1. Introduction
Organic acids (OA) are a large group of organic compounds that contain acidic properties and can be produced by microbial, plant, and animal organisms, while pure botanicals (PB) are secondary metabolites produced from plant metabolism. These compounds, such as OA and PB, have been studied in livestock production and were reported to have antimicrobial [
1] and immunomodulatory [
2] activities.
The use of the blend of OA and PB has been shown to consistently reduce the counts of
Campylobacter jejuni, a leading cause of foodborne illness in which the primary route of infection is undercooked poultry meat, from the caecal samples of poultry [
1]. In addition, Grilli et al. [
3] have demonstrated that a blend of OA and PB reduced the relative expression of interleukin 6 and 12, as well as transforming growth factor beta (TGF-β) in the ileum of pigs, which are inflammation markers. However, these biocompounds may be affected by the action of gastric juice in monogastric animals, which would reduce the efficacity of them; therefore, a microencapsulation process was developed [
4].
Microencapsulated blends of organic acids and pure botanicals (mOAPBs) have been used in the diet of monogastric animals to improve gut health and production [
1,
5], and, more recently, have been tested in calf and lactating cattle [
6,
7]. Dietary supplementation of mOAPBs in livestock animals has been shown to improve intestinal health by increasing villi length [
8] and promote maturation of the intestinal mucosa [
3]. Huyben et al. [
8] demonstrated that supplementation of mOAPBs to rainbow trout (
Oncorhynchus mykiss) can promote a reduction in the relative abundance of opportunistic pathogens in the gastrointestinal tract, such as
Aeromonas hydrophila and
Acinetobacter spp.
Additionally, when evaluated in broilers, Feye et al. [
9] reported that mOAPBs had organ-specific activity, which means that mOAPBs were effective in promoting a healthier microbiome in the jejunum; however, it was not effective in the ileum. Authors reported an increase in microorganisms of the family
Lactobacilliaceae and a reduction in microorganisms of the family
Staphylococcaceae in the jejunum of broilers fed mOAPBs when compared to the jejunum of animals fed a control diet without mOAPBs supplementation.
In ruminants, one of the first studies evaluating this specific blend of mOAPBs, which contains citric acid, sorbic acid, thymol, and vanillin, as a dietary supplement had the objective of evaluating the effects of the mOAPBs on heat-stressed lactating cows [
6]. Authors reported that dietary supplementation of mOAPBs partially restored the feed intake of heat-stressed animals and improved the energy-corrected milk (27.4 vs. 30.2 kg/d), yield of protein (0.69 vs. 0.80 kg/d), and lactose (1.08 vs. 1.26 kg/d) when compared to heat-stress animals with no mOAPBs supplementation. Authors also evaluated the intestinal permeability, which was not statistically significant (
p = 0.18); however, there was an interesting pattern that should be considered in future research, in which mOAPBs may reduce the intestinal permeability of heat-stressed cows.
Although most of the studies which have evaluated mOAPBs supplementation have focused on intestinal activity, we hypothesized that microencapsulation may not completely prevent partial ruminal degradation. Therefore, a fraction of its ingredients may be released in the rumen, thus causing changes in the microbiome and affecting the microbiome microenvironments (solid- and liquid-associated). Thus, the main objective of this study is to evaluate the effects of dietary supplementation of mOAPBs on ruminal microbiome in a dual-flow continuous culture system.
2. Materials and Methods
The University of Florida’s Institutional Animal Care and Use Committee approved all procedures using animals.
2.1. Experimental Design and Diet
An experiment using 8 dual-flow continuous culture fermenters was designed. The detailed description of ruminal fluid donors, feed preparation, diet composition, and experimental design was described in our companion paper [
10]. Briefly, the experiment was carried out in a duplicated 4 × 4 Latin square design with 4 treatments and 4 fermentation periods, using 8 dual-flow continuous culture fermenters. The basal diet was formulated to meet the requirements of a 680 kg high-producing Holstein dairy cow with an estimated intake of 25 kg of DM. The basal diet was composed of 47.2% corn silage, 14.5% grass hay, 15.5% ground corn, 20.5% soybean meal, 0.19% calcium phosphate, 0.14% magnesium oxide, 1.43% calcium carbonate, and 0.50% trace mineral premix. The nutritional composition of the basal diet was 16.1% crude protein, 2.43% ether extract, 30.9% neutral detergent fiber, 17.8% acid detergent fiber, and 32% starch.
The components of the mOAPBs tested (AviPlus® P, Vetagro S.p.A., Reggio Emilia, Italy) were hydrogenated and refined palm oil (55.6%), citric acid (25.0%), sorbic acid (16.7%), thymol (1.7%), and vanillin (1.0%). Four levels of supplement inclusion were used: 0, 0.12, 0.24, or 0.36% DM (which correspond to 0, 30, 60, and 90 g per 25 kg of TMR). All experimental diets were formulated to have comparable nutritional compositions, with 16.1% CP, 30.9% NDF, and 32.0% starch. Treatments were randomly assigned within Latin square for each period.
Two ruminally cannulated lactating Holstein dairy cows were used as ruminal content donors.
2.2. Dual-Flow Continuous Culture System
The experiment was carried out in a dual-flow continuous culture system, briefly, ruminal content from two ruminally cannulated lactating Holstein dairy cows were collected as inoculum. Cows averaged 229 days in milk, a 730 kg body weight, 26.7 kg milk yield, and were ruminally cannulated and kept at the Dairy Research Unit of the University of Florida in a free-stall barn with the other cows of the herd receiving a total mixed ratio (TMR) containing 50% corn silage. On the first day of each experimental period, the ruminal content was collected from the cows 2 h after the morning feeding, filtered through 4 layers of cheesecloth and stored in a prewarmed thermos. The inoculum was transported within 30 min of the collection to the laboratory and incubated in the prewarmed dual-flow in vitro system.
Ruminal fluid from both cows were pooled in a ratio of 1:1 (v/v) from each cow and distributed into the 8 pre-warmed fermenter vessels. Each fermenter was fed twice daily (730 h and 1730 h) at a constant rate of 106 g DM d−1 (53 g each meal). Each experimental period was comprised of 10 d of fermentation, where the first 7 d was used as an adaptation of the microbiome to the diet and system, and the last 3 d were used for sampling.
Separated effluent containers, for solid and liquid samples, were kept in a cold-water bath (4 °C) to prevent further fermentation and avoid DNA degradation. Solid effluent was filtered through 4 layers of cheesecloth and the solid material retained was collected to access the solid-attached microbiome microenvironment (SAM). The sample within the liquid effluent container was used to access the liquid-attached microbiome microenvironment (LAM). At 3, 6, and 9 h after morning feeding on days 8, 9, and 10, a 5 mL (volume) of the solid and liquid sample was collected and pooled across timepoints and days from each fermenter and frozen at −80 °C. Frozen samples were stored in a cooler with dry ice and shipped to Fera Diagnostics and Biologicals, Corp. (College Station, TX, USA) for DNA extraction and sequencing of the V4 region of 16S rRNA of bacteria using the primer 515F [
11] and single-end sequencing in a MiSeq Illumina Platform with 300 cycles. All raw sequences were deposited in the Sequence Read Archive of the National Center for Biotechnology Information (
https://www.ncbi.nlm.nih.gov/sra, deposited on 10 March 2023) under access no. PRJNA943156.
2.3. Data Processing and Microbiome Analysis
Processing and statistical analyses of the microbiome data were carried out in R (RStudio 3.0.1) using the pipeline described by Lobo et al. [
12]. Briefly, sequenced amplicons were processed using the DADA2 package of R and taxonomy was assigned using the 16S rRNA SILVA v. 138 database [
13]. Single-end reads were demultiplexed and the quality profiles of reads were inspected, filtered, and trimmed based on the quality scores. Reads were trimmed at position 30 and 280 base pair and the maximum number of expected errors (“maxEE”) was set to 0.5 (quality plot before and after cleaning can be visualized in the
Supplementary Figure S1a,b). A further quality check was carried out to remove chimeras. The taxonomy was assigned using the SILVA database as earlier mentioned and the taxonomy classification algorithm used was
IdTaxa.
Additional filtration to remove ASVs classified at the taxonomic level order as “Chloroplast” or family as “Mitochondria” was carried out. The
Microbiome package was used to merge the reads at the phylum level, and the Bacteroidota:Firmicutes ratio was calculated. The data was rarefied for the calculation of diversity indices. The calculation of alpha diversity (Observed, Chao1, Fisher, Shannon, and Inverse Simpson), was carried out using the
phyloseq and
microbiome packages of R. Mean coverage of sample microbiome after rarefaction was 0.99 for all samples (
Table S1).
2.4. Statistical Analysis
The experiment followed a duplicated 4 × 4 Latin Square design with four treatments and four periods, the data of Bacteroidota:Firmicutes ratio and alpha diversity from SAM and LAM was analyzed and combined. The normality of residuals and homogeneity of variance were examined for each continuous dependent variable using the Shapiro–Wilk test from the UNIVARIATE procedure of SAS 9.4 (SAS Institute Inc., Cary, NC, USA). Statistical analyses for the Bacteroidota:Firmicutes ratio and alpha diversity indices were performed using the MIXED procedure of SAS 9.4, using the model:
where
Yijkl is the observation
ijkl,
µ is the overall mean,
Ti is the fixed effect of treatment (
i = 1 to 4),
Fj is the fixed effect of fraction (
j = 1 to 2),
Ti ×
Fj is the interaction effect of treatment (
i = 1 to 4) and fraction (
j = 1 to 2),
Pk is the random effect of period (
k = 1 to 4),
Sl is the random effect of square (
l = 1 to 2),
S(
F)
kl is the random effect of fermenter (
k = 1 to 8) nested within the Latin square (
l = 1 to 2), and
eijkl is the random residual. The effect of the levels of inclusion of mOAPBs was depicted by polynomial contrasts (linear and quadratic) as well as by contrasting diets with and without supplementation (control vs. mOAPBs at 30, 60, and 90 g/d of supplementation). Significance was declared at
p ≤ 0.05, and tendency was declared at 0.05 <
p ≤ 0.10.
On the taxonomic data, the Prevalence Interval for Microbiome Evaluation (PIME) denoising algorithm was also applied to the
phyloseq object [
14]. The dissimilarity analysis was carried out using filtered data from PIME and data was merged at the taxonomic genus level. Function ordinate from
phyloseq, Bray–Curtis, and Euclidian distances were used. To test the treatment effect, PERMANOVA analysis from the
vegan package was carried out using 999 permutations and adonis2 function for analyzing and partitioning sums of squares using dissimilarities. The model to test the effects of the microbiome beta diversity between microbiome microenvironment (PAB vs. LAB) and mOAPBs treatment (control vs. mOAPB) were considered as fixed effects and had period, latin square, and fermenter as random effects. To evaluate the effect of mOAPBs supplementation within each microenvironment, SAM or LAM, the phyloseq object was split by microenvironment (SAM and LAM), then PERMANOVA was carried out individually for each sub dataset using mOAPB treatment (control vs. mOAPB) as the fixed effect and period, latin square, and fermenter as random effects.
The differential relative abundance at genus level on PIME filtered data was analyzed using the ALDEx2 package. ALDEx2 is a package that can analyze the differential abundance between two groups of the studied cohort, and for this analysis, we contrasted microenvironment (SAM vs. LAM). Within each microenvironment the effect of mOAPBs supplementation was evaluated (control vs. mOAPB). The Wilcoxon rank test was used to test the hypothesis and significance was declared when the p-value adjusted for Benjamini–Hochberg correction was ≤0.05 and the median effect size was ≥1.
3. Results
The results of alpha diversity indices and Bacteroidota:Firmicutes ratio are presented in
Table 1. There was no interaction effect between treatment and microenvironment (
p > 0.74), as well as no main effect of treatment on the analyzed variables (
p > 0.50). The solid-attached microbiome microenvironment had a lower Bacteroidota:Firmicutes ratio (
p < 0.001) and tended (
p = 0.09) to have a greater Inverse Simpson Index when compared to LAM. When evaluated using the orthogonal contrasts, there was no linear (
p > 0.88), quadratic (
p > 0.15), or mOAPB (
p > 0.46) effect. The results of beta-diversity are presented in
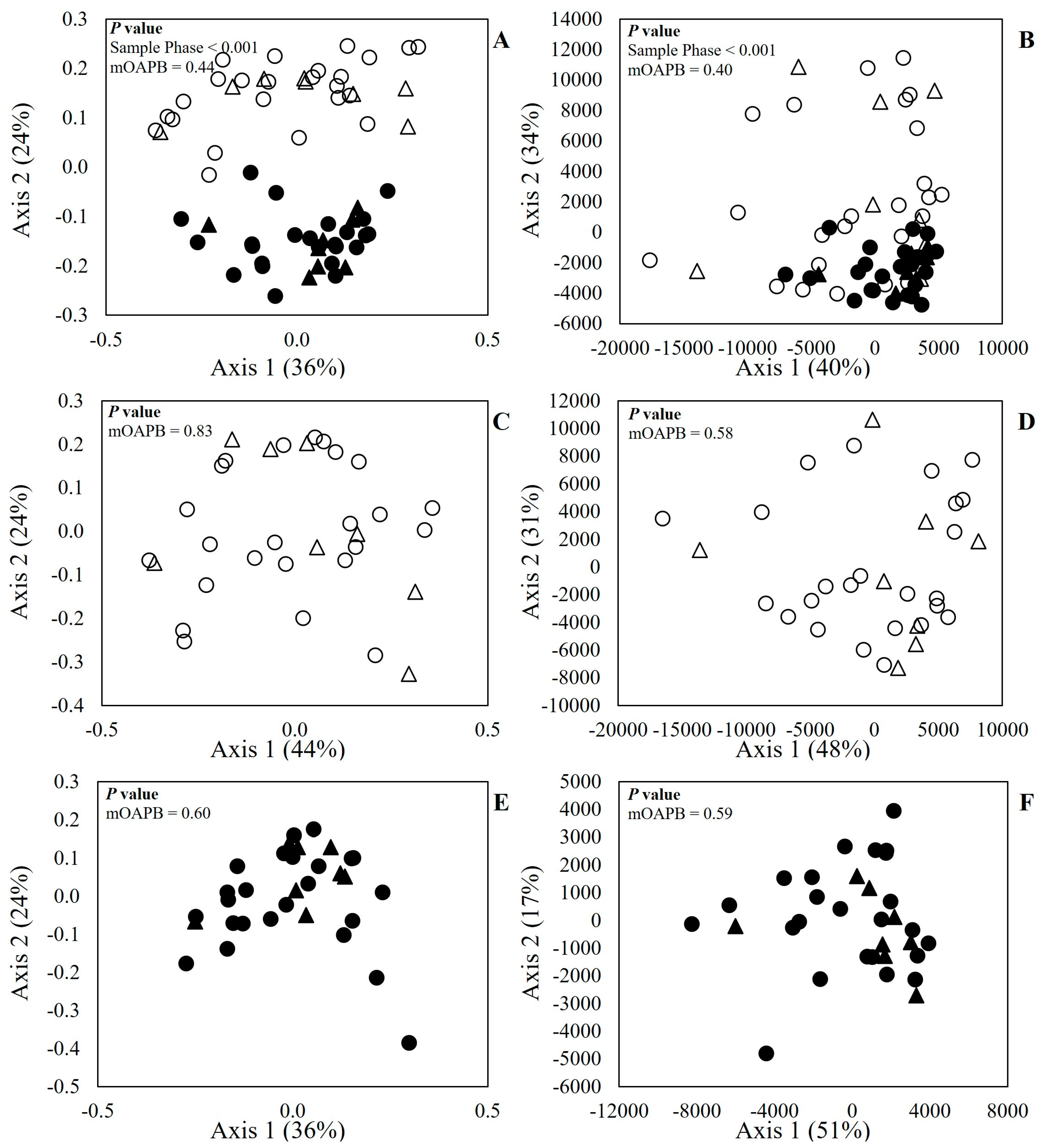
Figure 1. There was a visual cluster formation, where the microbiome profiles of LAM samples were similar to each other, and it was less comparable to SAM (
Figure 1A,B).
Dissimilarity was also confirmed by the PERMANOVA test, where the
p value was smaller than 0.001 for both Bray–Curtis and Euclidean distances. When the effect of treatment (
Figure 1C–F) was evaluated within SAM or LAM, there was no effect (
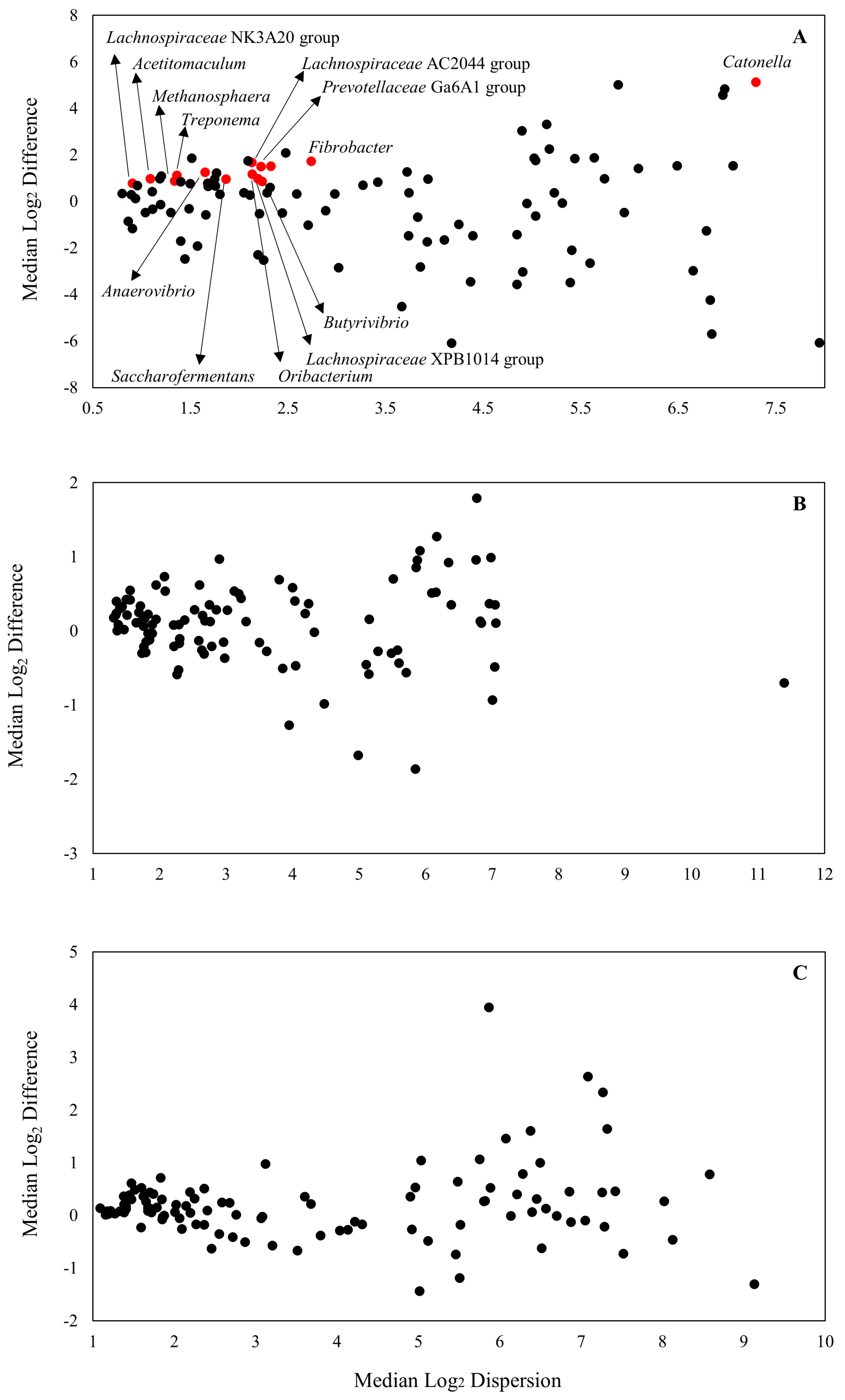
p > 0.58) of mOAPBs on the microbiome profile of the in vitro ruminal microenvironment. The MA plot is presented in
Figure 2. Out of the 101 ASVs identified at genus level in the analysis, there were 14 that had a greater relative abundance in SAM when compared to LAM samples (
Figure 2A and
Supplementary Table S2).
There were eight ASVs with greater abundance in SAM, which were classified as
Lachnospiraceae from the phylum Firmicutes. These ASVs corresponded to microorganisms related to the genus
Butyrivibrio,
Lachnospiraceae,
Oribacterium,
Catonella, and
Acetitomaculum. There were also other ASVs with a greater relative abundance in SAM classified as Firmicutes, such as
Saccharofermentans and
Anaerovibrio, from the
Hungateiclostridiaceae and
Selenomonadaceae Family. Some ASVs identified as genus
Prevotellaceae,
Methanosphera,
Fibrobacter, and
Treponema also had a greater relative abundance in SAM samples. There was no effect of mOAPBs on the differential abundance of microorganisms in the in vitro ruminal environment (
Figure 2B,C).
4. Discussion
The goal of this study was to evaluate the effects of levels of dietary mOAPBs supplementation on the solid- and liquid-associated microbiome microenvironment using an in vitro dual-flow continuous culture system. Results have demonstrated that mOAPBs supplementation did not affect the ruminal microbiome even when used in a dose 2 times greater than the recommended dose. These results suggest that microencapsulation might be effective in protecting biocompounds against ruminal fermentation and would increase the likelihood of their release in the intestine, which is the main site of activity of these compounds. In addition, there was a difference between the microbiome of LAM and SAM, which was expected, due to the fact that the profile of substrates to support microbial growth on both microenvironments are different from each other.
Solid- and liquid-associated microbiome microenvironment were composited mainly by Firmicutes, Bacteroidota, followed by Proteobacteria, which accounted for 94.9% and 84.8% of the identified reads for SAM and LAM, respectively. These values corroborate with previous research that has reported in vivo [
15] and in vitro [
16] ruminal microbiome composition. Several studies evaluating mice gut microbiome have reported that microorganisms of the Firmicutes phylum are more efficient in extracting energy from substrates than Bacteroidota, thus a reduction in the Firmicutes-to-Bacteroidota ratio could be beneficial to improving substrate utilization [
17]. Jami et al. [
15] evaluated the correlation of Firmicute-to-Bacteroidota ratio in dairy cattle using physiological and production parameters. The authors reported that the ratio was highly correlated (Pearson r = 0.72) with daily milk fat yield, which indicates that the cattle model mirrors the findings in other mammalian species models, such as mice. Our results show that SAM has a smaller Firmicute-to-Bacteroidota ratio, which indicates that SAM has a potential to be more efficient in nutrient utilization than LAM.
Our results showed that SAM has a greater Inverse Simpson index, which indicates a greater diversity of bacteria. This difference in diversity of bacteria can be attributed to the concentration of nutrients available within each microenvironment, for example, in the liquid fraction soluble nutrients are the main substrates which support bacterial growth, and thus they can correspond from 5 to more than 50% of protein [
18] and non-fiber carbohydrates [
19] from the feed, depending on the ingredients and processing. While the remaining insoluble fraction of the feed contains a greater diversity of nutrients, such as protein, non-fiber carbohydrates, fiber, and lipids, which can support a greater range of microbial community. These results corroborate data reported by Li et al. [
20] who evaluated the solid, liquid, and epithelium-associated microbiome and found significant ecological differences between the three microenvironments.
In a recent study, Zhao et al. [
21] evaluated the effect of solid- and liquid-associated ruminal microorganisms on fermentation and the microbiome. The authors reported that solid-associated bacteria were enriched for fiber degrading microorganisms, such as
Treponema,
Succinivibrio, and
Ruminococcus. These results corroborate the differential abundance data in our study, in which microorganisms closely related to the
Lachnospiraceae family and the genera
Prevotellaceae,
Methanosphera,
Fibrobacter, and
Treponema had a greater abundance in the solid microenvironment when compared to liquid. Microorganisms of the
Lachnospiraceae family [
22], as well as microorganisms of the genera
Prevotella [
23] and
Fibrobacter [
24], are well known carbohydrate fermenters, with most of the species having α- and β-glucosidase, xylanase, pectin methyl-esterase, β-xylosidase, α- and β-galactosidase, α-amylase, and other enzymes.
The greater abundance of carbohydrate degrading microorganisms observed in the SAM microenvironment is caused by the greater proportion of fiber and non-fiber carbohydrates present in the insoluble fraction of the feed. As mentioned earlier, microorganisms must attach to the insoluble fraction of the feed, which would allow them to take advantage of their enzymes to degrade biopolymers and release substrates, such as glucose and amino acids, that can be used as energy or building blocks for bacterial cell components. An interesting result from our study is that there was no effect of mOAPBs on the in vitro ruminal microbiome. These results align with fermentation parameters that are reported in our companion paper [
10].
mOAPBs have been studied as modulators of the intestine microbiota, with successful modulation of ileal and jejunal microbiota in chickens [
9], rainbow trout [
8,
25], and other livestock animals. Most of these studies reported that mOAPBs increased the abundance of microorganisms in the
Lactobacilliaceae family, which is a diverse family of lactic acid bacteria. They are correlated with improvements in gut health through their modulation of the immune system in the host animal, as well as protection against pathogens [
26]. Yoshimaru et al. [
27] reported that around 35% of the compounds that are microencapsulated are usually released during ruminal fermentation, which would enable them to influence the ruminal microbiome and fermentation. Due to these facts, we have hypothesized that part of the mOAPBs would be released during the ruminal fermentation process and would modulate the ruminal microbiome.
Nevertheless, even with a dosage two times greater than the recommended dose, no effects of mOAPBs on ruminal fermentation and microbiome were observed. The lack of effect of mOAPBs in the in vitro rumen microbiome can be attributed to the utilization of lipid microencapsulation technology in these compounds. The goal of microencapsulation is to protect the active compounds against ruminal fermentation and enable their release in the intestine, where they can function as modulators of the microbiome and immune system.
Thus, we can infer that microencapsulation was efficient in protecting the active compounds against ruminal fermentation and increasing the likelihood of their release in the intestine of the animal. However, the efficiency of lipid microencapsulation is highly dependent on the environment pH, where lower pHs can result in greater emulsification of the lipids used for encapsulation and hence liberate the active compounds on the rumen with potential effects in the ruminal microbiome [
28]. The range of pH in our study varied from 5.98 to 6.45, which may not have disrupted the lipid encapsulation and hence may not have released the active compounds within it. Consequently, further research using experimental models with a greater range of pHs is required to further assess the efficiency of lipid microencapsulation and evaluate the effects of mOAPBs as modulators of the ruminal microbiome.
5. Conclusions
The tested mOAPBs did not modulate the microbiome of either the solid or liquid microenvironment of the in vitro ruminal simulator, which may be explained by the high resistance of the microencapsulation technology within the pH range of the experiment. The liquid and solid microenvironment differed in alpha- and beta-diversity, which indicates that the profile of the microbial population in the two microenvironments is different. In addition, the solid microenvironment had a greater concentration of microorganisms that are closely related to the Lachnospiraceae family and genera Prevotellaceae, Methanosphera, Fibrobacter, and Treponema, which are well known ruminal carbohydrate fermenters.
Further investigations should focus on the supplementation of mOAPBs in scenarios where the diet would entice different ruminal pHs, such as pasture or high-concentrate diet. Such experiments would allow dairy nutritionists to better understand the efficiency of the microencapsulation process and the possible side effects of these biocompounds. In addition, further investigations into the effects of mOAPBs in the intestine of dairy cows are required, to better understand the site and mode of action of these biocompounds in improving gut health and production.





{kind=link}
{kind=link}