
LED White Light Treatment Delays Postharvest Senescence of ‘Zaosu’ Pear Fruit with Inhibited Chlorophyll Degradation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Color, Weight Loss, and Firmness
2.3. Ascorbic Acid Content
2.4. Sugar-to-Acid Ratio
2.5. Respiratory Intensity and Ethylene Production
2.6. Chlorophyll Content and Related Enzyme Activities
2.7. Fluorescence Quantitative PCR
2.8. Statistical Analysis
3. Results
3.1. Effect of LED White Light on the Postharvest Quality of Pear Fruit
3.2. Effect of LED White Light on Respiratory Intensity and Ethylene Production of Pear Fruit
3.3. Effect of LED White Light on the Expression of Genes Related to the Ethylene Biosynthesis Pathway in Pear Fruit
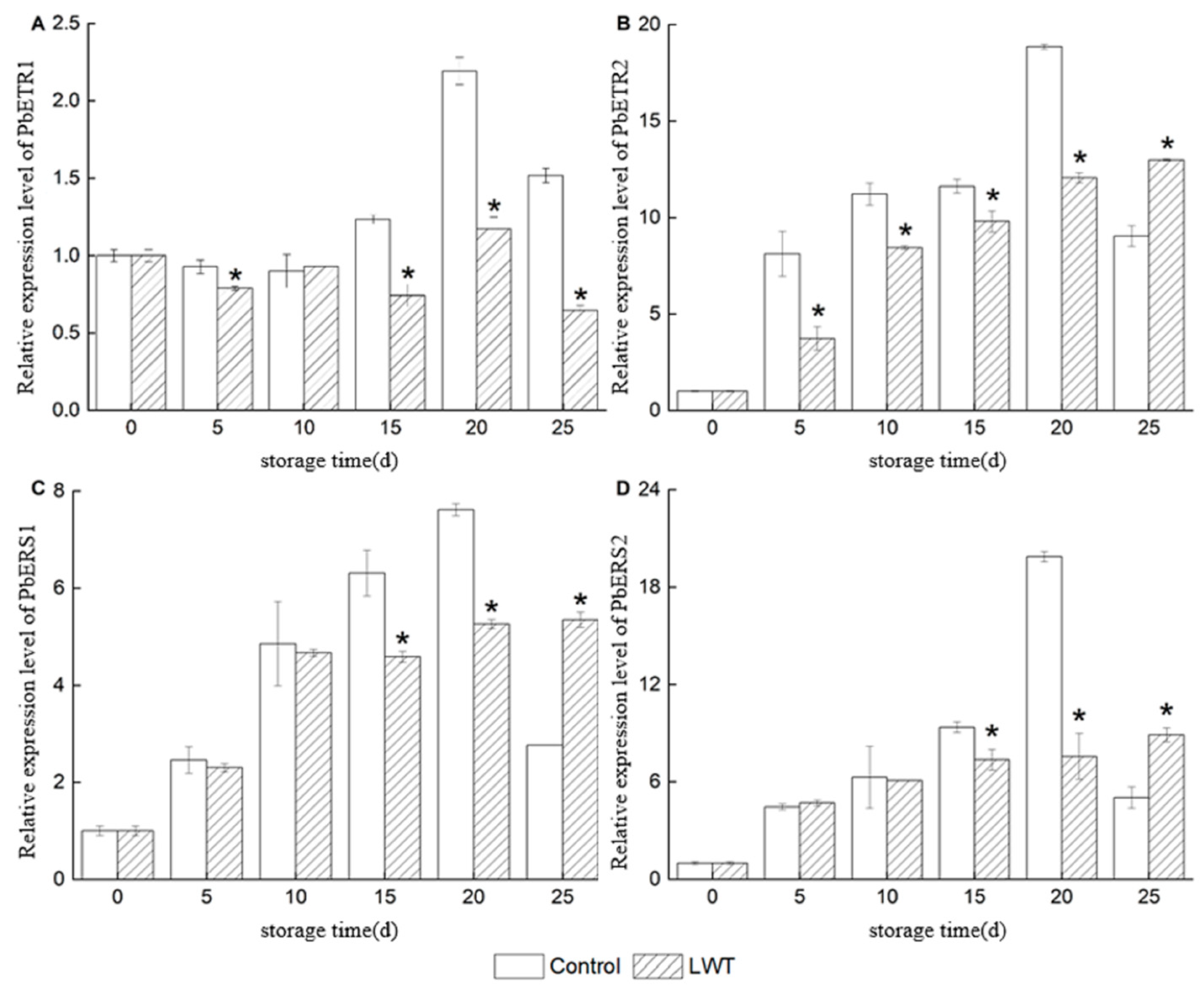
3.4. Effect of LED White Light on the Expression of Ethylene Signal Transduction Pathway-Related Genes in Pear Fruit
3.5. Effect of LED White Light on the Color Change and Chlorophyll Content of Pear Fruit
3.6. Effect of LED White Light on the Activities of Enzymes Related to Chlorophyll Synthesis and Metabolism in Pear Fruit
3.7. Effect of LED White Light on the Activities and Gene Expression of Enzymes Related to Chlorophyll Metabolism in Pear Fruit
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wei, D.M.; Xu, J.; Dong, F.S.; Liu, X.G.; Wu, X.H.; Zheng, Y.Q. Penicillium and patulin distribution in pears contaminated with Penicillium expansum. Determination of patulin in pears by UHPLC-MS/MS. J. Integr. Agric. 2017, 16, 1645–1651. [Google Scholar] [CrossRef]
- Saquet, A.A. Storage of pears. Sci. Hortic. 2019, 246, 1009–1016. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Wang, T.; Fang, C.; Wang, J. Methyl salicylate delays peel yellowing of ‘Zaosu’ pear (Pyrus bretschneideri) during storage by regulating chlorophyll metabolism and maintaining chloroplast ultrastructure. J. Sci. Food Agric. 2019, 99, 4816–4824. [Google Scholar] [CrossRef] [PubMed]
- Armitage, A.D.; Cockerton, H.M.; Sreenivasaprasad, S.; Woodhall, J.; Lane, C.R.; Harrison, R.J.; Clarkson, J.P. Genomics evolutionary history and diagnostics of the Alternaria alternata species group including apple and Asian pear pathotypes. Front. Microbiol. 2020, 10, 3124. [Google Scholar] [CrossRef] [PubMed]
- Kaewsuksaeng, S.; Tatmala, N.; Srilaong, V.; Pongprasert, N. Postharvest heat treatment delays chlorophyll degradation and maintains quality in Thai lime (Citrus aurantifolia Swingle cv. Paan) fruit. Postharvest Biol. Technol. 2015, 100, 1–7. [Google Scholar] [CrossRef]
- Villarreal, N.M.; Rosli, H.G.; Martínez, G.A.; Civello, P.M. Polygalacturonase activity and expression of related genes during ripening of strawberry cultivars with contrasting fruit firmness. Postharvest Biol. Technol. 2008, 47, 141–150. [Google Scholar] [CrossRef]
- Zhan, L.; Li, J.; Huang, W.; Song, C.; Li, J.; Pang, L.; Li, Y. Light irradiation affects the total antioxidant capacity, total phenolic compounds, phenolic acids, and related enzyme activities of minimally processed spinach (Spinacia oleracea L.). J. Food Process. Preserv. 2020, 44, e14825. [Google Scholar] [CrossRef]
- Zhang, X.-J.; Zhang, M.; Chitrakar, B.; Devahastin, S.; Guo, Z. Novel combined use of red-white LED illumination and modified atmosphere packaging for maintaining storage quality of postharvest pakchoi. Food Bioprocess Technol. 2022, 15, 590–605. [Google Scholar] [CrossRef]
- Bae, J.M.; Lee, D.U.; Jeong, M.C.; Choi, J.H. Change of quality characteristics in fresh-cut ‘Romaine’ lettuce by heat treatment. Korean J. Food Preserv. 2016, 23, 27–33. [Google Scholar] [CrossRef]
- Youn, K.-S.; Park, E.-H.; Yoon, K.Y. Quality characteristics and antioxidant activity of bitter melon (Momordica charantia L.) dried by different methods. Korean J. Food Preserv. 2019, 26, 185–193. [Google Scholar] [CrossRef]
- Tkaczewska, J. Peptides and protein hydrolysates as food preservatives and bioactive components of edible films and coatings—A review. Trends Food Sci. Technol. 2020, 106, 298–311. [Google Scholar] [CrossRef]
- Lum, G.B.; DeEll, J.R.; Hoover, G.J.; Subedi, S.; Shelp, B.J.; Bozzo, G.G. 1-Methylcylopropene and controlled atmosphere modulate oxidative stress metabolism and reduce senescence-related disorders in stored pear fruit. Postharvest Biol. Technol. 2017, 129, 52–63. [Google Scholar] [CrossRef]
- García-Martín, J.F.; Olmo, M.; García, J.M. Effect of ozone treatment on postharvest disease and quality of different citrus varieties at laboratory and at industrial facility. Postharvest Biol. Technol. 2018, 137, 77–85. [Google Scholar] [CrossRef]
- Han, Q.; Gao, H.; Chen, H.; Fang, X.; Wu, W. Precooling and ozone treatments affects postharvest quality of black mulberry (Morus nigra) fruits. Food Chem. 2017, 221, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, Y.; Lin, H.; Lin, M.; Li, H.; Yuan, F.; Chen, Y.; Xiao, J. Effects of paper containing 1-MCP postharvest treatment on the disassembly of cell wall polysaccharides and softening in Younai plum fruit during storage. Food Chem. 2018, 264, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-García, P.F.; Ventura-Aguilar, R.I.; del Río-García, J.C.; Hernández-López, M.; Guillén-Sánchez, D.; Salazar-Piña, D.A.; Ramos-García, M.d.L.; Bautista-Baños, S. Edible chitosan/propolis coatings and their effect on ripening, development of Aspergillus flavus, and sensory quality in fig fruit, during controlled storage. Plants 2021, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Cackett, L.; Luginbuehl, L.H.; Schreier, T.B.; Lopez-Juez, E.; Hibberd, J.M. Chloroplast development in green plant tissues: The interplay between light, hormone, and transcriptional regulation. New Phytol. 2022, 233, 2000–2016. [Google Scholar] [CrossRef]
- Kim, B.-S.; Youm, S.; Kim, Y.-K. Sterilization of harmful microorganisms in hydroponic cultivation using an ultraviolet LED light source. Sens. Mater. 2020, 32, 3773–3785. [Google Scholar] [CrossRef]
- Shi, J.; Gao, L.; Zuo, J.; Wang, Q.; Wang, Q.; Fan, L. Exogenous sodium nitroprusside treatment of broccoli florets extends shelf life, enhances antioxidant enzyme activity, and inhibits chlorophyll-degradation. Postharvest Biol. Technol. 2016, 116, 98–104. [Google Scholar] [CrossRef]
- Xu, F.; Tang, Y.; Dong, S.; Shao, X.; Wang, H.; Zheng, Y.; Yang, Z. Reducing yellowing and enhancing antioxidant capacity of broccoli in storage by sucrose treatment. Postharvest Biol. Technol. 2016, 112, 39–45. [Google Scholar] [CrossRef]
- Kasim, M.U.; Kasim, R. While continuous white LED lighting increases chlorophyll content (SPAD), green LED light reduces the infection rate of lettuce during storage and shelf-life conditions. J. Food Process. Preserv. 2017, 41, e13266. [Google Scholar] [CrossRef]
- Skrovankova, S.; Mlcek, J.; Sochor, J.; Baron, M.; Kynicky, J.; Jurikova, T. Determination of ascorbic acid by electrochemical techniques and other methods. Int. J. Electrochem. Sci. 2015, 3, 2421–2431. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, L.; Zhang, L.; Lv, H.; He, Q.; Guo, L.; Zhang, X.; He, H.; Ren, S.; Zhang, N.; et al. Melatonin promotes carotenoid biosynthesis in an ethylene-dependent manner in tomato fruits. Plant Sci. 2020, 298, 110580. [Google Scholar] [CrossRef] [PubMed]
- Esteban, R.; García-Plazaola, J.I.; Hernández, A.; Fernández-Marín, B. On the recalcitrant use of Arnon’s method for chlorophyll determination. New Phytol. 2018, 217, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, D.; Zhang, X.; Feng, X.; Mi, H.; Ge, Y.; Lv, J. Comprehensive responses of aroma production in ‘Benihoppe’ strawberry to low oxygen associated with the changes of key gene expressions and energy levels. J. Sci. Food Agric. 2023, 103, 1856–1863. [Google Scholar] [CrossRef] [PubMed]
- Charoenchongsuk, N.; Ikeda, K.; Itai, A.; Oikawa, A.; Murayama, H. Comparison of the expression of chlorophyll-degradation-related genes during ripening between stay-green and yellow-pear cultivars. Sci. Hortic. 2015, 181, 89–94. [Google Scholar] [CrossRef]
- Cheng, Y.; Dong, Y.; Yan, H.; Ge, W.; Shen, C.; Guan, J.; Liu, L.; Zhang, Y. Effects of 1-MCP on chlorophyll degradation pathway-associated genes expression and chloroplast ultrastructure during the peel yellowing of Chinese pear fruits in storage. Food Chem. 2012, 135, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, L.; Fan, C.; Wang, P.; Cui, M.; Liu, L.; Yang, H.; Wang, J. Impact of methyl salicylate on storage quality, ethylene action, and protein profiling of ‘Zaosu’ pear (Pyrus bretschneideri). Sci. Hortic. 2020, 264, 109196. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Schmittgen. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lado, J.; Alós, E.; Rodrigo, M.J.; Zacarías, L. Light avoidance reduces ascorbic acid accumulation in the peel of Citrus fruit. Plant Sci. 2015, 231, 138–147. [Google Scholar] [CrossRef]
- Chen, X.-L.; Wang, L.-C.; Li, T.; Yang, Q.-C.; Guo, W.-Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 6926. [Google Scholar] [CrossRef] [PubMed]
- Hasperué, J.H.; Rodoni, L.M.; Guardianelli, L.M.; Chaves, A.R.; Martínez, G.A. Use of LED light for Brussels sprouts postharvest conservation. Sci. Hortic. 2016, 213, 281–286. [Google Scholar] [CrossRef]
- Lv, J.; Zhang, M.; Zhang, J.; Ge, Y.; Li, C.; Meng, K.; Li, J. Effects of methyl jasmonate on expression of genes involved in ethylene biosynthesis and signaling pathway during postharvest ripening of apple fruit. Sci. Hortic. 2018, 229, 157–166. [Google Scholar] [CrossRef]
- Chen, A.; Liu, L.; Liu, X.; Fu, Y.; Li, J.; Zhao, J.; Hou, X. Exploring the differential stages of the pigment metabolism by pre-harvest bagging and post-harvest ethylene de-greening of Eureka lemon peel. PeerJ 2021, 9, e11504. [Google Scholar] [CrossRef]
- Vergara-Domínguez, H.; Ríos, J.J.; Gandul-Rojas, B.; Roca, M. Chlorophyll catabolism in olive fruits (var. Arbequina and Hojiblanca) during maturation. Food Chem. 2016, 212, 604–611. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Gene ID | Primer Sequence |
|---|---|---|
| PbNYC | JN167997 | F: GGCATAGCATGTGATGTTTG |
| R: GCACCAGCATTGTTTATCC | ||
| PbNOL | JN167996 | F: CCGTGAGGCAATAAAAATGATG |
| R: CGCTTTGTTGCCCCGTAT | ||
| PbCLH1 | JN168001 | F: CAACGTTCATAGACTTCCCCTACG |
| R: GGAGAGTACAAGGCAAGAGCTGC | ||
| PbSGR1 | JN168000 | F: ATTCAACAAGGTGGAGTGCTGG |
| R: GCCGCTGTTGTTTTCCTGG | ||
| PbPPH | JN168999 | F: CTGCTGCATCATTGGCCTC |
| R: CTCGTAATATGGAGCTTCAGGC | ||
| PbPAO | JN168998 | F: GCGACATCGACGAGCAAGTT |
| R: TCGAAAGCGACCCACGATTC | ||
| PbRCCR | JN168002 | F: TTCATAGACTTCCCCTACGTGTCG |
| R: GGAGAGTACAAGGCAAGAGCTGC | ||
| EF1-α | AH009876 | F: CAAGTATGCCTGGGTGCTTG |
| R: TCAGCCTGTGAAGTTCCAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mi, H.; Zhou, X.; Yang, J.; Chen, J.; Liu, B. LED White Light Treatment Delays Postharvest Senescence of ‘Zaosu’ Pear Fruit with Inhibited Chlorophyll Degradation. Horticulturae 2024, 10, 32. https://doi.org/10.3390/horticulturae10010032
Mi H, Zhou X, Yang J, Chen J, Liu B. LED White Light Treatment Delays Postharvest Senescence of ‘Zaosu’ Pear Fruit with Inhibited Chlorophyll Degradation. Horticulturae. 2024; 10(1):32. https://doi.org/10.3390/horticulturae10010032
Chicago/Turabian StyleMi, Hongbo, Xu Zhou, Jun Yang, Jingxin Chen, and Bin Liu. 2024. "LED White Light Treatment Delays Postharvest Senescence of ‘Zaosu’ Pear Fruit with Inhibited Chlorophyll Degradation" Horticulturae 10, no. 1: 32. https://doi.org/10.3390/horticulturae10010032
APA StyleMi, H., Zhou, X., Yang, J., Chen, J., & Liu, B. (2024). LED White Light Treatment Delays Postharvest Senescence of ‘Zaosu’ Pear Fruit with Inhibited Chlorophyll Degradation. Horticulturae, 10(1), 32. https://doi.org/10.3390/horticulturae10010032