.F.R.; Soković, M.; Van Griensven, L.J.L.D. Chemical characterization and biological activity of Chaga (Inonotus obliquus), a medicinal “mushroom”. J. Ethnopharmacol. 2015, 162, 323–332. [Google Scholar] [CrossRef]
- Kumar, K. Role of edible mushroom as functional foods: A review. South Asian J. Food Technol. Environ. 2015, 1, 211–218. [Google Scholar] [CrossRef]
- Muszyńska, B.; Kała, K.; Rojowski, J.; Grzywacz, A.; Opoka, W. Composition and biological properties of Agaricus bisporus fruiting bodies—A Review. Pol. J. Food Nutr. Sci. 2017, 67, 173–181. [Google Scholar] [CrossRef]
- Sánchez, C. Cultivation of Pleurotus ostreatus and other edible mushrooms. Appl. Microbiol. Biotechnol. 2010, 85, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Torres-Martínez, B.D.M.; Vargas-Sánchez, R.D.; Torrescano-Urrutia, G.R.; Esqueda, M.; Rodríguez-Carpena, J.G.; Fernández-López, J.; Perez-Alvarez, J.A.; Sánchez-Escalante, A. Pleurotus genus as a potential ingredient for meat products. Foods 2022, 11, 779. [Google Scholar] [CrossRef] [PubMed]
- Olivier, J.M.; Houdeau, G.; Delpech, P.; Laborde, J. Developments in the cultivation of specialty mushrooms with emphasis on Pleurotus and Shiitake. Mushroom Inf. 1994, 96, 5–19. [Google Scholar]
- Wisitrassameewong, K.; Karunarathna, S.C.; Thongklang, N.; Zhao, R.; Callac, P.; Moukha, S.; Férandon, C.; Chukeatirote, E.; Hyde, K.D. Agaricus subrufescens: A review. Saudi J. Biol. Sci. 2012, 19, 131–146. [Google Scholar] [CrossRef]
- Gülser, C.; Pekşen, A. Using tea waste as a new casing material in mushroom (Agaricus bisporus (L.) Sing.) cultivation. Bioresour. Technol. 2003, 88, 153–156. [Google Scholar] [CrossRef]
- Pardo-Giménez, A.; Pardo, J.E.; Dias, E.S.; Rinker, D.L.; Caitano, C.E.C.; Zied, D.C. Optimization of cultivation techniques improves the agronomic behavior of Agaricus subrufescens. Sci. Rep. 2020, 10, 8154. [Google Scholar] [CrossRef]
- Sassine, Y.N. (Ed.) Mushrooms: Agaricus bisporus Crop Production Science in Horticulture; CAB International: Boston, MA, USA, 2021. [Google Scholar]
- Wang, Q.; Li, B.B.; Li, H.; Han, J.R. Yield, dry matter and polysaccharides content of the mushroom Agaricus blazei produced on asparagus straw substrate. Sci. Hortic. 2010, 125, 16–18. [Google Scholar] [CrossRef]
- Zaghi Junior, L.L.; Linde, G.A.; Colauto, N.B. Carbon-to-nitrogen ratios for Agaricus brasiliensis on the axenic method. Acta Sci. Agron. 2010, 32, 55–60. [Google Scholar]
- Colak, M.; Ergun, B.; Simsek, H.; Hilmi, T.; Yilmaz, F. Cultivation of Agaricus bisporus on wheat straw and waste tea leaves based composts and locally available casing materials Part III: Dry matter, protein, and carbohydrate contents of Agaricus bisporus. Afr. J. Biotechnol. 2007, 6, 2855–2859. [Google Scholar] [CrossRef]
- Lavelli, V.; Proserpio, C.; Gallotti, F.; Laureati, M.; Pagliarini, E. Circular reuse of bio-resources: The role of Pleurotus spp. in the development of functional foods. Food Funct. 2018, 9, 1353–1372. [Google Scholar] [CrossRef] [PubMed]
- Dedousi, M.; Melanouri, E.-M.; Diamantopoulou, P. Carposome productivity of Pleurotus ostreatus and Pleurotus eryngii growing on agro-industrial residues enriched with nitrogen, calcium salts and oils. Carbon Resour. Convers. 2023, 6, 150–165. [Google Scholar] [CrossRef]
- Economou, C.N.; Philippoussis, A.N.; Diamantopoulou, P.A. Spent mushroom substrate for a second cultivation cycle of Pleurotus mushrooms and dephenolization of agro-industrial wastewaters. FEMS Microbiol. Lett. 2020, 367, fnaa060. [Google Scholar] [CrossRef]
- Melanouri, E.-M.; Dedousi, M.; Diamantopoulou, P. Cultivating Pleurotus ostreatus and Pleurotus eryngii mushroom strains on agro-industrial residues in solid-state fermentation. Part I: Screening for growth, endoglucanase, laccase and biomass production in the colonization phase. Carbon Resour. Convers. 2022, 5, 61–70. [Google Scholar] [CrossRef]
- Papadaki, A.; Kachrimanidou, V.; Papanikolaou, S.; Philippoussis, A.; Diamantopoulou, P. Upgrading grape pomace through Pleurotus spp. cultivation for the production of enzymes and fruiting bodies. Microorganisms 2019, 7, 207. [Google Scholar] [CrossRef]
- Philippoussis, A.; Zervakis, G.; Diamantopoulou, P. Bioconversion of agricultural lignocellulosic wastes through the cultivation of the edible mushrooms Agrocybe aegerita, Volvariella volvacea and Pleurotus spp. World J. Microbiol. Biotechnol. 2001, 17, 191–200. [Google Scholar] [CrossRef]
- Reddy, G.V.; Ravindra Babu, P.; Komaraiah, P.; Roy, K.R.R.M.; Kothari, I.L. Utilization of banana waste for the production of lignolytic and cellulolytic enzymes by solid substrate fermentation using two Pleurotus species (P. ostreatus and P. sajor-caju). Process Biochem. 2003, 38, 1457–1462. [Google Scholar] [CrossRef]
- Garg, V.K.; Gupta, R. Vermicomposting of Agro-Industrial Processing Waste. In Biotechnology for Agro-Industrial Residues Utilisation: Utilisation of Agro-Residues; Springer: Dordrecht, The Netherlands, 2009; pp. 431–456. [Google Scholar]
- Philippoussis, A.N. Production of mushrooms using agro-industrial residues as substrates. In Biotechnology for Agro-Industrial Residues Utilisation; Singh Nee’ Nigam, P., Pandey, A., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 163–196. [Google Scholar]
- Zhang, R.; Li, X.; Fadel, J.G. Oyster mushroom cultivation with rice and wheat straw. Bioresour. Technol. 2002, 82, 277–284. [Google Scholar] [CrossRef]
- Croan, S. Conversion of wood waste into value-added products by edible and medicinal Pleurotus (Fr.) P. Karst. species (Agaricales sl, Basidiomycetes). Int. J. Med. Mushrooms 2000, 2, 73–80. [Google Scholar] [CrossRef]
- Sivaprakasam, K.; Kandaswamy, T. Waste materials for the cultivation of Pleurotus sajor-caju. Mushroom J. 1981, 101, 178–179. [Google Scholar]
- Yildiz, S.; Yildiz, Ü.C.; Gezer, E.D.; Temiz, A. Some lignocellulosic wastes used as raw material in cultivation of the Pleurotus ostreatus culture mushroom. Process Biochem. 2002, 38, 301–306. [Google Scholar] [CrossRef]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise review on the molecular structure and function relationship of β-glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef]
- Maheshwari, G.; Sowrirajan, S.; Joseph, B. β-glucan, a dietary fiber in effective prevention of lifestyle diseases—An insight. Bioact. Carbohydr. Diet. Fibre 2019, 19, 100187. [Google Scholar] [CrossRef]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-glucan in foods and ts physiological functions. J. Nutr. Sci. Vitaminol. 2018, 64, 8–17. [Google Scholar] [CrossRef]
- Wang, Q.; Sheng, X.; Shi, A.; Hu, H.; Yang, Y.; Liu, L.; Fei, L.; Liu, H. β-Glucans: Relationships between modification, conformation and functional Activities. Molecules 2017, 22, 257. [Google Scholar] [CrossRef]
- Melanouri, E.-M.; Dedousi, M.; Diamantopoulou, P. Cultivating Pleurotus ostreatus and Pleurotus eryngii mushroom strains on agro-industrial residues in solid-state fermentation. Part II: Effect on productivity and quality of carposomes. Carbon Resour. Convers. 2022, 5, 52–60. [Google Scholar] [CrossRef]
- Philippoussis, A.N.; Diamantopoulou, P.; Zervakis, G. Monitoring of mycelial growth and fructification on Lentinula edodes on several lignocellulosic residues. In Mushroom Biology and Mushroom Products; Sánchez, S.E., Hverta, G., Montiel, E., Eds.; Universidad Auto´noma del Estado de Mocelos: Cuernavaca, Mexico, 2003. [Google Scholar]
- Diamantopoulou, P.; Philippoussis, A. Production attributes of Agaricus bisporus white and off-white strains and the effect of calcium chloride irrigation on productivity and quality. Sci. Hortic. 2001, 91, 379–391. [Google Scholar] [CrossRef]
- Manolopoulou, E.; Philippoussis, A.; Lambrinos, G.; Diamantopoulou, P. Evaluation of productivity and postharvest quality during storage of five Agaricus bisporus strains. J. Food Qual. 2007, 30, 646–663. [Google Scholar] [CrossRef]
- Dias, E.S.; Zied, D.C.; Rinker, D.L. Physiologic response of Agaricus subrufescens using different casing materials and practices applied in the cultivation of Agaricus bisporus. Fungal Biol. 2013, 117, 569–575. [Google Scholar] [CrossRef]
- Llarena-Hernández, C.R.; Largeteau, M.L.; Ferrer, N.; Regnault-Roger, C.; Savoie, J.-M. Optimization of the cultivation conditions for mushroom production with European wild strains of Agaricus subrufescens and Brazilian cultivars: Optimization of the cultivation conditions of A. subrufescens. J. Sci. Food Agric. 2014, 94, 77–84. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Papanikolaou, S.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Patterns of major metabolites biosynthesis by different mushroom fungi grown on glucose-based submerged cultures. Bioprocess Biosyst. Eng. 2014, 37, 1385–1400. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y.; Blackburn, J.W. Batch stage study of lipid production from crude glycerol derived from yellow grease or animal fats through microalgal fermentation. Bioresour. Technol. 2010, 101, 6745–6750. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sarantou, S.; Stoforos, N.G.; Kalantzi, O.; Papanikolaou, S. Biotechnological valorization of biodiesel-derived glycerol: Trials with the non-conventional yeasts Yarrowia lipolytica and Rhodosporidium sp. Carbon Resour. Convers. 2021, 4, 61–75. [Google Scholar] [CrossRef]
- AFNOR (Association Française de Normalisation). Recueil de Normes Françaises. Produits Agricoles Alimentaires: Directives Générales Pour le Dosage de L’azote Avec Minéralisation Selon la Méthode de Kjedahl; AFNOR: Paris, France, 1984. [Google Scholar]
- Allen, S.E. (Ed.) Chemical Analysis of Ecological Materials; Blackwell Scientific: Oxford, UK, 1974. [Google Scholar]
- Kalra, Y.P.; Maynard, D.G. Methods Manual for Forest Soil and Plant Analysis; Information Report, No. NOR-X-319; Forestry Canada, Northwest Region, Northern Forestry Centre: Edmonton, AB, Canada, 1991. [Google Scholar]
- Barros, L.; Cruz, T.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C.F.R. Wild and commercial mushrooms as source of nutrients and nutraceuticals. Food Chem. Toxicol. 2008, 46, 2742–2747. [Google Scholar] [CrossRef]
- ISO 9936:2016; Animal and Vegetable Fats and Oils—Determination of Tocopherol and Tocotrienol Contents by High-Performance Liquid Chromatography. ISO: Geneva, Switzerland, 2016.
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Boonsong, S.; Klaypradit, W.; Wilaipun, P. Antioxidant activities of extracts from five edible mushrooms using different extractants. Agric. Nat. Resour. 2016, 50, 89–97. [Google Scholar] [CrossRef]
- Dilfy, S.H.; Hanawi, M.J.; Al-bideri, A.W.; Jalil, A.T. Determination of chemical composition of cultivated mushrooms in Iraq with Spectrophotometrically and High Performance Liquid Chromatographic. J. Green Eng. 2020, 10, 6200–6216. [Google Scholar]
- Reis, F.S.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Chemical composition and nutritional value of the most widely appreciated cultivated mushrooms: An inter-species comparative study. Food Chem. Toxicol. 2012, 50, 191–197. [Google Scholar] [CrossRef]
- Ulziijargal, E.; Mau, J.-L. Nutrient compositions of culinary-medicinal mushroom fruiting bodies and mycelia. Int. J. Med. Mushrooms 2011, 13, 343–349. [Google Scholar] [CrossRef]
- Irshad, A.; Tahir, A.; Sharif, S.; Khalid, A.; Ali, S.; Naz, A.; Sadia, H.; Ameen, A. Determination of nutritional and biochemical composition of selected Pleurotus spps. BioMed Res. Int. 2023, 2023, 8150909. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Mountzouris, K.C.; Chatzipavlidis, I.; Zervakis, G. Bioconversion of lignocellulosic residues by Agrocybe cylindracea and Pleurotus ostreatus mushroom fungi—Assessment of their effect on the final product and spent substrate properties. Food Chem. 2014, 161, 127–135. [Google Scholar] [CrossRef]
- Mau, J.-L.; Beelman, R.B.; Ziegler, G.R.; Royse, D.J. Effect of nutrient supplementation on flavor, quality, and shelf life of the cultivated mushroom, Agaricus bisporus. Mycologia 1991, 83, 142–149. [Google Scholar] [CrossRef]
- Hammond, J.B.W.; Nichols, R. Carbohydrate metabolism in Agaricus bisporus (Lange) Sing.: Changes in soluble carbohydrates during growth of mycelium and sporophore. J. Gen. Microbiol. 1976, 93, 309–320. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Ruthes, A.C.; van Arkel, J.; Chanput, W.; Iacomini, M.; Wichers, H.J.; Van Griensven, L.J. Polysaccharides from Agaricus bisporus and Agaricus brasiliensis show similarities in their structures and their immunomodulatory effects on human monocytic THP-1 cells. BMC Complement. Altern. Med. 2011, 11, 58. [Google Scholar] [CrossRef]
- Sharma, S.K.; Gautam, N. Chemical, bioactive, and antioxidant potential of twenty wild culinary mushroom species. BioMed Res. Int. 2015, 2015, 346508. [Google Scholar] [CrossRef]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spĕvácek, J.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Baeva, E.; Bleha, R.; Lavrova, E.; Sushytskyi, L.; Čopíková, J.; Jablonský, I.; Klouček, P.; Synytsya, A. Polysaccharides from basidiocarps of cultivating mushroom Pleurotus ostreatus: Isolation and structural characterization. Molecules 2019, 24, 2740. [Google Scholar] [CrossRef]
- Baeva, E.; Bleha, R.; Sedliaková, M.; Sushytskyi, L.; Švec, I.; Čopíková, J.; Jablonsky, I.; Klouček, P.; Synytsya, A. Evaluation of the cultivated mushroom Pleurotus ostreatus basidiocarps using Vibration spectroscopy and chemometrics. Appl. Sci. 2020, 10, 8156. [Google Scholar] [CrossRef]
- Atoji-Henrique, K.; Henrique, D.S.; Glória, L.S.; Mazaro, S.M.; Casagrande, M. Influence of substrate composition on beta-glucans production and growth of Ganoderma lucidum. J. Agric. Sci. 2017, 9, 190. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Larou, E.; Mountzouris, K.C.; Zervakis, G.I. Detoxification of olive mill wastewater and bioconversion of olive crop residues into high-value-added biomass by the choice edible mushroom Hericium erinaceus. Appl. Biochem. Biotechnol. 2016, 180, 195–209. [Google Scholar] [CrossRef]
- Manzi, P.; Pizzoferrato, L. Beta-glucans in edible mushrooms. Food Chem. 2000, 68, 315–318. [Google Scholar] [CrossRef]
- Bekiaris, G.; Tagkouli, D.; Koutrotsios, G.; Kalogeropoulos, N.; Zervakis, G.I. Pleurotus mushrooms content in glucans and ergosterol assessed by ATR-FTIR spectroscopy and multivariate analysis. Foods 2020, 9, 535. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Fungal β-glucans and mammalian immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Cunha Zied, D.; Sánchez, J.E.; Noble, R.; Pardo-Giménez, A. Use of spent mushroom substrate in new mushroom crops to promote the transition towards a circular economy. Agronomy 2020, 10, 1239. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, M.; Fang, Z. Valorization of mushroom by-products: A review. J. Sci. Food Agric. 2022, 102, 5593–5605. [Google Scholar] [CrossRef]
- Bak, W.C.; Park, J.H.; Park, Y.A.; Ka, K.H. Determination of glucan contents in the fruiting bodies and mycelia of Lentinula edodes Cultivars. Mycobiology 2014, 42, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Avni, S.; Ezove, N.; Hanani, H.; Yadid, I.; Karpovsky, M.; Hayby, H.; Gover, O.; Hadar, Y.; Schwartz, B.; Danay, O. Olive mill waste enhances α-glucan content in the edible mushroom Pleurotus eryngii. Int. J. Mol. Sci. 2017, 18, 1564. [Google Scholar] [CrossRef] [PubMed]
- Hoa, H.T.; Wang, C.-L.; Wang, C.-H. The effects of different substrates on the growth, yield, and nutritional composition of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Tolera, K.D.; Abera, S. Nutritional quality of Oyster mushroom (Pleurotus ostreatus) as affected by osmotic pretreatments and drying methods. Food Sci. Nutr. 2017, 5, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Akyüz, M.; Kirbag, S. Nutritive Value of Pleurotus eryngii (DC. ex Fr.) Quel. var. eryngii grown on various agro-wastes. Philipp. Agric. 2009, 92, 327–331. [Google Scholar]
- Delmas, J.; Poitou, N. Nitrogen compounds and more especially amino acids in cultivated mushrooms and in composts. Mushroom Sci. 1965, 6, 193–202. [Google Scholar]
- Tshinyangu, K.K.; Hennebert, G.L. Protein and chitin nitrogen contents and protein content in Pleurotus ostreatus var. columbinus. Food Chem. 1996, 57, 223–227. [Google Scholar] [CrossRef]
- Wang, D.; Sakoda, A.; Suzuki, M. Biological efficiency and nutritional value of Pleurotus ostreatus cultivated on spent beer grain. Bioresour. Technol. 2001, 78, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Dinssa, B.; Temesgen, S.; Mosisa, W. Effect of substrate (waste paper, leaves of P. juliflora and sugar cane bagasse) on nutritional composition of P. ostreatus at Oda Bultum University, Chiro, Ethiopia. Am. J. Agric. For. 2020, 8, 46. [Google Scholar]
- Sardar, H.; Ali, M.A.; Anjum, M.A.; Nawaz, F.; Hussain, S.; Naz, S.; Karimi, S.M. Agro-industrial residues influence mineral elements accumulation and nutritional composition of king oyster mushroom (Pleurotus eryngii). Sci. Hortic. 2017, 225, 327–334. [Google Scholar] [CrossRef]
- Rapior, S.; Mousain, D.; Plassard, C.; Andary, C.; Salsac, L. Influence of nitrogen source on growth of Cortinarius orellanus and on accumulation of nitrogen and phosphorus in mycelium. Trans. Br. Mycol. Soc. 1988, 90, 181–185. [Google Scholar] [CrossRef]
- Nasiri, F.; Tarzi, B.G.; Bassiri, A.R.; Hoseini, S.E.; Aminafshar, M. Comparative study on the main chemical composition of button mushroom’s (Agaricus bisporus) cap and stipe. J. Food Biosci. Technol. 2013, 3, 41–48. [Google Scholar]
- Crisan, E.V.; Sands, A. Edible mushrooms: Nutritional value. In The Biology and Cultivation of Edible Mushrooms; Chang, S.T., Hayes, W.A., Eds.; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Alam, N.; Amin, R.; Khan, A.; Ara, I.; Shim, M.-J.; Lee, M.-W.; Lee, T.-S. Nutritional analysis of cultivated mushrooms in Bangladesh—Pleurotus ostreatus, Pleurotus sajor-caju, Pleurotus florida and Calocybe indica. Mycobiology 2008, 36, 228–232. [Google Scholar] [CrossRef]
- Teklit, G. Chemical composition and nutritional value of the most widely used mushrooms cultivated in Mekelle Tigray Ethiopia. J. Nutr. Food Sci. 2015, 5, 1000408. [Google Scholar]
- Tsai, S.; Tsai, H.; Mau, J. Non-volatile taste components of Agaricus blazei, Agrocybe cylindracea and Boletus edulis. Food Chem. 2008, 107, 977–983. [Google Scholar] [CrossRef]
- Huang, L.C.; Mau, J.L. Antioxidant properties and polysaccharide composition analysis of Antrodia camphorata and Agaricus blazei. Master’s Thesis, National Chung-Hsing University, Taichung, Taiwan, 2000. [Google Scholar]
- Diamantopoulou, P.; Papanikolaou, S.; Katsarou, E.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Mushroom polysaccharides and lipids synthesized in liquid agitated and static cultures. Part II: Study of Volvariella volvacea. Appl. Biochem. Biotechnol. 2012, 167, 1890–1906. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Papanikolaou, S.; Aggelis, G.; Philippoussis, A. Adaptation of Volvariella volvacea metabolism in high carbon to nitrogen ratio media. Food Chem. 2016, 196, 272–280. [Google Scholar] [CrossRef]
- Bengü, A. The fatty acid composition in some economic and wild edible mushrooms in Turkey. Prog. Nutr. 2020, 22, 185–192. [Google Scholar] [CrossRef]
- Kayode, R.M.O.; Olakulehin, T.F.; Adedeji, B.S.; Ahmed, O.; Aliyu, T.H.; Badmos, A.H.A. Evaluation of amino acid and fatty acid profiles of commercially cultivated oyster mushroom (Pleurotus sajor-caju) grown on gmelina wood waste. Niger. Food J. 2015, 33, 18–21. [Google Scholar] [CrossRef]
- Rosemary-Kinge, T.; Adi, E.; Mih, A.M.; Ache, N.; Nji, T. Effect of substrate on the growth, nutritional and bioactive components of Pleurotus ostreatus and Pleurotus florida. Afr. J. Biotechnol. 2016, 15, 1476–1486. [Google Scholar] [CrossRef]
- Yamagishi, K.; Folsom, A.R.; Steffen, L.M.; ARIC Study Investigators. Plasma fatty acid composition and incident ischemic stroke in middle-aged adults: The Atherosclerosis Risk in Communities (ARIC) Study. Cerebrovasc. Dis. 2013, 36, 38–46. [Google Scholar] [CrossRef]
- Cardoso, R.V.C.; Carocho, M.; Fernandes, Â.; Zied, D.C.; Cobos, J.D.V.; González-Paramás, A.M.; Ferreira, I.C.F.R.; Barros, L. Influence of calcium silicate on the chemical properties of Pleurotus ostreatus var. florida (Jacq.) P. Kumm. J. Fungi 2020, 6, 299. [Google Scholar]
- Elmastas, M.; Isildak, O.; Turkekul, I.; Temur, N. Determination of antioxidant activity and antioxidant compounds in wild edible mushrooms. J. Food Compos. Anal. 2007, 20, 337–345. [Google Scholar] [CrossRef]
- Da Paz, M.F.; Breyer, C.A.; Longhi, R.F.; Oviedo, M.S.V.P. Determining the basic composition and total phenolic compounds of Pleurotus sajor-caju cultivated in three different substrates by solid state bioprocess. J. Biotechnol. Biodivers. 2012, 3, 11–14. [Google Scholar] [CrossRef]
- Wong, F.; Chai, T.; Tan, S.; Yong, A. Evaluation of bioactivities and phenolic content of selected edible mushrooms in Malaysia. Trop. J. Pharm. Res. 2014, 12, 1011. [Google Scholar] [CrossRef]
- Elhusseiny, S.M.; El-Mahdy, T.S.; Awad, M.F.; Elleboudy, N.S.; Farag, M.M.S.; Aboshanab, K.M.; Yassien, M.A. Antiviral, cytotoxic, and antioxidant activities of three edible Agaricomycetes mushrooms: Pleurotus columbinus, Pleurotus sajor-caju, and Agaricus bisporus. J. Fungi 2021, 7, 645. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Yanni, A.E.; Koutrotsios, G.; Aloupi, M. Bioactive microconstituents and antioxidant properties of wild edible mushrooms from the island of Lesvos, Greece. Food Chem. Toxicol. 2013, 55, 378–385. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Stathopoulos, P.; Kaliora, A.C.; Zervakis, G.I. Bioactive compounds and antioxidant activity exhibit high intraspecific variability in Pleurotus ostreatus mushrooms and correlate well with cultivation performance parameters. World J. Microbiol. Biotechnol. 2017, 33, 98. [Google Scholar] [CrossRef]
- Butkhup, L.; Samappito, W.; Jorjong, S. Evaluation of bioactivities and phenolic contents of wild edible mushrooms from northeastern Thailand. Food Sci. Biotechnol. 2018, 27, 193–202. [Google Scholar] [CrossRef]
- Strapáč, I.; Kuruc, M.; Baranová, M. Determination of antioxidant parameters of Pleurotus mushrooms growing on different wood substrates. Folia Vet. 2017, 61, 53–58. [Google Scholar] [CrossRef]
- Manzi, P.; Gambelli, L.; Marconi, S.; Vivanti, V.; Pizzoferrato, L. Nutrients in edible mushrooms: An inter-species comparative study. Food Chem. 1999, 65, 477–482. [Google Scholar] [CrossRef]
- Kalač, P.; Burda, J.; Stašková, I. Concentrations of lead, cadmium, mercury and copper in mushrooms in the vicinity of a lead smelter. Sci. Total Environ. 1991, 105, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.; Grewal, R.; Goyal, R.K. Vitamin and mineral content of Agaricus bisporus (white button) and Pleurotus sajor-caju (dhingri) mushrooms. Int. J. Food Sci. Nutr. 2020, 5, 100–102. [Google Scholar]
- Chong, K.S.; YEE, C.F.; Lee, J.; Atong, M. Nutritional properties of some edible wild mushrooms in Sabah. J. Appl. Sci. 2007, 7, 2216–2221. [Google Scholar]
- Demirbaş, A. Heavy metal bioaccumulation by mushrooms from artificially fortified soils. Food Chem. 2001, 74, 293–301. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungi/ Compost | Moisture (%) | Ash (%) | IPSs (%, w/w) | Proteins (%, w/w) | Lipids (%, w/w) | TPC (mg GAE/g) | DPPH∙ (mg trolox/g) | ABTS∙+ (mg trolox/g) | FRAP (mg trolox/g) | |
|---|---|---|---|---|---|---|---|---|---|---|
| A. bisporus (C/N = 10) | c | 87.64 ± 3.59 | 9.21 ± 0.86 | 36.68 ± 0.65 | 39.58 ± 2.97 | 2.68 ± 0.10 | 12.59 ± 1.16 | 0.96 ± 0.04 | 1.31 ± 0.21 | 2.23 ± 0.24 |
| p | 88.65 ± 3.42 | 9.20 ± 1.08 | 39.08 ± 0.75 | 40.51 ± 3.54 | 2.74 ± 0.22 | 13.12 ± 0.87 | 0.99 ± 0.07 | 1.21 ± 0.10 | 2.25 ± 0.37 | |
| s | 81.95 ± 2.65 | 9.12 ± 0.96 | 31.36 ± 1.64 | 32.04 ± 2.64 | 2.36 ± 0.14 | 11.23 ± 0.66 | 1.04 ± 0.11 | 1.30 ± 0.06 | 2.30 ± 0.12 | |
| A. bisporus (C/N = 13) | c | 89.51 ± 2.55 | 8.55 ± 0.87 | 30.46 ± 1.28 | 36.16 ± 3.50 | 2.74 ± 0.09 | 10.49 ± 0.58 | 0.82 ± 0.09 | 1.20 ± 0.08 | 1.85 ± 0.09 |
| p | 87.62 ± 2.13 | 8.50 ± 0.88 | 37.30 ± 0.80 | 38.98 ± 2.70 | 2.92 ± 0.13 | 10.80 ± 0.42 | 0.83 ± 0.04 | 0.80 ± 0.05 | 1.82 ± 0.11 | |
| s | 80.94 ± 2.11 | 8.46 ± 1.02 | 22.46 ± 0.89 | 29.89 ± 3.28 | 2.08 ± 0.03 | 7.85 ± 0.61 | 0.75 ± 0.02 | 0.88 ± 0.03 | 0.74 ± 0.20 | |
| A. subrufescens (C/N = 10) | c | 86.34 ± 2.82 | 8.11 ± 1.13 | 32.44 ± 0.32 | 43.59 ± 2.14 | 3.54 ± 0.20 | 13.49 ± 0.65 | 1.10 ± 0.07 | 1.58 ± 0.05 | 2.48 ± 0.18 |
| p | 86.12 ± 2.61 | 8.06 ± 1.14 | 30.02 ± 0.21 | 44.26 ± 1.87 | 3.70 ± 0.12 | 13.00 ± 0.47 | 0.97 ± 0.09 | 1.51 ± 0.13 | 2.16 ± 0.09 | |
| s | 82.91 ± 2.37 | 8.12 ± 1.20 | 27.62 ± 0.51 | 40.12 ± 1.21 | 2.66 ± 0.08 | 12.99 ± 0.52 | 1.00 ± 0.12 | 1.61 ± 0.21 | 2.22 ± 0.07 | |
| A. subrufescens (C/N = 13) | c | 87.58 ± 3.89 | 8.35 ± 2.08 | 29.96 ± 0.98 | 33.22 ± 2.21 | 3.18 ± 0.26 | 15.49 ± 0.88 | 2.10 ± 0.16 | 1.78 ± 0.08 | 2.68 ± 0.12 |
| p | 87.62 ± 4.02 | 8.10 ± 1.24 | 27.25 ± 0.83 | 40.50 ± 3.22 | 3.44 ± 0.18 | 16.89 ± 0.92 | 2.97 ± 0.10 | 1.71 ± 0.09 | 2.56 ± 0.35 | |
| s | 80.34 ± 3.23 | 8.36 ± 1.02 | 22.50 ± 1.19 | 25.40 ± 2.12 | 2.60 ± 0.25 | 13.99 ± 0.67 | 2.00 ± 0.08 | 0.73 ± 0.06 | 2.42 ± 0.29 |
| Carbohydrates (%, w/w of Total IPSs) | |||||
|---|---|---|---|---|---|
| Substrates/Fungi | Glucose | Fructose | Mannitol | Arabitol | |
| P. ostreatus | |||||
| WS * | 144 c | 70.5 ± 4.8 | 20.2 ± 1.7 | 9.3 ± 0.9 | nd ** |
| 144 p | 64.2 ± 2.3 | 25.3 ± 2.3 | 10.5 ± 0.3 | nd | |
| 144 s | 84.4 ± 3.9 | 15.6 ± 1.7 | nd | nd | |
| 150 c | 72.3 ± 4.2 | 16.5 ± 0.9 | 11.2 ± 1.4 | nd | |
| 150 p | 68.7 ± 4.6 | 15.3 ± 0.6 | 10.2 ± 0.8 | 5.8 ± 0.4 | |
| 150 s | 85.4 ± 3.8 | 14.6 ± 1.3 | nd | nd | |
| BOS | 144 c | 72.1 ± 3.9 | 17.4 ± 0.6 | 10.5 ± 0.7 | nd |
| 144 p | 71.5 ± 4.0 | 18.7 ± 0.3 | 9.8 ± 0.4 | nd | |
| 144 s | 75.4 ± 3.2 | 10.5 ± 0.1 | 14.1 ± 1.0 | nd | |
| 150 c | 73.4 ± 2.1 | 16.5 ± 0.9 | 10.1 ± 0.8 | nd | |
| 150 p | 67.5 ± 1.8 | 15.8 ± 0.8 | 9.7 ± 0.4 | 7.2 ± 0.3 | |
| 150 s | 82.2 ± 3.4 | 12.4 ± 1.1 | 5.4 ± 0.5 | nd | |
| RB | 144 c | 73.5 ± 4.5 | 13.4 ± 0.2 | 13.1 ± 0.9 | nd |
| 144 p | 74.3 ± 3.3 | 13.6 ± 0.9 | 12.1 ± 1.2 | nd | |
| 144 s | 76.1 ± 2.1 | 13.3 ± 0.7 | 10.6 ± 0.7 | nd | |
| 150 c | 72.4 ± 4.2 | 12.2 ± 0.4 | 10.4 ± 0.6 | 5.6 ± 0.2 | |
| 150 p | 70.2 ± 3.1 | 10.7 ± 0.3 | 9.1 ± 0.8 | 10.0 ± 0.4 | |
| 150 s | 70.6 ± 2.8 | 12.6 ± 0.9 | 9.7 ± 0.3 | 7.1 ± 0.3 | |
| BWS | 144c | 73.8 ± 2.5 | 14.7 ± 0.9 | 11.5 ± 0.9 | nd |
| 144 p | 70.2 ± 2.0 | 12.4 ± 0.7 | 17.4 ± 0.8 | nd | |
| 144 s | 80.5 ± 1.8 | 12.6 ± 1.2 | 6.9 ± 0.9 | nd | |
| 150 c | 69.2 ± 2.7 | 13.5 ± 0.9 | 9.3 ± 0.6 | 8.0 ± 0.5 | |
| 150 p | 70.4 ± 3.2 | 13.4 ± 0.8 | 16.2 ± 0.9 | nd | |
| 150 s | 72.6 ± 2.9 | 14.7 ± 0.4 | 12.7 ± 0.8 | nd | |
| CR | 144 c | 73.9 ± 3.6 | 15.9 ± 0.7 | 10.2 ± 1.1 | nd |
| 144 p | 72.7 ± 3.8 | 16.8 ± 1.3 | 10.5 ± 0.3 | nd | |
| 144 s | 81.4 ± 3.1 | 11.4 ± 0.9 | 7.2 ± 0.8 | nd | |
| 150 c | 73.6 ± 2.7 | 14.1 ± 0.2 | 6.3 ± 0.9 | 6.0 ± 0.2 | |
| 150 p | 70.8 ± 1.9 | 11.3 ± 0.5 | 9.5 ± 0.3 | 8.4 ± 0.4 | |
| 150 s | 80.5 ± 3.9 | 12.8 ± 0.8 | 6.7 ± 0.7 | nd | |
| P. eryngii | |||||
| WS | 166 c | 78.7 ± 2.7 | 15.2 ± 1.5 | 6.1 ± 0.4 | nd |
| 166 p | 76.6 ± 4.1 | 13.3 ± 1.1 | 10.1 ± 0.9 | nd | |
| 166 s | 81.6 ± 2.2 | 11.4 ± 0.9 | 7.0 ± 0.5 | nd | |
| 173-6 c | 72.1 ± 3.2 | 16.7 ± 0.8 | 11.2 ± 0.8 | nd | |
| 173-6 p | 70.6 ± 2.7 | 16.6 ± 1.2 | 12.8 ± 1.1 | nd | |
| 173-6 s | 75.2 ± 2.9 | 14.5 ± 0.5 | 10.3 ± 0.8 | nd | |
| BOS | 166 c | 74.3 ± 3.1 | 11.4 ± 0.7 | 7.3 ± 0.4 | 7.0 ± 0.5 |
| 166 p | 77.6 ± 3.7 | 13.6 ± 1.1 | 8.8 ± 0.5 | nd | |
| 166 s | 82.6 ± 2.8 | 10.4 ± 0.8 | 7.0 ± 0.1 | nd | |
| 173-6 c | 73.6 ± 2.2 | 15.5 ± 1.6 | 10.9 ± 1.0 | nd | |
| 173-6 p | 71.8 ± 2.7 | 15.9 ± 0.7 | 12.3 ± 1.3 | nd | |
| 173-6 s | 75.7 ± 3.6 | 14.6 ± 1.3 | 9.7 ± 0.6 | nd | |
| RB | 166 c | 75.3 ± 4.2 | 13.5 ± 0.8 | 11.2 ± 0.4 | n.d |
| 166 p | 68.7 ± 3.7 | 15.5 ± 0.4 | 15.8 ± 1.4 | n.d | |
| 166 s | 80.3 ± 3.9 | 10.5 ± 0.9 | 9.2 ± 0.2 | n.d | |
| 173-6 c | 72.6 ± 2.2 | 12.7 ± 1.4 | 14.7 ± 0.8 | n.d | |
| 173-6 p | 69.9 ± 3.5 | 13.4 ± 1.4 | 16.7 ± 1.7 | n.d | |
| 173-6 s | 78.8 ± 4.5 | 12.6 ± 1.2 | 8.6 ± 0.9 | n.d | |
| BWS | 166 c | 76.6 ± 3.1 | 12.6 ± 0.7 | 10.8 ± 0.4 | n.d |
| 166 p | 73.2 ± 3.8 | 10.0 ± 0.5 | 7.6 ± 0.2 | 9.2 ± 0.6 | |
| 166 s | 79.9 ± 2.4 | 12.2 ± 0.9 | 7.9 ± 0.2 | n.d | |
| 173-6 c | 75.6 ± 2.8 | 13.4 ± 1.0 | 11.0 ± 1.1 | n.d | |
| 173-6 p | 68.5 ± 1.2 | 13.7 ± 2.4 | 8.2 ± 0.6 | 9.6 ± 0.1 | |
| 173-6 s | 78.6 ± 2.4 | 12.2 ± 2.1 | 9.2 ± 0.3 | n.d | |
| CR | 166 c | 73.9 ± 2.9 | 15.8 ± 1.1 | 10.3 ± 0.7 | n.d |
| 166 p | 65.4 ± 2.1 | 16.6 ± 1.4 | 8.4 ± 0.4 | 9.6 ± 0.8 | |
| 166 s | 79.9 ± 4.3 | 10.6 ± 0.8 | 9.5 ± 0.9 | n.d | |
| 173-6 c | 72.6 ± 2.7 | 13.6 ± 1.3 | 13.8 ± 0.7 | n.d | |
| 173-6 p | 70.1 ± 2.6 | 9.7 ± 0.6 | 10.7 ± 0.9 | 9.5 ± 0.5 | |
| 173-6 s | 78.1 ± 3.0 | 12.4 ± 1.1 | 9.5 ± 0.6 | n.d | |
| Agaricus spp. | |||||
| A. bisporus (C/N = 10) | c | 63.3 ± 3.6 | 23.2 ± 0.4 | 13.5 ± 0.9 | nd |
| p | 63.5 ± 2.2 | 24.2 ± 2.3 | 12.3 ± 0.6 | nd | |
| s | 67.4 ± 3.2 | 23.3 ± 0.9 | 9.3 ± 0.3 | nd | |
| A. bisporus (C/N = 13) | c | 63.8 ± 4.2 | 22.9 ± 1.6 | 13.3 ± 0.8 | nd |
| p | 61.3 ± 2.7 | 21.5 ± 1.8 | 9.1 ± 0.8 | 8.1 ± 0.7 | |
| s | 65.4 ± 2.9 | 23.9 ± 2.3 | 10.7 ± 0.4 | nd | |
| A. subrufescens (C/N = 10) | c | 65.5 ± 2.4 | 10.4 ± 1.3 | 13.5 ± 0.7 | 10.6 ± 0.9 |
| p | 64.4 ± 3.8 | 12.0 ± 0.5 | 11.9 ± 0.4 | 11.7 ± 0.4 | |
| s | 67.1 ± 3.1 | 22.7 ± 0.8 | 10.2 ± 0.2 | nd | |
| A. subrufescens (C/N = 13) | c | 64.2 ± 3.4 | 11.5 ± 1.7 | 15.1 ± 0.6 | 9.0 ± 0.5 |
| p | 63.9 ± 2.7 | 11.9 ± 1.2 | 12.3 ± 0.4 | 12.4 ± 1.1 | |
| s | 65.1 ± 1.9 | 10.4 ± 0.9 | 9.5 ± 0.7 | 14.9 ± 0.6 | |
| Glucans (%, w/w of Total IPSs) | ||||
|---|---|---|---|---|
| Substrates/Fungi | Total | α-Glucans | β-Glucans | |
| P. ostreatus | ||||
| WS * | 144 c | 36.28 ± 3.65 | 10.39 ± 2.65 | 25.89 ± 2.87 |
| 144 p | 38.23 ± 3.50 | 12.41 ± 2.40 | 25.82 ± 3.54 | |
| 144 s | 46.35 ± 2.87 | 4.14 ± 0.65 | 42.21 ± 3.24 | |
| 150 c | 47.94 ± 2.67 | 12.73 ± 1.50 | 35.22 ± 2.70 | |
| 150 p | 49.61 ± 3.14 | 11.25 ± 1.25 | 38.36 ± 2.65 | |
| 150 s | 56.51 ± 4.35 | 12.36 ± 1.64 | 44.15 ± 3.40 | |
| BOS | 144 c | 31.43 ± 3.54 | 5.79 ± 0.47 | 25.64 ± 3.35 |
| 144 p | 21.47 ± 3.50 | 3.24 ± 0.27 | 18.23 ± 2.64 | |
| 144 s | 35.54 ± 2.98 | 8.23 ± 1.02 | 27.31 ± 3.50 | |
| 150 c | 37.08 ± 3.40 | 10.72 ± 1.60 | 26.36 ± 3.60 | |
| 150 p | 33.64 ± 3.21 | 9.23 ± 1.02 | 24.41 ± 3.40 | |
| 150 s | 38.12 ± 2.67 | 12.70 ± 1.84 | 25.42 ± 4.02 | |
| RB | 144 c | 39.33 ± 2.56 | 7.10 ± 1.01 | 32.23 ± 2.50 |
| 144 p | 32.06 ± 2.41 | 4.54 ± 1.30 | 27.52 ± 2.60 | |
| 144 s | 44.36 ± 2.60 | 9.32 ± 1.23 | 35.04 ± 3.40 | |
| 150 c | 44.39 ± 2.41 | 8.23 ± 1.30 | 36.16 ± 3.21 | |
| 150 p | 43.13 ± 2.50 | 7.93 ± 1.08 | 35.20 ± 3.40 | |
| 150 s | 48.65 ± 3.02 | 9.29 ± 1.20 | 39.36 ± 2.97 | |
| BWS | 144 c | 34.21 ± 2.54 | 4.03 ± 1.02 | 30.18 ± 2.50 |
| 144 p | 31.91 ± 2.60 | 4.65 ± 1.20 | 27.26 ± 2.60 | |
| 144 s | 44.55 ± 3.20 | 6.32 ± 1.54 | 38.23 ± 2.40 | |
| 150 c | 38.44 ± 2.31 | 5.81 ± 1.34 | 32.63 ± 2.64 | |
| 150 p | 35.90 ± 2.40 | 5.63 ± 1.40 | 30.27 ± 2.60 | |
| 150 s | 44.08 ± 3.20 | 6.04 ± 1.50 | 38.04 ± 3.04 | |
| CR | 144 c | 40.84 ± 3.41 | 11.06 ± 1.58 | 29.78 ± 2.80 |
| 144 p | 37.97 ± 2.65 | 11.72 ± 1.47 | 26.25 ± 2.64 | |
| 144 s | 49.59 ± 2.70 | 11.38 ± 1.80 | 38.21 ± 2.70 | |
| 150 c | 41.05 ± 2.15 | 15.38 ± 1.40 | 25.67 ± 3.04 | |
| 150 p | 37.12 ± 2.34 | 14.32 ± 1.65 | 22.80 ± 3.14 | |
| 150 s | 53.53 ± 3.41 | 16.89 ± 1.76 | 36.64 ± 2.64 | |
| P. eryngii | ||||
| WS | 166 c | 49.30 ± 3.08 | 7.70 ± 1.30 | 41.60 ± 2.64 |
| 166 p | 48.50 ± 2.74 | 10.25 ± 1.58 | 38.25 ± 2.58 | |
| 166 s | 53.63 ± 3.70 | 10.29 ± 1.36 | 43.34 ± 2.70 | |
| 173-6 c | 52.51 ± 3.30 | 10.71 ± 1.80 | 41.80 ± 2.87 | |
| 173-6 p | 50.51 ± 3.51 | 10.50 ± 1.56 | 40.01 ± 2.70 | |
| 173-6 s | 64.21 ± 3.45 | 14.26 ± 1.47 | 49.95 ± 2.95 | |
| BOS | 166 c | 48.75 ± 3.34 | 8.77 ± 1.54 | 39.98 ± 2.85 |
| 166 p | 47.77 ± 2.65 | 9.52 ± 1.63 | 38.25 ± 2.95 | |
| 166 s | 61.92 ± 3.66 | 14.52 ± 1.80 | 47.40 ± 2.85 | |
| 173-6 c | 52.36 ± 2.60 | 10.68 ± 1.65 | 41.68 ± 2.97 | |
| 173-6 p | 46.69 ± 2.54 | 9.65 ± 1.02 | 37.04 ± 2.75 | |
| 173-6 s | 55.82 ± 2.40 | 13.11 ± 2.34 | 42.71 ± 2.64 | |
| RB | 166 c | 38.02 ± 2.64 | 8.17 ± 1.25 | 29.88 ± 3.50 |
| 166 p | 35.37 ± 2.37 | 8.03 ± 1.60 | 27.34 ± 2.65 | |
| 166 s | 47.12 ± 2.40 | 8.87 ± 1.41 | 38.25 ± 3.40 | |
| 173-6 c | 38.76 ± 2.30 | 6.73 ± 1.34 | 32.04 ± 2.50 | |
| 173-6 p | 31.91 ± 2.41 | 2.94 ± 0.50 | 28.97 ± 3.04 | |
| 173-6 s | 51.98 ± 3.66 | 6.84 ± 0.94 | 45.14 ± 2.94 | |
| BWS | 166 c | 37.69 ± 2.68 | 9.84 ± 1.25 | 27.85 ± 2.85 |
| 166 p | 36.06 ± 2.57 | 9.01 ± 1.30 | 27.05 ± 2.60 | |
| 166 s | 42.92 ± 3.50 | 12.52 ± 1.54 | 30.40 ± 3.24 | |
| 173-6 c | 37.88 ± 3.24 | 11.23 ± 1.80 | 26.65 ± 3.05 | |
| 173-6 p | 30.25 ± 3.60 | 10.01 ± 1.20 | 20.24 ± 3.14 | |
| 173-6 s | 40.47 ± 3.50 | 11.24 ± 1.34 | 29.23 ± 2.90 | |
| CR | 166 c | 45.04 ± 2.41 | 7.70 ± 0.90 | 37.44 ± 3.40 |
| 166 p | 40.86 ± 2.05 | 7.65 ± 0.67 | 33.21 ± 4.02 | |
| 166 s | 52.15 ± 2.47 | 8.84 ± 1.02 | 43.31 ± 4.51 | |
| 173-6 c | 58.34 ± 2.64 | 13.30 ± 1.80 | 45.04 ± 3.70 | |
| 173-6 p | 57.34 ± 2.07 | 12.20 ± 1.60 | 45.14 ± 3.51 | |
| 173-6 s | 62.59 ± 2.32 | 14.26 ± 1.54 | 48.33 ± 2.84 | |
| Agaricus spp. | ||||
| A. bisporus (C/N = 10) | c | 15.03 ± 1.64 | 4.23 ± 1.23 | 10.80 ± 1.57 |
| p | 10.26 ± 1.37 | 2.04 ± 0.56 | 8.22 ± 1.20 | |
| s | 24.39 ± 1.25 | 8.47 ± 1.40 | 13.91 ± 1.60 | |
| A. bisporus (C/N = 13) | c | 12.27 ± 1.20 | 2.27 ± 0.24 | 10.00 ± 1.23 |
| p | 10.12 ± 1.41 | 2.10 ± 0.65 | 8.02 ± 1.05 | |
| s | 20.10 ± 1.26 | 7.24 ± 1.32 | 12.86 ± 1.60 | |
| A. subrufescens (C/N = 10) | c | 15.89 ± 1.30 | 4.32 ± 0.52 | 11.57 ± 1.40 |
| p | 12.13 ± 1.14 | 2.12 ± 0.65 | 10.01 ± 1.54 | |
| s | 15.74 ± 1.65 | 2.13 ± 1.05 | 12.61 ± 2.04 | |
| A. subrufescens (C/N = 13) | c | 14.58 ± 2.11 | 4.11 ± 0.09 | 11.02 ± 1.89 |
| p | 12.04 ± 1.09 | 2.00 ± 0.65 | 9.87 ± 1.32 | |
| s | 14.97 ± 1.42 | 1.98 ± 0.05 | 11.79 ± 1.87 | |
| Substrate | C14:0 | C15:0 | C15:1 | C16:0 | C18:0 | C18:1 | C18:2 | C20:0 | C22:0 | C24:0 | Other | Poly-unsaturated | U.I. *** | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. ostreatus | ||||||||||||||
| WS * | 144 | 0.26 ± 0.08 | 0.98 ± 0.05 | 0.14 ± 0.05 | 11.1 ± 1.33 | 0.95 ± 0.04 | 9.62 ± 1.04 | 67.3 ± 3.65 | 0.14 ± 0.03 | 0.29 ± 0.05 | 0.84 ± 0.07 | 2.66 | 67.6 | 1.44 |
| 150 | 0.24 ± 0.06 | 0.69 ± 0.07 | 0.12 ± 0.05 | 11.3 ± 1.00 | 1.12 ± 0.05 | 9.50 ± 1.22 | 69.3 ± 3.05 | 0.10 ± 0.02 | 0.22 ± 0.04 | 0.51 ± 0.04 | 2.77 | 69.5 | 1.48 | |
| BOS | 144 | 0.27 ± 0.06 | 1.14 ± 0.10 | 0.26 ± 0.06 | 12.1 ± 1.08 | 0.95 ± 0.03 | 8.32 ± 1.06 | 68.3 ± 2.78 | nd | 0.42 ± 0.04 | nd | 2.42 | 68.5 | 1.45 |
| 150 | 0.30 ± 0.08 | 0.93 ± 0.08 | 0.11 ± 0.06 | 12.1 ± 1.40 | 1.30 ± 0.04 | 6.90 ± 1.24 | 68.3 ± 2.79 | 0.18 ± 0.03 | 0.36 ± 0.05 | 0.57 ± 0.05 | 2.32 | 68.5 | 1.44 | |
| RB | 144 | 0.24 ± 0.07 | 0.75 ± 0.07 | nd ** | 11.6 ± 1.58 | 0.84 ± 0.05 | 8.90 ± 1.31 | 72.4 ± 3.51 | nd | nd | 0.13 ± 0.03 | 2.16 | 72.6 | 1.54 |
| 150 | 0.52 ± 0.09 | 0.57 ± 0.08 | nd | 13.1 ± 0.08 | 1.68 ± 0.05 | 8.60 ± 1.56 | 66.5 ± 3.07 | nd | 0.33 ± 0.05 | 0.26 ± 0.03 | 2.83 | 66.9 | 1.42 | |
| BWS | 144 | 0.26 ± 0.05 | 1.25 ± 0.09 | 0.59 ± 0.09 | 12.3 ± 1.65 | 1.18 ± 0.06 | 8.02 ± 1.34 | 67.8 ± 2.71 | 0.14 ± 0.02 | 0.30 ± 0.04 | 1.15 ± 0.04 | 3.24 | 68.8 | 1.44 |
| 150 | 0.27 ± 0.07 | 1.34 ± 0.11 | 0.55 ± 0.06 | 12.0 ± 1.27 | 1.21 ± 0.05 | 7.97 ± 1.64 | 67.5 ± 2.90 | 0.15 ± 0.03 | 0.35 ± 0.05 | 1.28 ± 0.06 | 3.19 | 68.1 | 1.43 | |
| CR | 144 | 0.40 ± 0.02 | 1.09 ± 0.10 | 0.29 ± 0.03 | 10.7 ± 1.32 | 0.69 ± 0.03 | 7.59 ± 0.02 | 72.0 ± 2.84 | nd | 0.11 ± 0.04 | 0.22 ± 0.05 | 2.97 | 72.4 | 1.45 |
| 150 | 0.43 ± 0.05 | 0.74 ± 0.08 | 0.19 ± 0.06 | 11.6 ± 1.04 | 1.21 ± 0.06 | 10.6 ± 1.74 | 68.4 ± 2.64 | nd | 0.12 ± 0.04 | 0.25 ± 0.06 | 2.02 | 68.6 | 1.48 | |
| P. eryngii | ||||||||||||||
| WS | 166 | 0.53 ± 0.06 | 2.00 ± 0.10 | 0.14 ± 0.02 | 12.8 ± 1.41 | 1.65 ± 0.07 | 20.7 ± 2.22 | 56.2 ± 3.01 | 0.10 ± 0.02 | nd | 0.24 ± 0.06 | 2.86 | 56.4 | 1.33 |
| 173-6 | 0.32 ± 0.05 | 1.21 ± 0.09 | nd | 11.9 ± 1.05 | 2.05 ± 0.05 | 25.6 ± 2.16 | 52.9 ± 2.47 | 0.21 ± 0.03 | 0.23 ± 0.05 | 0.82 ± 0.06 | 1.67 | 53.2 | 1.31 | |
| BOS | 166 | 0.49 ± 0.06 | 1.66 ± 0.10 | nd | 11.7 ± 1.22 | 1.61 ± 0.06 | 18.7 ± 2.40 | 59.1 ± 3.04 | 0.08 ± 0.01 | 0.21 ± 0.04 | 0.42 ± 0.05 | 2.27 | 59.9 | 1.37 |
| 173-6 | 0.34 ± 0.07 | 1.10 ± 0.09 | 0.20 ± 0.05 | 11.5 ± 1.37 | 1.90 ± 0.04 | 22.2 ± 1.65 | 55.7 ± 3.56 | 0.39 ± 0.02 | 0.23 ± 0.05 | 0.84 ± 0.06 | 2.02 | 56.2 | 1.34 | |
| RB | 166 | 0.35 ± 0.07 | 1.52 ± 0.08 | 0.19 ± 0.05 | 12.5 ± 0.09 | 1.57 ± 0.06 | 22.4 ± 1.54 | 55.6 ± 3.54 | 0.18 ± 0.04 | 0.24 ± 0.04 | 0.85 ± 0.05 | 2.54 | 57.4 | 1.34 |
| 173-6 | 0.47 ± 0.05 | 1.64 ± 0.04 | 0.23 ± 0.05 | 11.9 ± 1.03 | 1.63 ± 0.05 | 20.9 ± 1.34 | 55.4 ± 3.67 | 0.22 ± 0.05 | 0.25 ± 0.05 | 0.87 ± 0.04 | 2.37 | 56.8 | 1.32 | |
| BWS | 166 | 0.33 ± 0.06 | 1.67 ± 0.05 | 0.32 ± 0.06 | 12.7 ± 1.10 | 1.85 ± 0.06 | 21.3 ± 1.50 | 58.7 ± 4.02 | 0.17 ± 0.03 | nd | 0.86 ± 0.03 | 2.70 | 57.2 | 1.39 |
| 173-6 | 0.37 ± 0.05 | 1.57 ± 0.06 | 0.10 ± 0.03 | 11.5 ± 1.08 | 1.73 ± 0.04 | 22.7 ± 1.70 | 57.3 ± 4.01 | 0.20 ± 0.04 | 0.21 ± 0.03 | 0.78 ± 0.04 | 1.90 | 57.7 | 1.37 | |
| CR | 166 | 0.54 ± 0.07 | 1.60 ± 0.07 | nd | 11.3 ± 0.90 | 1.93 ± 0.06 | 22.4 ± 2.14 | 53.4 ± 3.61 | 0.27 ± 0.06 | nd | 0.54 ± 0.06 | 2.45 | 55.7 | 1.29 |
| 173-6 | 0.57 ± 0.08 | 1.47 ± 0.04 | nd | 12.6 ± 0.84 | 2.08 ± 0.05 | 23.5 ± 1.70 | 56.7 ± 3.40 | 0.34 ± 0.04 | 0.20 ± 0.04 | 0.47 ± 0.04 | 1.60 | 57.1 | 1.37 | |
| A. bisporus | ||||||||||||||
| C/N = 10 | 0.49 ± 0.08 | 0.55 ± 0.05 | 0.20 ± 0.02 | 10.9 ± 1.04 | 3.68 ± 0.65 | 1.36 ± 0.06 | 69.3 ± 3.20 | 1.82 ± 0.03 | 1.16 ± 0.05 | 1.22 ± 0.04 | 1.94 | 70.0 | 1.40 | |
| C/N = 13 | 0.60 ± 0.07 | 0.76 ± 0.05 | 1.64 ± 0.08 | 12.6 ± 1.51 | 3.13 ± 0.10 | 0.93 ± 0.05 | 67.8 ± 3.23 | 1.40 ± 0.03 | 1.03 ± 0.06 | 0.19 ± 0.03 | 1.62 | 68.1 | 1.38 | |
| A. subrufescens | ||||||||||||||
| C/N = 10 | 0.43 ± 0.03 | 0.71 ± 0.04 | nd | 12.4 ± 1.00 | 2.91 ± 0.07 | 1.68 ± 0.06 | 63.3 ± 2.80 | 1.60 ± 0.03 | 3.40 ± 0.12 | 1.97 ± 0.05 | 3.90 | 64.3 | 1.28 | |
| C/N = 13 | 0.51 ± 0.02 | 0.80 ± 0.06 | nd | 13.9 ± 0.83 | 3.12 ± 0.06 | 1.41 ± 0.06 | 65.7 ± 3.12 | 1.20 ± 0.05 | 3.10 ± 0.08 | 1.65 ± 0.01 | 2.82 | 66.8 | 1.33 | |
| Substrate/Fungi | α-Tocopherol | α-Tocotrienol | β-Tocotrienol | δ-Tocopherol | |
|---|---|---|---|---|---|
| P. ostreatus | |||||
| WS * | 144 c | 0.78 ± 0.04 | 0.28 ± 0.04 | 0.27 ± 0.03 | nd ** |
| 144 p | 0.79 ± 0.03 | 0.26 ± 0.03 | 0.27 ± 0.04 | nd | |
| 144 s | 0.67 ± 0.03 | 0.30 ± 0.03 | 0.15 ± 0.02 | 0.42 ± 0.02 | |
| 150 c | 0.13 ± 0.03 | 0.71 ± 0.03 | 0.16 ± 0.03 | 1.19 ± 0.09 | |
| 150 p | 0.14 ± 0.03 | 0.73 ± 0.03 | 0.16 ± 0.03 | 1.20 ± 0.09 | |
| 150 s | 0.12 ± 0.04 | 0.65 ± 0.04 | 0.10 ± 0.02 | 0.99 ± 0.05 | |
| BOS | 144 c | nd | 1.21 ± 0.10 | 0.06 ± 0.01 | 1.98 ± 0.11 |
| 144 p | nd | 1.31 ± 0.10 | 0.07 ± 0.01 | 2.05 ± 0.10 | |
| 144 s | nd | 1.10 ± 0.09 | 0.06 ± 0.01 | 1.7 ± 0.14 | |
| 150 c | 1.02 ± 0.02 | 0.18 ± 0.02 | 0.34 ± 0.03 | nd | |
| 150 p | 1.12 ± 0.02 | 0.19 ± 0.02 | 0.34 ± 0.03 | nd | |
| 150 s | 1.08 ± 0.02 | 0.31 ± 0.02 | 0.35 ± 0.04 | nd | |
| RB | 144 c | 0.75 ± 0.02 | 0.15 ± 0.02 | 0.22 ± 0.03 | nd |
| 144 p | 0.76 ± 0.05 | 0.19 ± 0.05 | 0.24 ± 0.04 | nd | |
| 144 s | 0.7 ± 0.040 | 0.14 ± 0.04 | 0.20 ± 0.04 | nd | |
| 150 c | 0.62 ± 0.01 | 0.1 ± 0.010 | 0.17 ± 0.02 | nd | |
| 150 p | 0.67 ± 0.03 | 0.1 ± 0.030 | 0.20 ± 0.02 | nd | |
| 150 s | 0.67 ± 0.02 | 0.09 ± 0.02 | 0.15 ± 0.02 | nd | |
| BWS | 144 c | 1.14 ± 0.02 | 0.31 ± 0.02 | 0.17 ± 0.02 | 0.42 ± 0.02 |
| 144 p | 1.30 ± 0.04 | 0.35 ± 0.04 | 0.19 ± 0.04 | 0.40 ± 0.05 | |
| 144 s | 1.10 ± 0.06 | 0.30 ± 0.06 | 0.18 ± 0.03 | 0.40 ± 0.07 | |
| 150 c | 0.71 ± 0.02 | 0.15 ± 0.02 | 0.22 ± 0.03 | nd | |
| 150 p | 0.72 ± 0.03 | 0.16 ± 0.03 | 0.23 ± 0.02 | nd | |
| 150 s | 0.70 ± 0.03 | 0.15 ± 0.03 | 0.18 ± 0.03 | nd | |
| CR | 144 c | 0.82 ± 0.01 | 0.10 ± 0.01 | 0.28 ± 0.03 | 0.16 ± 0.02 |
| 144 p | 0.83 ± 0.01 | 0.10 ± 0.01 | 0.29 ± 0.04 | 0.17 ± 0.03 | |
| 144 s | 0.81 ± 0.01 | 0.10 ± 0.01 | 0.26 ± 0.03 | 0.15 ± 0.02 | |
| 150 c | 1.12 ± 0.01 | 0.19 ± 0.01 | 0.33 ± 0.03 | nd | |
| 150 p | 1.03 ± 0.02 | 0.16 ± 0.02 | 0.32 ± 0.03 | nd | |
| 150 s | 0.91 ± 0.02 | 0.11 ± 0.02 | 0.30 ± 0.03 | nd | |
| P. eryngii | |||||
| WS | 166 c | nd | 0.29 ± 0.02 | nd | 0.50 ± 0.03 |
| 166 p | nd | 0.31 ± 0.02 | 0.11 ± 0.02 | 0.54 ± 0.02 | |
| 166 s | nd | 0.30 ± 0.02 | nd | 0.54 ± 0.02 | |
| 173-6 c | 0.75 ± 0.04 | 0.19 ± 0.02 | 0.2 ± 0.02 | nd | |
| 173-6 p | 0.77 ± 0.04 | 0.21 ± 0.02 | 0.24 ± 0.02 | nd | |
| 173-6 s | 0.58 ± 0.03 | 0.19 ± 0.02 | 0.19 ± 0.02 | nd | |
| BOS | 166 c | 0.07 ± 0.01 | 0.55 ± 0.04 | nd | 1.40 ± 0.04 |
| 166 p | 0.07 ± 0.01 | 0.56 ± 0.04 | nd | 1.43 ± 0.04 | |
| 166 s | 0.06 ± 0.01 | 0.49 ± 0.03 | nd | 1.13 ± 0.04 | |
| 173-6 c | nd | 0.68 ± 0.04 | nd | 0.64 ± 0.03 | |
| 173-6 p | 0.69 ± 0.03 | 0.29 ± 0.03 | 0.13 ± 0.02 | 0.32 ± 0.02 | |
| 173-6 s | 0.68 ± 0.03 | 0.27 ± 0.03 | 0.11 ± 0.02 | 0.32 ± 0.02 | |
| RB | 166 c | 0.75 ± 0.04 | 0.10 ± 0.01 | 0.24 ± 0.02 | nd |
| 166 p | 0.79 ± 0.04 | 0.10 ± 0.01 | 0.27 ± 0.03 | nd | |
| 166 s | 0.72 ± 0.04 | 0.09 ± 0.01 | 0.23 ± 0.02 | nd | |
| 173-6 c | 0.70 ± 0.04 | 0.10 ± 0.01 | 0.20 ± 0.04 | nd | |
| 173-6 p | 0.71 ± 0.05 | 0.12 ± 0.01 | 0.25 ± 0.04 | nd | |
| 173-6 s | 0.66 ± 0.06 | 0.10 ± 0.01 | 0.20 ± 0.03 | nd | |
| BWS | 166 c | 1.12 ± 0.02 | 0.31 ± 0.03 | 0.16 ± 0.02 | 0.43 ± 0.03 |
| 166 p | 1.14 ± 0.03 | 0.35 ± 0.03 | 0.17 ± 0.02 | 0.44 ± 0.02 | |
| 166 s | 1.11 ± 0.03 | 0.30 ± 0.02 | 0.16 ± 0.02 | 0.40 ± 0.03 | |
| 173-6 c | 1.00 ± 0.10 | 0.15 ± 0.01 | 0.19 ± 0.03 | nd | |
| 173-6 p | 1.16 ± 0.09 | 0.17 ± 0.01 | 0.24 ± 0.03 | nd | |
| 173-6 s | 0.90 ± 0.09 | 0.12 ± 0.01 | 0.15 ± 0.02 | nd | |
| CR | 166 c | 0.81 ± 0.03 | 0.11 ± 0.01 | 0.27 ± 0.03 | 0.16 ± 0.03 |
| 166 p | 0.82 ± 0.02 | 0.12 ± 0.01 | 0.28 ± 0.03 | 0.17 ± 0.02 | |
| 166 s | 0.80 ± 0.03 | 0.10 ± 0.02 | 0.24 ± 0.03 | 0.12 ± 0.02 | |
| 173-6 c | 1.11 ± 0.06 | 0.10 ± 0.02 | 0.38 ± 0.03 | nd | |
| 173-6 p | 1.12 ± 0.06 | 0.12 ± 0.02 | 0.39 ± 0.05 | nd | |
| 173-6 s | 0.99 ± 0.02 | 0.09 ± 0.01 | 0.35 ± 0.05 | nd | |
| A. bisporus | |||||
| C/N = 10 | c | 1.17 ± 0.10 | 0.39 ± 0.05 | 0.38 ± 0.05 | nd |
| p | 2.78 ± 0.10 | 0.28 ± 0.05 | 0.50 ± 0.03 | nd | |
| s | 1.02 ± 0.08 | 0.21 ± 0.05 | 0.37 ± 0.02 | nd | |
| C/N = 13 | c | 0.84 ± 0.03 | 0.19 ± 0.02 | 0.34 ± 0.03 | nd |
| p | 0.93 ± 0.06 | 0.25 ± 0.02 | 0.36 ± 0.03 | nd | |
| s | 0.81 ± 0.06 | 0.14 ± 0.02 | 0.24 ± 0.03 | nd | |
| A. subrufescens | |||||
| C/N = 10 | c | 2.07 ± 0.12 | 0.46 ± 0.08 | 0.36 ± 0.06 | nd |
| p | 1.86 ± 0.09 | 0.39 ± 0.06 | 0.42 ± 0.07 | nd | |
| s | 0.95 ± 0.08 | 0.35 ± 0.06 | 0.35 ± 0.03 | nd | |
| C/N = 13 | c | 1.98 ± 0.07 | 0.47 ± 0.03 | 0.34 ± 0.01 | nd |
| p | 2.13 ± 0.06 | 0.33 ± 0.02 | 0.38 ± 0.02 | nd | |
| s | 1.08 ± 0.05 | 0.40 ± 0.04 | 0.20 ± 0.02 | nd | |
| Substrate/Fungi | Mushroom Part | Moisture (%) | Ash (%) | TPC (mg GAE/g) | DPPH∙ (mg trolox/g) | ABTS∙+ (mg trolox/g) | FRAP (mg trolox/g) |
|---|---|---|---|---|---|---|---|
| P. ostreatus | |||||||
| WS * | 144 c | 88.80 ± 3.49 | 4.89 ± 1.11 | 24.02 ± 2.56 | 2.48 ± 0.25 | 3.21 ± 0.16 | 3.26 ± 0.28 |
| 144 p | 88.95 ± 2.52 | 5.19 ± 0.90 | 25.87 ± 3.23 | 2.09 ± 0.45 | 2.97 ± 0.11 | 2.92 ± 0.53 | |
| 144 s | 82.87 ± 2.66 | 3.41 ± 0.82 | 11.29 ± 2.98 | 0.44 ± 0.09 | 2.17 ± 0.09 | 0.25 ± 0.02 | |
| 150 c | 88.65 ± 3.85 | 5.21 ± 0.92 | 24.01 ± 3.06 | 2.69 ± 0.12 | 3.87 ± 0.14 | 4.11 ± 0.23 | |
| 150 p | 88.52 ± 2.68 | 5.50 ± 0.73 | 23.24 ± 1.28 | 2.39 ± 0.32 | 2.23 ± 0.20 | 2.24 ± 0.17 | |
| 150 s | 83.21 ± 1.84 | 4.02 ± 0.90 | 10.41 ± 1.76 | 0.35 ± 0.03 | 1.04 ± 0.07 | 1.58 ± 0.31 | |
| BOS | 144 c | 90.05 ± 3.96 | 6.40 ± 1.25 | 24.01 ± 2.05 | 3.63 ± 0.20 | 5.57 ± 0.43 | 4.52 ± 0.49 |
| 144 p | 90.25 ± 3.54 | 8.50 ± 2.40 | 23.24 ± 3.12 | 4.09 ± 0.45 | 5.73 ± 0.28 | 5.42 ± 0.63 | |
| 144 s | 85.24 ± 3.45 | 5.74 ± 3.40 | 10.41 ± 1.62 | 3.49 ± 0.23 | 5.05 ± 0.39 | 4.34 ± 0.38 | |
| 150 c | 89.32 ± 2.89 | 6.55 ± 1.31 | 40.39 ± 3.87 | 4.13 ± 0.18 | 5.73 ± 0.82 | 5.44 ± 0.23 | |
| 150 p | 88.95 ± 2.85 | 6.26 ± 1.21 | 36.62 ± 2.92 | 3.98 ± 0.27 | 4.70 ± 0.43 | 5.61 ± 0.47 | |
| 150 s | 85.3 ± 2.35 | 5.23 ± 1.25 | 23.11 ± 1.58 | 0.36 ± 0.04 | 2.48 ± 0.13 | 1.41 ± 0.19 | |
| RB | 144 c | 90.4 ± 3.62 | 6.74 ± 0.92 | 38.42 ± 2.60 | 4.90 ± 0.25 | 6.73 ± 0.24 | 6.58 ± 0.67 |
| 144 p | 89.98 ± 5.27 | 6.05 ± 1.02 | 40.83 ± 3.47 | 5.31 ± 0.08 | 9.26 ± 0.56 | 7.28 ± 0.77 | |
| 144 s | 85.65 ± 4.54 | 6.24 ± 0.96 | 36.02 ± 2.89 | 4.71 ± 0.25 | 6.04 ± 0.33 | 6.03 ± 0.48 | |
| 150 c | 88.24 ± 4.45 | 5.91 ± 0.83 | 48.91 ± 4.31 | 8.35 ± 0.73 | 9.97 ± 0.27 | 9.38 ± 0.36 | |
| 150 p | 89.92 ± 5.24 | 5.54 ± 1.24 | 51.02 ± 2.78 | 8.72 ± 0.57 | 11.27 ± 0.31 | 11.06 ± 0.41 | |
| 150 s | 85.24 ± 5.60 | 6.25 ± 1.35 | 48.01 ± 3.90 | 7.71 ± 0.44 | 8.42 ± 0.78 | 8.43 ± 0.29 | |
| BWS | 144 c | 88.94 ± 3.66 | 5.56 ± 0.94 | 38.35 ± 4.12 | 3.61 ± 0.22 | 6.13 ± 0.28 | 4.51 ± 0.20 |
| 144 p | 90.23 ± 4.54 | 5.50 ± 1.24 | 39.26 ± 3.88 | 3.73 ± 0.63 | 7.41 ± 0.49 | 4.64 ± 0.46 | |
| 144 s | 85.35 ± 6.02 | 6.27 ± 1.05 | 36.01 ± 3.23 | 3.44 ± 0.29 | 5.54 ± 0.20 | 4.22 ± 0.33 | |
| 150 c | 89.24 ± 2.85 | 5.13 ± 0.80 | 36.13 ± 3.75 | 4.82 ± 0.55 | 5.82 ± 0.37 | 5.92 ± 0.47 | |
| 150 p | 89.24 ± 5.52 | 4.86 ± 1.05 | 38.72 ± 4.01 | 4.97 ± 0.14 | 6.02 ± 0.19 | 6.15 ± 0.24 | |
| 150 s | 85.25 ± 5.34 | 5.32 ± 1.25 | 34.82 ± 2.66 | 4.66 ± 0.16 | 5.54 ± 0.34 | 5.66 ± 0.29 | |
| CR | 144 c | 88.52 ± 2.54 | 5.89 ± 0.94 | 59.11 ± 4.32 | 8.16 ± 0.85 | 7.40 ± 0.65 | 7.54 ± 0.76 |
| 144 p | 88.65 ± 3.62 | 5.80 ± 0.99 | 70.67 ± 3.65 | 8.42 ± 0.67 | 7.74 ± 0.27 | 7.76 ± 0.53 | |
| 144 s | 85.60 ± 3.34 | 5.98 ± 0.89 | 41.40 ± 3.25 | 5.08 ± 0.43 | 4.59 ± 0.39 | 4.56 ± 0.48 | |
| 150c | 88.91 ± 3.70 | 6.54 ± 0.92 | 61.23 ± 4.82 | 7.78 ± 0.27 | 7.01 ± 0.44 | 6.47 ± 0.39 | |
| 150 p | 89.52 ± 4.35 | 6.45 ± 1.05 | 62.79 ± 4.07 | 7.94 ± 0.31 | 7.77 ± 0.26 | 6.98 ± 0.58 | |
| 150 s | 85.14 ± 3.64 | 6.68 ± 1.20 | 51.01 ± 3.89 | 2.59 ± 0.19 | 3.83 ± 0.10 | 3.41 ± 0.32 | |
| P. eryngii | |||||||
| WS | 166 c | 89.63 ± 2.45 | 6.29 ± 0.90 | 31.45 ± 3.35 | 3.16 ± 0.24 | 5.91 ± 0.28 | 5.01 ± 0.29 |
| 166 p | 88.97 ± 3.51 | 6.51 ± 0.83 | 28.03 ± 2.08 | 2.93 ± 0.39 | 4.47 ± 0.32 | 4.60 ± 0.38 | |
| 166 s | 83.65 ± 2.32 | 5.97 ± 0.98 | 25.50 ± 2.65 | 1.90 ± 0.06 | 3.43 ± 0.17 | 3.27 ± 0.22 | |
| 173-6 c | 88.58 ± 2.45 | 5.73 ± 0.85 | 28.82 ± 1.18 | 3.61 ± 0.09 | 3.03 ± 0.11 | 3.23 ± 0.47 | |
| 173-6 p | 88.71 ± 2.64 | 5.87 ± 0.92 | 22.50 ± 3.02 | 3.34 ± 0.28 | 3.79 ± 0.27 | 3.15 ± 0.33 | |
| 173-6 s | 84.21 ± 3.52 | 3.57 ± 0.64 | 20.47 ± 2.80 | 2.25 ± 0.16 | 2.57 ± 0.14 | 1.27 ± 0.12 | |
| BOS | 166 c | 88.34 ± 2.41 | 6.05 ± 0.62 | 33.66 ± 2.04 | 4.23 ± 0.36 | 6.01 ± 0.25 | 5.36 ± 0.29 |
| 166 p | 88.57 ± 2.96 | 6.51 ± 0.85 | 24.19 ± 2.24 | 3.83 ± 0.23 | 5.98 ± 0.49 | 4.39 ± 0.22 | |
| 166 s | 81.36 ± 3.09 | 4.85 ± 0.90 | 19.20 ± 1.99 | 1.27 ± 0.05 | 2.34 ± 0.21 | 2.47 ± 0.10 | |
| 173-6 c | 87.64 ± 2.49 | 6.45 ± 0.86 | 28.56 ± 3.54 | 3.22 ± 0.28 | 4.26 ± 0.47 | 3.99 ± 0.43 | |
| 173-6 p | 87.20 ± 2.54 | 6.58 ± 0.84 | 22.69 ± 2.87 | 3.07 ± 0.15 | 3.01 ± 0.24 | 4.99 ± 0.28 | |
| 173-6 s | 81.41 ± 2.65 | 4.85 ± 0.99 | 13.57 ± 1.21 | 2.59 ± 0.27 | 2.06 ± 0.19 | 2.42 ± 0.15 | |
| RB | 166 c | 89.35 ± 3.01 | 4.65 ± 0.65 | 23.92 ± 2.23 | 12.12 ± 0.76 | 14.84 ± 0.39 | 11.12 ± 0.21 |
| 166 p | 88.23 ± 2.65 | 6.55 ± 1.01 | 25.21 ± 1.53 | 13.22 ± 0.59 | 15.45 ± 0.67 | 12.26 ± 0.38 | |
| 166 s | 80.10 ± 2.15 | 5.35 ± 0.57 | 19.21 ± 2.76 | 9.61 ± 0.42 | 11.95 ± 0.26 | 7.23 ± 0.09 | |
| 173-6 c | 87.26 ± 2.34 | 5.34 ± 0.66 | 27.68 ± 3.54 | 10.75 ± 0.34 | 11.55 ± 0.23 | 10.83 ± 0.25 | |
| 173-6 p | 88.64 ± 1.30 | 5.44 ± 0.71 | 28.09 ± 3.07 | 11.61 ± 0.53 | 13.38 ± 0.69 | 11.89 ± 0.43 | |
| 173-6 s | 82.35 ± 1.65 | 6.50 ± 0.60 | 26.13 ± 2.91 | 9.11 ± 0.49 | 10.24 ± 0.72 | 11.89 ± 0.61 | |
| BWS | 166 c | 87.69 ± 2.00 | 5.71 ± 1.20 | 27.02 ± 2.69 | 11.11 ± 0.94 | 13.83 ± 0.31 | 11.42 ± 0.73 |
| 166 p | 88.25 ± 2.30 | 5.55 ± 1.32 | 27.30 ± 3.21 | 10.76 ± 0.57 | 14.28 ± 0.96 | 12.13 ± 0.56 | |
| 166 s | 84.36 ± 1.91 | 5.31 ± 1.30 | 25.72 ± 2.45 | 9.93 ± 0.42 | 13.74 ± 0.64 | 9.38 ± 0.40 | |
| 173-6 c | 85.74 ± 1.85 | 5.47 ± 0.95 | 27.68 ± 1.03 | 11.29 ± 0.32 | 14.72 ± 0.78 | 11.84 ± 0.32 | |
| 173-6 p | 88.20 ± 1.54 | 5.23 ± 0.64 | 28.09 ± 1.82 | 11.40 ± 0.18 | 15.02 ± 0.92 | 13.41 ± 0.55 | |
| 173-6 s | 80.25 ± 1.68 | 6.14 ± 0.34 | 26.13 ± 1.63 | 7.23 ± 0.53 | 14.08 ± 0.53 | 9.44 ± 0.65 | |
| CR | 166 c | 90.21 ± 2.14 | 5.34 ± 0.57 | 53.70 ± 2.95 | 9.52 ± 0.29 | 9.87 ± 0.57 | 9.04 ± 0.33 |
| 166 p | 89.25 ± 1.68 | 5.41 ± 0.55 | 56.47 ± 2.50 | 11.12 ± 0.76 | 10.54 ± 0.29 | 9.72 ± 0.49 | |
| 166 s | 87.10 ± 1.87 | 4.87 ± 0.48 | 48.27 ± 3.32 | 8.84 ± 0.54 | 8.78 ± 0.72 | 8.66 ± 0.51 | |
| 173-6 c | 88.64 ± 1.74 | 4.68 ± 0.34 | 42.57 ± 2.09 | 10.58 ± 0.23 | 7.06 ± 0.69 | 6.04 ± 0.46 | |
| 173-6 p | 89.01 ± 1.67 | 4.88 ± 0.60 | 45.50 ± 2.78 | 11.26 ± 0.88 | 7.76 ± 0.42 | 7.53 ± 0.53 | |
| 173-6 s | 80.24 ± 1.99 | 5.47 ± 0.47 | 41.48 ± 3.01 | 7.44 ± 0.39 | 6.92 ± 0.37 | 5.66 ± 0.31 | |
| Mineral Content (w/w, d. w.) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Substrate/Fungi | K (g/Kg) | Na (g/kg) | Mg (g/kg) | P (g/kg) | Ca (g/kg) | Fe (ppm) | Mn (ppm) | Cu (ppm) | Zn (ppm) | B (ppm) | |
| P. ostreatus | |||||||||||
| WS * | 144 c | 250 | 8.0 | 18 | 82 | 19 | 68 | 6.3 | 12 | 66 | 1.0 |
| 144 p | 280 | 9.0 | 21 | 84 | 10 | 72 | 6.8 | 15 | 69 | 1.2 | |
| 144 s | 210 | 7.0 | 14 | 64 | 26 | 31 | 3.8 | 11 | 50 | nd ** | |
| 150 c | 270 | 5.0 | 17 | 77 | 20 | 71 | 8.5 | 11 | 67 | 0.3 | |
| 150 p | 290 | 4.0 | 19 | 80 | 10 | 70 | 9.0 | 12 | 70 | 1.2 | |
| 150 s | 200 | 7.5 | 13 | 50 | 28 | 24 | 8.6 | 7.6 | 41 | 1.8 | |
| BOS | 144 c | 290 | 11.0 | 21 | 92 | 17 | 99 | 10 | 17 | 72 | 0.3 |
| 144 p | 330 | 13.0 | 23 | 95 | 9 | 110 | 16 | 18 | 78 | 1.2 | |
| 144 s | 250 | 8.5 | 17 | 85 | 22 | 58 | 7.8 | 14 | 50 | 0.4 | |
| 150 c | 320 | 7.0 | 18 | 93 | 28 | 94 | 13 | 15 | 68 | 4.6 | |
| 150 p | 350 | 8.5 | 25 | 95 | 10 | 97 | 15 | 17 | 75 | 6.2 | |
| 150 s | 300 | 8.0 | 14 | 83 | 32 | 48 | 6.8 | 11 | 53 | 3.4 | |
| RB | 144 c | 290 | 2.0 | 14 | 88 | 18 | 82 | 10 | 20 | 58 | 15.6 |
| 144 p | 340 | 3.5 | 19 | 92 | 6 | 92 | 15 | 14 | 76 | 17.2 | |
| 144 s | 250 | 1.5 | 10 | 75 | 22 | 65 | 9 | 12 | 45 | 11.3 | |
| 150 c | 260 | 3.0 | 13 | 87 | 16 | 72 | 14 | 22 | 62 | 7.6 | |
| 150 p | 350 | 3.6 | 18 | 90 | 10 | 86 | 18 | 14 | 70 | 10.6 | |
| 150 s | 220 | 1.0 | 9 | 80 | 19 | 62 | 12 | 8 | 45 | 6.8 | |
| BWS | 144 c | 250 | 8.0 | 16 | 99 | 23 | 83 | 9.0 | 12 | 68 | 1.6 |
| 144 p | 300 | 10.0 | 20 | 100 | 12 | 90 | 12 | 15 | 90 | 1.8 | |
| 144 s | 170 | 5.0 | 9 | 65 | 28 | 58 | 7.0 | 9 | 45 | 0.9 | |
| 150 c | 270 | 1.0 | 20 | 78 | 20 | 91 | 11 | 11 | 72 | 2.0 | |
| 150 p | 300 | 1.5 | 25 | 80 | 10 | 100 | 18 | 15 | 95 | 2.5 | |
| 150 s | 120 | 0.8 | 17 | 70 | 26 | 80 | 9 | 10 | 50 | 1.8 | |
| CR | 144 c | 200 | 1.0 | 12 | 87 | 18 | 67 | 4.0 | 12 | 65 | 1.8 |
| 144 p | 230 | 1.2 | 12 | 98 | 8 | 98 | 4.2 | 11 | 71 | 1.5 | |
| 144 s | 130 | 1.2 | 11 | 65 | 21 | 54 | 2.8 | 5.6 | 37 | 3.2 | |
| 150 c | 220 | 0.9 | 11 | 80 | 16 | 70 | 8.3 | 11 | 68 | 2.1 | |
| 150 p | 240 | 1.1 | 15 | 86 | 9 | 91 | 9.4 | 12.6 | 72 | 2.7 | |
| 150 s | 150 | 8.8 | 9 | 60 | 20 | 55 | 5.6 | 8.7 | 48 | 1.1 | |
| P. eryngii | |||||||||||
| WS | 166 c | 350 | 5.0 | 12.5 | 85 | 17 | 65 | 5.4 | 6.0 | 26 | 0.8 |
| 166 p | 370 | 7.0 | 15 | 89 | 8 | 90 | 5.9 | 9.3 | 29 | 1.1 | |
| 166 s | 330 | 4.0 | 9 | 44 | 23 | 32 | 2.9 | 5.6 | 19 | 0.6 | |
| 173-6 c | 330 | 5.0 | 10 | 86 | 16.50 | 56 | 4.7 | 6.1 | 23 | 0.9 | |
| 173-6 p | 350 | 4.5 | 15 | 99 | 9 | 97 | 5.2 | 10 | 25 | 1.2 | |
| 173-6 s | 210 | 5.0 | 11 | 45 | 20 | 25 | 3.2 | 6.6 | 18 | 0.6 | |
| BOS | 166 c | 340 | 4.0 | 14 | 86 | 16 | 50 | 5.7 | 10 | 24 | 1.1 |
| 166 p | 380 | 6.0 | 17 | 95 | 9 | 65 | 6.9 | 15 | 29 | 2.2 | |
| 166 s | 340 | 5.0 | 9 | 50 | 20 | 33 | 4.1 | 6.7 | 25 | 2.7 | |
| 173-6 c | 320 | 11.0 | 15 | 88 | 18 | 60 | 7.2 | 6.6 | 32 | nd | |
| 173-6 p | 350 | 12.0 | 18 | 95 | 8 | 70 | 7.8 | 7.9 | 40 | nd | |
| 173-6 | 290 | 7.0 | 7 | 70 | 22 | 35 | 6.2 | 4.5 | 25 | nd | |
| RB | 166 c | 340 | 1.2 | 20 | 91 | 18 | 31 | 8.2 | 20 | 55 | 1.6 |
| 166 p | 370 | 2.5 | 22 | 99 | 6 | 37 | 8.9 | 42 | 80 | 2.4 | |
| 166 s | 300 | 0.9 | 10 | 60 | 22 | 20 | 7.6 | 16 | 30 | 2.0 | |
| 166 c | 310 | 1.4 | 14 | 97 | 16 | 41 | 7.3 | 11 | 64 | 4.7 | |
| 166 p | 330 | 2.5 | 18 | 100 | 10 | 48 | 8.9 | 15 | 72 | 7.9 | |
| 166 s | 260 | 1.0 | 12.5 | 78 | 19 | 36 | 6.5 | 9.6 | 50 | 4.0 | |
| BWS | 166 c | 320 | 1.5 | 19 | 90 | 19 | 60 | 9.0 | 50 | 64 | 1.5 |
| 166 p | 350 | 2.5 | 22 | 100 | 9 | 70 | 9.8 | 62 | 90 | 2.0 | |
| 166 s | 270 | 0.9 | 10 | 50 | 22 | 42 | 6.8 | 28 | 34 | 0.9 | |
| 173-6 c | 340 | 1.6 | 15 | 75 | 17 | 55 | 8.6 | 38 | 70 | 5.2 | |
| 173-6 p | 370 | 2.0 | 17 | 90 | 9 | 69 | 9.8 | 45 | 86 | 7.2 | |
| 173-6 s | 270 | 0.8 | 10 | 65 | 20 | 38 | 5.6 | 15 | 37 | 3.4 | |
| CR | 166 c | 350 | 2.3 | 19 | 87 | 13 | 50 | 1.6 | 14 | 78 | 2.4 |
| 166 p | 370 | 3.0 | 22 | 95 | 6 | 65 | 2.7 | 17 | 88 | 2.8 | |
| 166 s | 290 | 2.0 | 15 | 55 | 18 | 40 | 1.2 | 10 | 51 | 2.0 | |
| 173-6 c | 310 | 0.9 | 14 | 90 | 17 | 61 | 6.2 | 8.3 | 62 | 4.4 | |
| 173-6 p | 340 | 3.7 | 21 | 95 | 7 | 58 | 8.2 | 16 | 87 | <0.1 | |
| 173-6 | 210 | 1.5 | 10 | 65 | 20 | 55 | 5.4 | 8.0 | 58 | 2.9 | |
| Agaricus spp. | |||||||||||
| A. bisporus (C/N = 10) | c | 410 | 6.8 | 21 | 98 | nd | 46 | 2.5 | 44 | 68 | 3.2 |
| p | 450 | 7.8 | 23 | 100 | 2 | 57 | 4.0 | 56 | 75 | 3.6 | |
| s | 320 | 5.8 | 17.5 | 56 | nd | 38 | 1.2 | 30 | 60 | 2.8 | |
| A. bisporus (C/N = 13) | c | 390 | 7.3 | 19 | 89 | 15 | 44 | 1.1 | 33 | 71 | 1.6 |
| p | 430 | 8.0 | 20 | 94 | 3 | 50 | 2.5 | 40 | 76 | 2.0 | |
| s | 370 | 5.2 | 15.4 | 43 | 10 | 32 | n.d | 25 | 60 | 1.2 | |
| A. subrufescens (C/N = 10) | c | 360 | 10.5 | 13 | 87 | 14 | 62 | 2.5 | 40 | 90 | 1.1 |
| p | 400 | 17 | 17 | 98 | 2 | 53 | 3.8 | 60 | 150 | 1.5 | |
| s | 270 | 5.7 | 8.4 | 62 | 5 | 31 | 2.0 | 20 | 60 | nd | |
| A. subrufescens (C/N = 13) | c | 380 | 8.3 | 15 | 92 | 13 | 67 | 2.9 | 46 | 97 | 1.3 |
| p | 400 | 9.5 | 19 | 103 | 5 | 50 | 3.7 | 58 | 145 | 1.7 | |
| s | 270 | 6.2 | 9.20 | 75 | 10 | 35 | 1.8 | 22 | 68 | 1.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diamantopoulou, P.; Fourtaka, K.; Melanouri, E.M.; Dedousi, M.; Diamantis, I.; Gardeli, C.; Papanikolaou, S. Examining the Impact of Substrate Composition on the Biochemical Properties and Antioxidant Activity of Pleurotus and Agaricus Mushrooms. Fermentation 2023, 9, 689. https://doi.org/10.3390/fermentation9070689
Diamantopoulou P, Fourtaka K, Melanouri EM, Dedousi M, Diamantis I, Gardeli C, Papanikolaou S. Examining the Impact of Substrate Composition on the Biochemical Properties and Antioxidant Activity of Pleurotus and Agaricus Mushrooms. Fermentation. 2023; 9(7):689. https://doi.org/10.3390/fermentation9070689
Chicago/Turabian StyleDiamantopoulou, Panagiota, Katerina Fourtaka, Eirini Maria Melanouri, Marianna Dedousi, Ilias Diamantis, Chrysavgi Gardeli, and Seraphim Papanikolaou. 2023. "Examining the Impact of Substrate Composition on the Biochemical Properties and Antioxidant Activity of Pleurotus and Agaricus Mushrooms" Fermentation 9, no. 7: 689. https://doi.org/10.3390/fermentation9070689
APA StyleDiamantopoulou, P., Fourtaka, K., Melanouri, E. M., Dedousi, M., Diamantis, I., Gardeli, C., & Papanikolaou, S. (2023). Examining the Impact of Substrate Composition on the Biochemical Properties and Antioxidant Activity of Pleurotus and Agaricus Mushrooms. Fermentation, 9(7), 689. https://doi.org/10.3390/fermentation9070689