
Evaluation of Effect of Brassinolide in Brassica juncea Leaves under Drought Stress in Field Conditions

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Physiological Parameters
2.3. Enzyme Extraction and Assay
2.4. Non-Enzymatic Estimations
2.5. Extraction and Estimation of Oxidative Stress Indicators
2.6. Statistical Analysis
3. Results
3.1. Physiological Parameters
3.2. Oxidative Stress Indicators
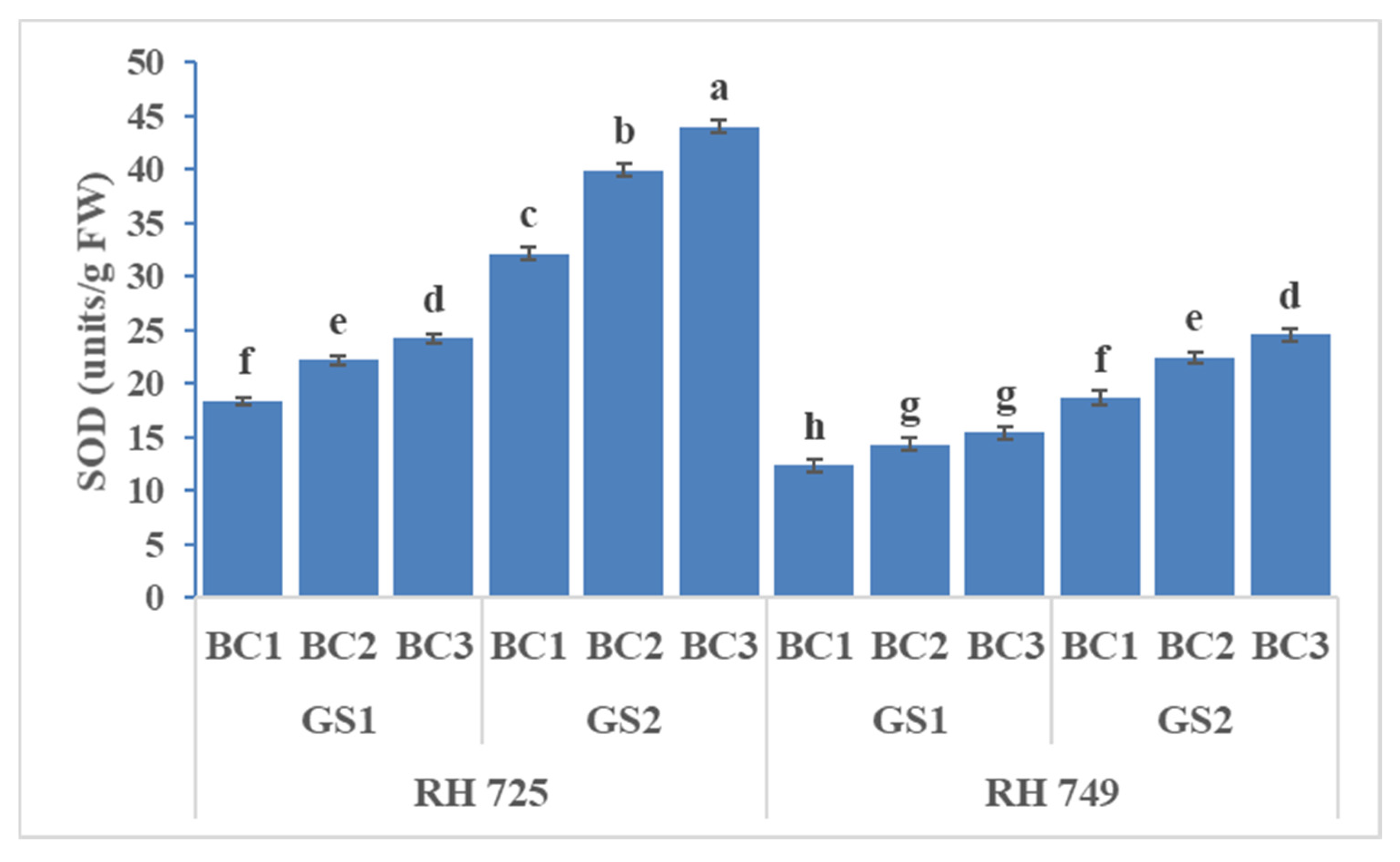
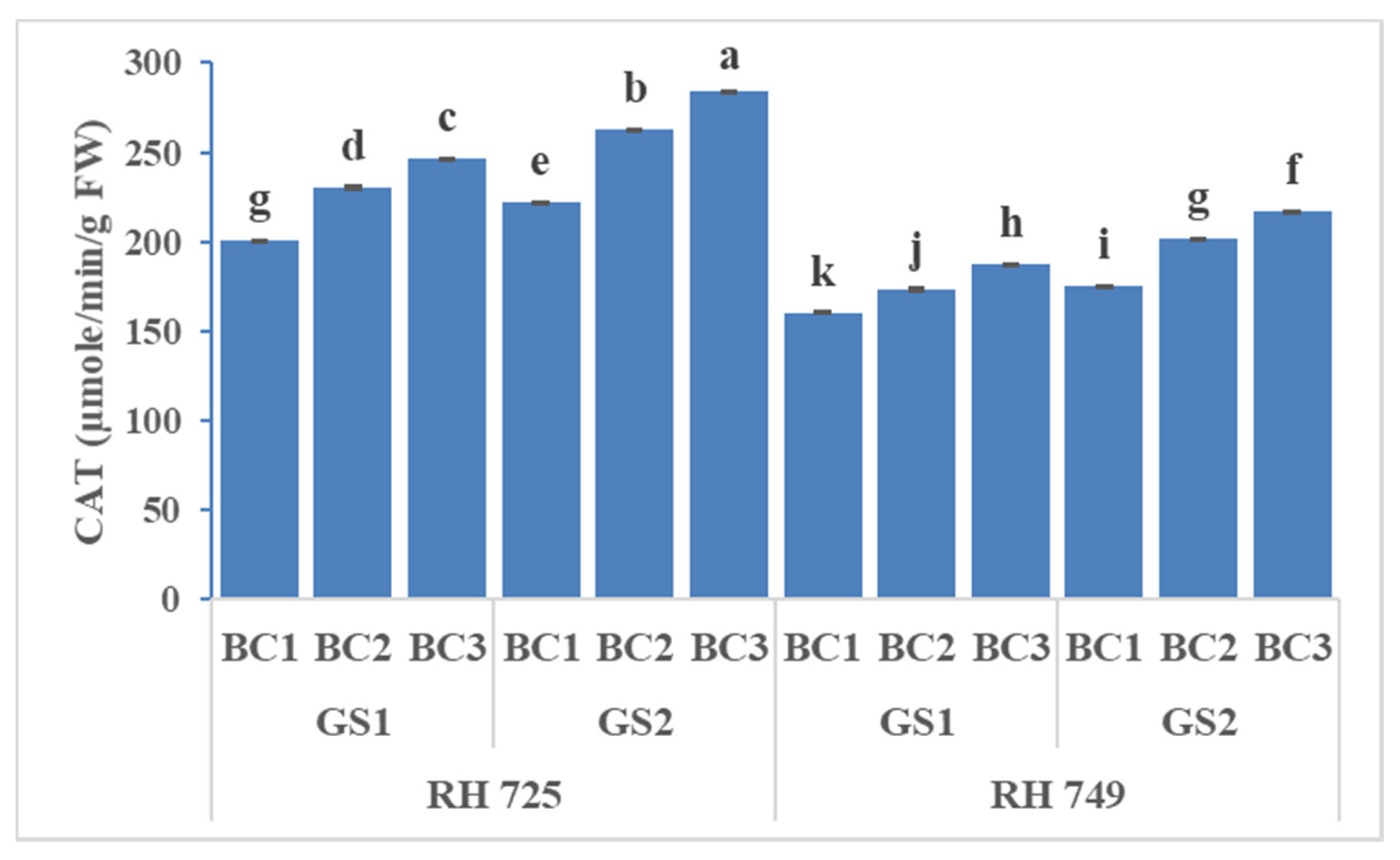
3.3. Enzymatic Antioxidants
3.4. Non-Enzymatic Antioxidants
3.5. Correlation Analysis among Different Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, M.; Avtar, R.; Lakra, N.; Hooda, E.; Singh, V.K.; Bishnoi, M.; Kumari, N.; Punia, R.; Kumar, N.; Choudhary, R.R. Genetic and Proteomic Basis of Sclerotinia Stem Rot Resistance in Indian Mustard [Brassica juncea (L.) Czern & Coss.]. Genes 2021, 12, 1784. [Google Scholar]
- Norton, R.; Burton, W.; Salisbury, P. Canola quality Brassica juncea for Australia. In Proceeding of 4th International Crop Science Congress, Gosford, Australia, 26 September–1 October 2004; p. 5. [Google Scholar]
- Mahmood, T.; Ashraf, M.; Shahbaz, M. Does exogenous application of glycine-betaine as a pre-sowing seed treatment improve growth and regulate some key physiological attributes in wheat plants grown under water deficit conditions? Pak. J. Bot. 2009, 41, 1291–1302. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Anjum, N.A.; Sofo, A.; Scopa, A.; Roychoudhury, A.; Gill, S.S.; Iqbal, M.; Lukatkin, A.S.; Pereira, E.; Duarte, A.C.; Ahmad, I. Lipids and proteins—Major targets of oxidative modifications in abiotic stressed plants. Environ. Sci. Pollut. Res. 2014, 22, 4099–4121. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Avtar, R.; Pal, A.; Punia, R.; Singh, V.K.; Bishnoi, M.; Singh, A.; Choudhary, R.R.; Mandhania, S. Genotype-Specific Antioxidant Responses and Assessment of Resistance Against Sclerotinia sclerotiorum Causing Sclerotinia Rot in Indian Mustard. Pathogens 2020, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- El-Mashad, A.A.A.; Mohamed, H.I. Brassinolide alleviates salt stress and increases antioxidant activity of cowpea plants (Vigna sinensis). Protoplasma 2012, 249, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.; Noctor, G. Oxygen processing in photosynthesis: Regulation and signaling. New Phytol. 2000, 146, 359–388. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Arora, N.; Bhardwaj, R.; Sharma, P.; Arora, H.K. 28-Homobrassinolide alleviates oxidative stress in salt-treated maize (Zea mays L.) plants. Braz. J. Plant Physiol. 2008, 20, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hasan, S.A.; Hayat, Q.; Ahmad, A. Brassinosteroids protect Lycopersicon esculentum from cadmium toxicity applied as shotgun approach. Protoplasma 2010, 239, 3–14. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.A.; Hayat, S.; Ali, B.; Ahmad, A. 28-homobrassinolide protects chickpea (Cicer arietinum) from cadmium toxicity by stimulating antioxidants. Environ. Pollut. 2008, 151, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Fariduddin, Q.; Khanam, S.; Hasan, S.A.; Ali, B.; Hayat, S.; Ahmad, A. Effect of 28-homobrassinolide on the drought stress-induced changes in photosynthesis and antioxidant system of Brassica juncea L. Acta Physiol.Plant 2009, 31, 889–897. [Google Scholar] [CrossRef]
- Ali, B.; Hayat, S.; Ahmad, A. 28-Homobrassinolide ameliorates the saline stress in chickpea (Cicer arietinum L.). Environ. Exp. Bot. 2007, 59, 217–223. [Google Scholar] [CrossRef]
- Behnamnia, M.; Kalantari, K.M.; Ziaie, J. The effects of brassinosteroid on the induction of biochemical changes in Lycopersicon esculentum under drought stress. Turk. J. Botany 2009, 33, 417–428. [Google Scholar]
- Zhang, M.; Zhai, Z.; Tian, X.; Duan, L.; Li, Z. Brassinolide alleviated the adverse effect of water deficits on photosynthesis and the antioxidant of soybean (Glycine max L.). Plant Growth Regul. 2008, 56, 257–264. [Google Scholar] [CrossRef]
- Shen, X.Y.; Dai, J.Y.; Hu, A.C.; Gu, W.L.; He, R.Y.; Zheng, B. Studies on physiological effects of brassinolide on drought resistance in maize. J. Shenyang Agric. Univ. 1990, 21, 191–195. [Google Scholar]
- Gill, M.B.; Cai, K.; Zhang, G.; Zeng, F. Brassinolide alleviates the drought-induced adverse effects in barley by modulation of enzymatic antioxidants and ultrastructure. Plant Growth Regul. 2017, 82, 447–455. [Google Scholar] [CrossRef]
- Silva, F.V.D.F.; Mendes, B.D.S.; Rocha, M.D.S.; Brito, J.F.D.; Beltrão, N.E.D.M.; Sofiatti, V. Photosynthetic pigments and gas exchange in castor bean under conditions of above the optimal temperature and high CO2. Acta Sci. Agron. 2015, 37, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Chem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Sinha, A.K. Calorimetric assay of catalase. Anal. Chem. 1972, 47, 389–395. [Google Scholar]
- Shannon, L.M.; Key, E.; Law, J.Y. Peroxidase isoenzymes from horse reddish roots: Isolation and physical properties. J. Biol. Chem. 1966, 241, 2166–2172. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Roe, J.H. Chemical determination of ascorbic dehydroascorbic and diketogluconic acids. Methods Biochem. Anal. 1954, 1, 115–139. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Gong, J.R.; Zhao, A.F.; Huang, Y.M.; Zhang, X.S.; Zhang, C.L. Water relations, gas exchange, photochemical efficiency, and peroxidative stress of four plant species in the Heihe drainage basin of northern China. Photosynthetica 2006, 44, 355–364. [Google Scholar] [CrossRef]
- Ali, M.A.; Jabran, K.; Awan, S.I.; Abbas, A.; Zulkiffal, M.; Acet, T.; Farooq, J.; Rehman, A. Morpho-physiological diversity and its implications for improving drought tolerance in grain sorghum at different growth stages. Aust. J. Crop Sci. 2011, 5, 311–320. [Google Scholar]
- Kumari, A.; Avtar, R.; Kumari, N.; Jattan, M.; Rani, B. Screening for drought tolerance in Indian mustard (Brassica juncea L.) genotypes based on yield contributing characters and physiological parameters. J. Oilseed Brassica 2019, 10, 1–7. [Google Scholar]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Dehghan, M.; Balouchi, H.; Yadavi, A.; Zare, E. Improve wheat (Triticum aestivum) performance by brassinolide application under different irrigation regimes. S. Afr. J. Bot. 2020, 130, 259–267. [Google Scholar] [CrossRef]
- Chen, L.; Yang, H.; Fang, Y.; Guo, W.; Chen, H.; Zhang, X.; Dai, W.; Chen, S.; Hao, Q.; Yuan, S.; et al. Overexpression of GmMYB14 improves high-density yield and drought tolerance of soybean through regulating plant architecture mediated by the brassinosteroid pathway. Plant Biotechnol. J. 2021, 19, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Tůmová, L.; Tarkowská, D.; Řehořová, K.; Marková, H.; Kočová, M.; Rothová, O.; Čečetka, P.; Holá, D. Drought-tolerant and drought-sensitive genotypes of maize (Zea mays L.) differ in contents of endogenous brassinosteroids and their drought-induced changes. PLoS ONE 2018, 13, e0197870. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, M.; Jahanbin, S.; Latifmanesh, H.; Farajee, H.; Mirshekari, A. Effect of brassinolide foliar application on some physiological and agronomic characteristics of sunflower (Helianthus annuus L.) under drought stress conditions. J. Crop Prod. 2021, 14, 31–48. [Google Scholar]
- Jangid, K.K.; Dwivedi, P. Physiological and biochemical changes by nitric oxide and brassinosteroid in tomato (Lycopersicon esculentum Mill.) under drought stress. Acta Physiol Plant. 2017, 39, 73. [Google Scholar] [CrossRef]
- Verma, J.; Kakralya, B.L.; Jakhar, M.L. Effect of Brassinolide on physiological aspects of Chick pea (Cicer arietinum L.) under drought conditions. J. Plant Sci. Res. 2012, 28, 151–155. [Google Scholar]
- Sun, Y.; Fan, X.Y.; Cao, D.M.; Tang, W.; He, K.; Zhu, J.Y. Integration of brassinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis. Dev. Cell. 2010, 19, 765–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.J.; Huang, L.F.; Zhou, Y.H.; Mao, W.H.; Shi, K.; Wu, J.X.; Asami, T.; Chen, Z.; Yu, J.Q. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 2009, 230, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Akram, N.A.; Arteca, R.N.; Foolad, M.R. The physiological, biochemical and molecular roles of brassinosteroids and salicylic acid in plant processes and salt tolerance. Crit. Rev. Plant Sci. 2010, 29, 162–190. [Google Scholar] [CrossRef]
- Li, L.; Van Staden, J.; Jager, A.K. Effects of plant growth regulators on the antioxidant system in seedlings of two maize cultivars subjected to water stress. Plant Growth Regul. 1998, 25, 81–87. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, Y.X.; Hou, H.; Jiang, Q.; Chen and Tang, R.H. Chilling acclimation induced changes in the distribution of H2O2 and antioxidant system of strawberry leaves. Agric. J. 2008, 3, 286–291. [Google Scholar]
- Shahbaz, M.; Ashraf, M.; Athar, H. Dose exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Kumari, S.; Thakur, A. The Effects of Water Stress and Brassinosteroid on Apple Varieties. Int. J. Econ. Plants 2019, 6, 1–6. [Google Scholar] [CrossRef]
- Tyagi, A.; Santha, I.M.; Mehta, S.L. Effect of water stress on proline content and transcript levels in Lathyrus sativus. Indian J. Biochem. Biophys. 1999, 36, 207–210. [Google Scholar]
- Smirnoff, N. Ascorbate biosynthesis and function in photoprotection. Philos. Trans. R Soc. Lond. B Biol. Sci. 2000, 355, 1455–1464. [Google Scholar] [CrossRef]
- Bajji, M.; Lutts, S.; Kinet, J.M. Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum Desf.) cultivars performing differently in arid conditions. Plant Sci. J. 2001, 160, 669–681. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Alyemeni, M.N.; Al-Quwaiz, S.M. Effect of 28-homobrassinolide on the performance of sensitive and resistant varieties of Vigna radiata. Saudi J. Biol. Sci. 2016, 23, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Ozdemir, F.; Bor, M.; Demiral, T.; Turkan, I. Effects of 24- epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul. 2004, 42, 203–211. [Google Scholar] [CrossRef]
- Kumari, N.; Avtar, R.; Kumari, A.; Sharma, B.; Rani, B.; Sheoran, R.K. Antioxidative response of Indian mustard subjected to drought stress. J. Oilseed Brassica 2018, 9, 40–44. [Google Scholar]
- Rani, B.; Madan, S.; Pooja, K.S.; Sharma, K.D.; Kumari, N.; Kumar, A. Mitigating the effect of drought stress on yield in wheat (Triticum aestivum) using arbuscular mycorrhiza fungi (Glomus mosseae). Indian J. Agric. Sci. 2018, 88, 95–100. [Google Scholar]
- Ershova, A.N.; Khripach, V.A. Effect of epibrassinolide on lipid peroxidation in Pisum sativum at normal aeration and under oxygen deficiency. Russ. J. Plant Physiol. 1996, 43, 750–752. [Google Scholar]
- Robinson, J.M.; Bunce, J.A. Influence of Drought-Induced Water Stress on Soybean and Spinach Leaf Ascorbate-Dehydro ascorbate level and Redox Status. Int. J. Plant Sci. 2000, 161, 271–279. [Google Scholar] [CrossRef]
- Ogweno, J.O.; Song, X.S.; Shi, K.; Hu, W.H.; Mao, W.H.; Zhou, Y.H.; Nogués, S. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J. Plant Growth Regul. 2008, 27, 49–57. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A. Singlet oxygen production in photosynthesis. J. Exp. Bot. 2005, 56, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Jungklang, J.; Saengnil, K.; Uthaibutra, J. Effects of water-deficit stress and paclobutrazol on growth, relative water content, electrolyte leakage, proline content and some antioxidant changes in Curcuma alismatifolia Gagnep. cv. Chiang Mai Pink. Saudi J. Biol. Sci. 2017, 24, 1505–1512. [Google Scholar] [CrossRef] [Green Version]
- Houimli, S.I.M.; Denden, M.; Mouhandes, B.D. Effects of 24-epibrassinolide on growth, chlorophyll, electrolyte leakage and proline by pepper plants under NaCl-stress. Eur. Asian J. Biosci. 2010, 4, 96–104. [Google Scholar] [CrossRef]
- Coban, O.; Baydar, N.G. Brassinosteroid effects on some physical and biochemical properties and secondary metabolite accumulation in peppermint (Mentha piperita L.) under salt stress. Ind. Crop. Prod. 2016, 86, 251–258. [Google Scholar] [CrossRef]
- Farooq, M.; Aziz, T.; Wahid, A.; Lee, D.J.; Siddique, K.H. Chilling tolerance in maize: Agronomic and physiological approaches. Crop Pasture Sci. 2009, 60, 501–516. [Google Scholar] [CrossRef]
- Yu, J.Q.; Huang, L.F.; Hu, W.H.; Zhou, Y.H.; Mao, W.H.; Ye, S.F.; Nogue’s, S. A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp. Bot. 2004, 55, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.; Fariduddin, Q.; Hayat, S.; Hasan, S.A.; Ahmad, A. Protective responses of 28-homobrssinolide in cultivars of Triticumaestivum with different levels of nickel. Arch. Environ. Contam. Toxicol. 2011, 60, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Asami, T. Suppression of Wolffiaarrhiza growth by brassinazole, an inhibitor of brassinosteroid biosynthesis and its restoration by endogenous 24-epibrassinolide. Phytochemistry 2005, 66, 1787–1796. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SV | Df | Mean Squares | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOD | POX | CAT | ASA | PRO | CC | H2O2 | MDA | EL | SC | PR | TR | ||
| C | 1 | 1335.17 ** | 61.96 ** | 27,408.11 ** | 1333.49 ** | 26.87 ** | 22.14 ** | 38,037.56 ** | 59.50 ** | 225.91 ** | 0.65 ** | 147.87 ** | 5.48 ** |
| ST | 1 | 1400.01 ** | 18.30 ** | 6702.90 ** | 2295.71 ** | 46.10 ** | 18.16 ** | 24,628.80 ** | 44.94 ** | 2334.83 ** | 0.29 ** | 98.01 ** | 59.78 ** |
| C × ST | 1 | 190.35 ** | 0.40 | 88.59 ** | 19.90 ** | 0.61 | 0.79 | 150.79 ** | 8.05 ** | 8.17 ** | 0.01 ** | 12.39 ** | 0.08 ** |
| BC | 2 | 137.58 ** | 31.86 ** | 5914.39 ** | 1068.52 ** | 11.94 ** | 7.58 ** | 18,979.63 ** | 178.78 ** | 242.52 ** | 0.14 ** | 98.06 ** | 6.72 ** |
| C × BC | 2 | 14.73 ** | 1.79 | 320.08 ** | 33.10 ** | 0.51 | 0.47 | 178.68 ** | 0.84 | 4.07 * | 0.02 ** | 8.20 ** | 0.20 ** |
| ST × BC | 2 | 15.35 ** | 0.48 | 192.53 ** | 6.57 ** | 0.63 | 0.48 | 124.60 ** | 5.13 ** | 1.26 | 0.01 ** | 2.13 ** | 0.54 ** |
| C × ST × BC | 2 | 1.95 * | 0.01 | 2.95 * | 0.47 | 0.00 | 0.12 | 94.83 ** | 1.09 | 0.29 | 0.00 | 0.46 ** | 0.06 ** |
| Error | 22 | 0.39 | 0.90 | 0.61 | 0.99 | 0.53 | 0.82 | 0.84 | 0.83 | 0.78 | 0.00 | 0.02 ** | 0.01 ** |
| Variables | SOD | POX | CAT | CC | ASA | PRO | H2O2 | MDA | EL | SC | PR | TR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOD | 1.000 | 0.864 ** | 0.911 ** | 0.960 ** | 0.091 | 0.920 ** | −0.909 ** | −0.644* | −0.640 * | 0.948 ** | 0.903 ** | 0.857 ** |
| POX | 1.000 | 0.974 ** | 0.962 ** | 0.494 | 0.901 ** | −0.981 ** | −0.873 ** | −0.220 | 0.945 ** | 0.955 ** | 0.739 ** | |
| CAT | 1.000 | 0.963 ** | 0.442 | 0.916 ** | −0.969 ** | −0.811 ** | −0.338 | 0.983 ** | 0.950 ** | 0.743 ** | ||
| CC | 1.000 | 0.263 | 0.957 ** | −0.978 ** | −0.801 ** | −0.455 | 0.970 ** | 0.959 ** | 0.856 ** | |||
| ASA | 1.000 | 0.118 | −0.355 | −0.481 | 0.657 * | 0.332 | 0.388 ** | −0.172 | ||||
| PRO | 1.000 | −0.953 ** | −0.824 ** | −0.537 | 0.941 ** | 0.927 ** | 0.943 ** | |||||
| H2O2 | 1.000 | 0.874 ** | 0.353 | −0.957 ** | −0.960 ** | −0.832 ** | ||||||
| MDA | 1.000 | 0.025 | −0.774 ** | −0.854 ** | −0.711 ** | |||||||
| EL | 1.000 | −0.434 | −0.304 | −0.666 ** | ||||||||
| SC | 1.000 | 0.970 ** | 0.806 ** | |||||||||
| PR | 1.000 | 0.817 ** | ||||||||||
| TR | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naveen, N.; Kumari, N.; Avtar, R.; Jattan, M.; Ahlawat, S.; Rani, B.; Malik, K.; Sharma, A.; Singh, M. Evaluation of Effect of Brassinolide in Brassica juncea Leaves under Drought Stress in Field Conditions. Horticulturae 2021, 7, 514. https://doi.org/10.3390/horticulturae7110514
Naveen N, Kumari N, Avtar R, Jattan M, Ahlawat S, Rani B, Malik K, Sharma A, Singh M. Evaluation of Effect of Brassinolide in Brassica juncea Leaves under Drought Stress in Field Conditions. Horticulturae. 2021; 7(11):514. https://doi.org/10.3390/horticulturae7110514
Chicago/Turabian StyleNaveen, Naveen, Nisha Kumari, Ram Avtar, Minakshi Jattan, Sushil Ahlawat, Babita Rani, Kamla Malik, Anubhuti Sharma, and Manjeet Singh. 2021. "Evaluation of Effect of Brassinolide in Brassica juncea Leaves under Drought Stress in Field Conditions" Horticulturae 7, no. 11: 514. https://doi.org/10.3390/horticulturae7110514
APA StyleNaveen, N., Kumari, N., Avtar, R., Jattan, M., Ahlawat, S., Rani, B., Malik, K., Sharma, A., & Singh, M. (2021). Evaluation of Effect of Brassinolide in Brassica juncea Leaves under Drought Stress in Field Conditions. Horticulturae, 7(11), 514. https://doi.org/10.3390/horticulturae7110514