
Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Growth
2.2. Seed Biopriming and Plant Growth
2.3. Pigments Quantification
2.4. Carbohydrates Content (TSS and Starch)
2.5. Photosynthetic Performance and Gas Exchange
2.6. Total Phenolic Content
2.7. DPPH Scavenging Activity
2.8. Superoxide (O2−)
2.9. Statistical Analysis
3. Results
3.1. Shoot and Root Length, and Biomass
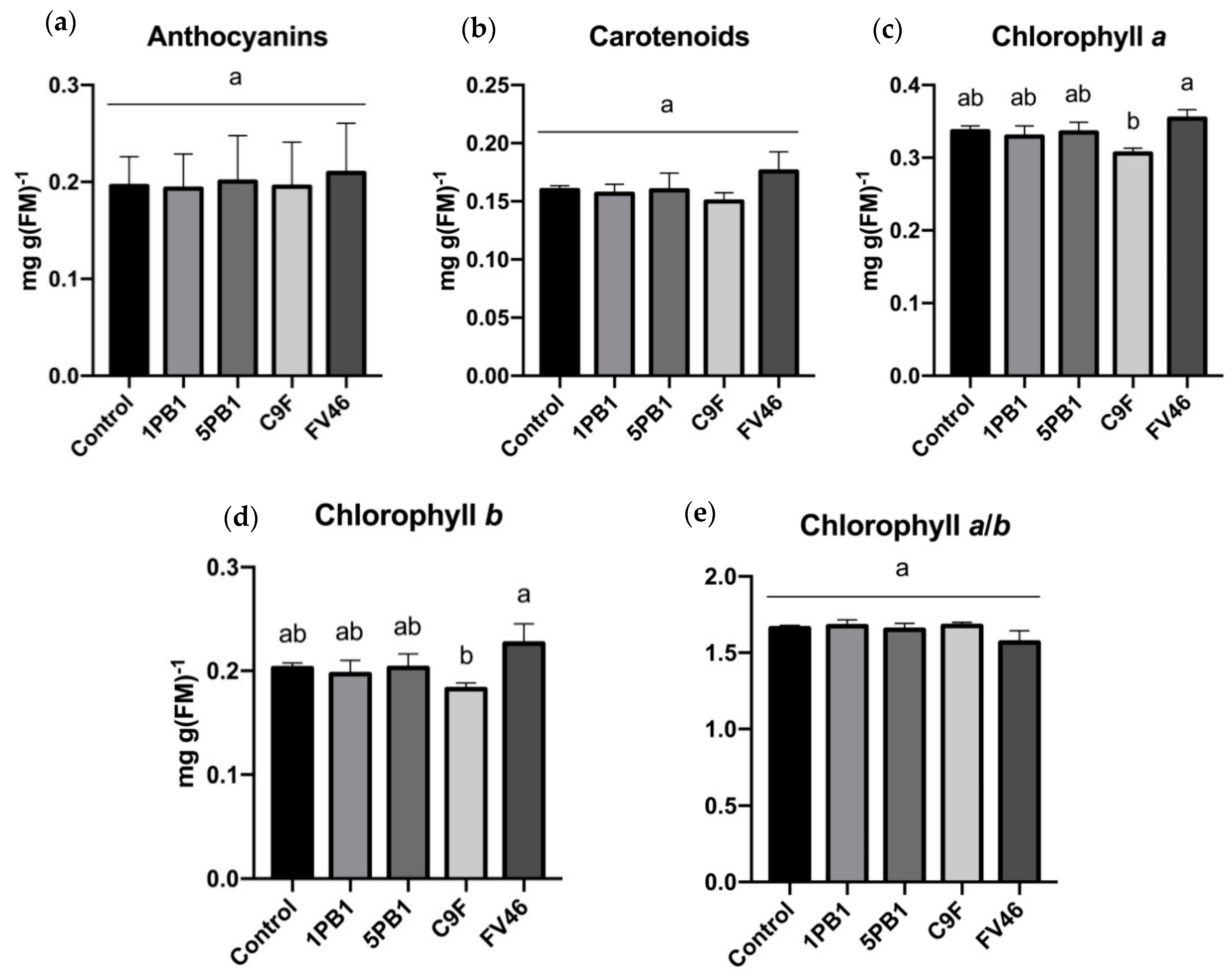
3.2. Pigments Content
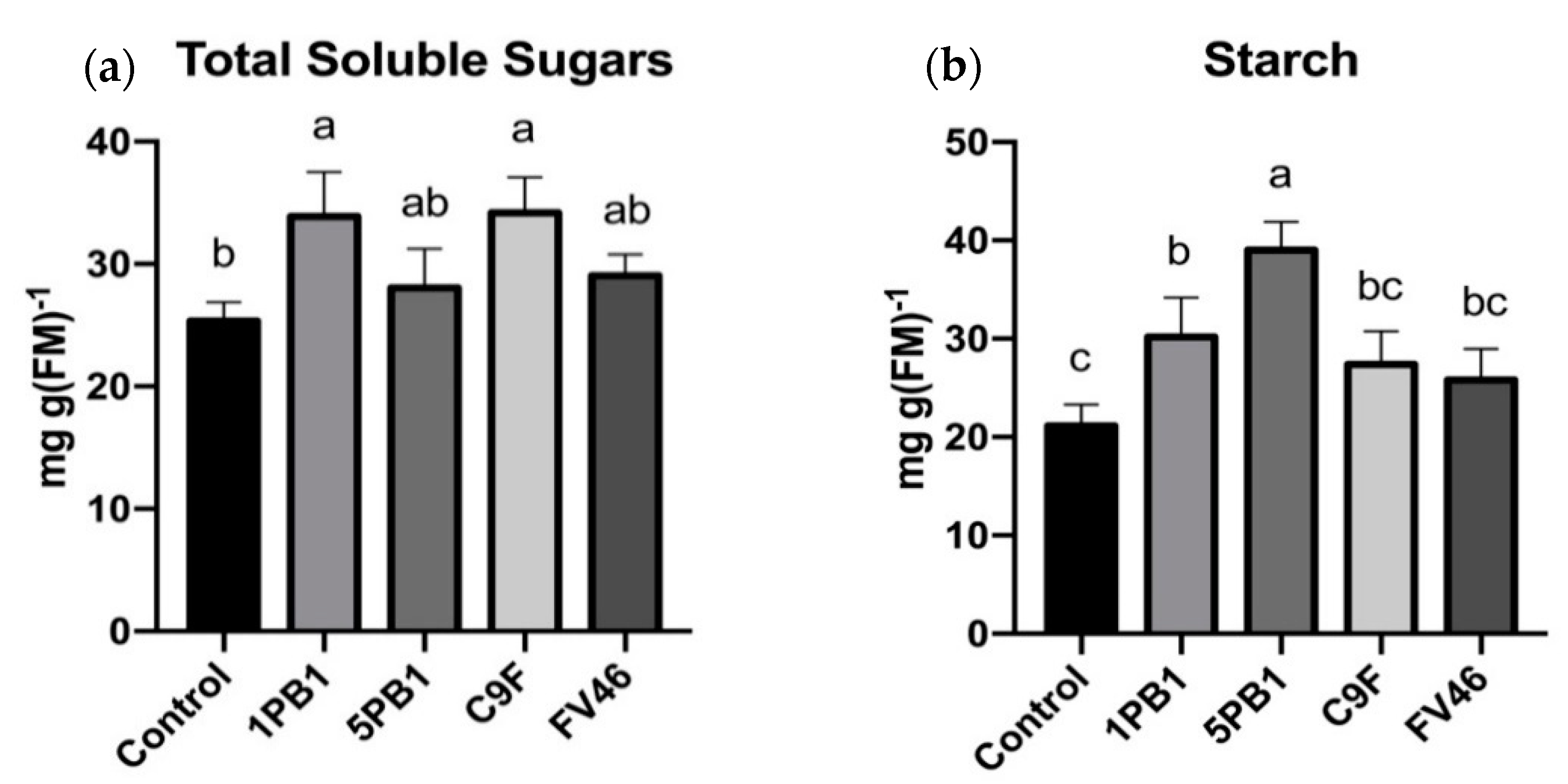
3.3. Carbohydrates Content
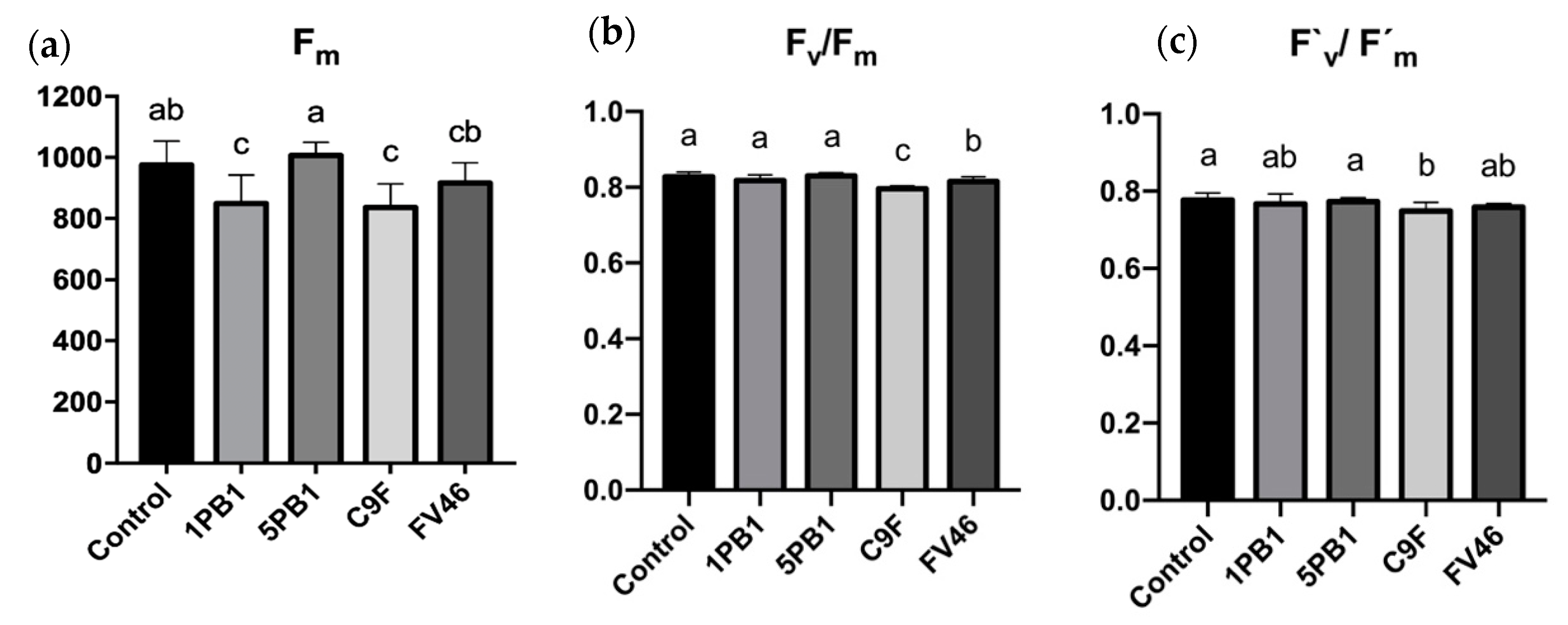
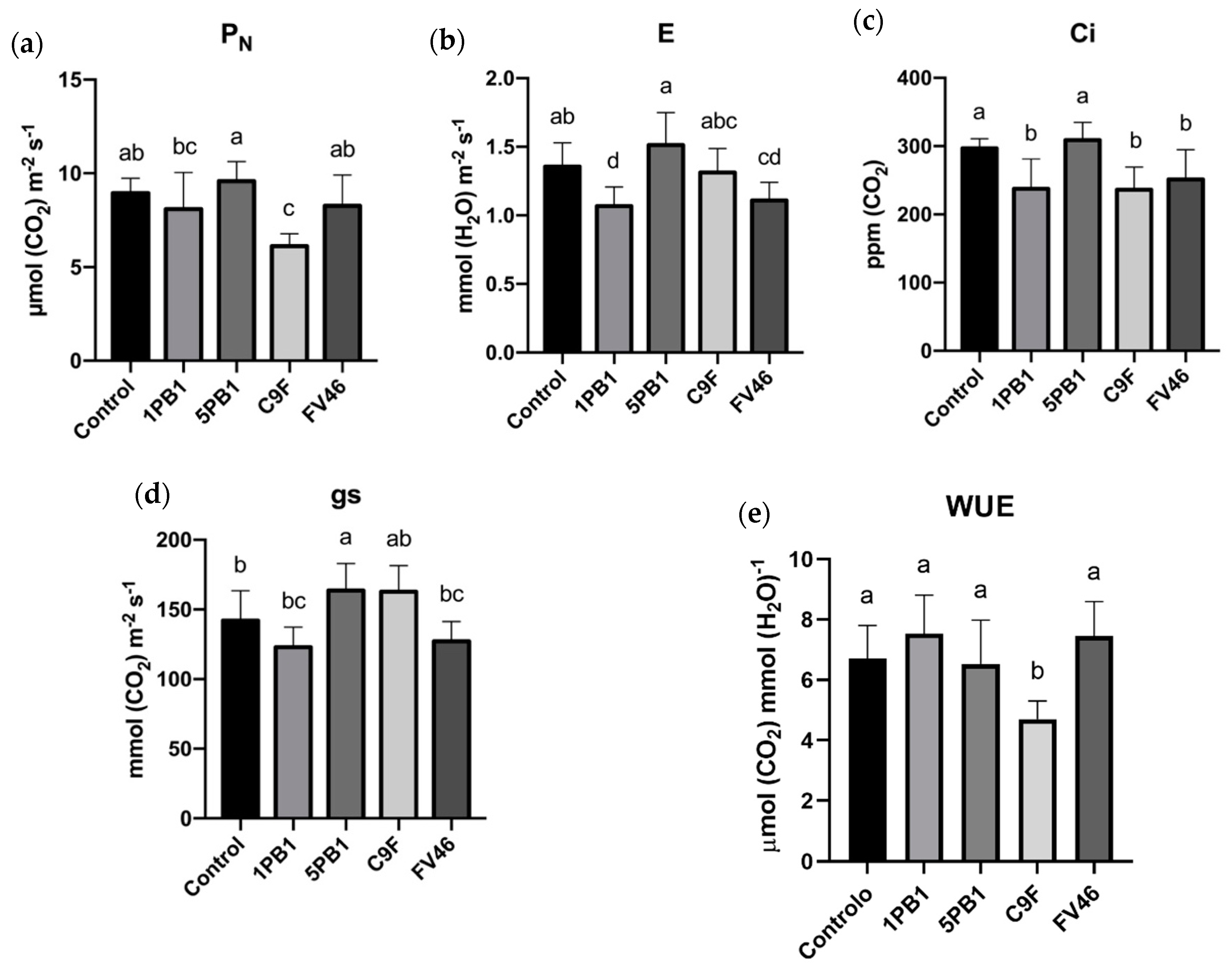
3.4. Photosynthetic Performance
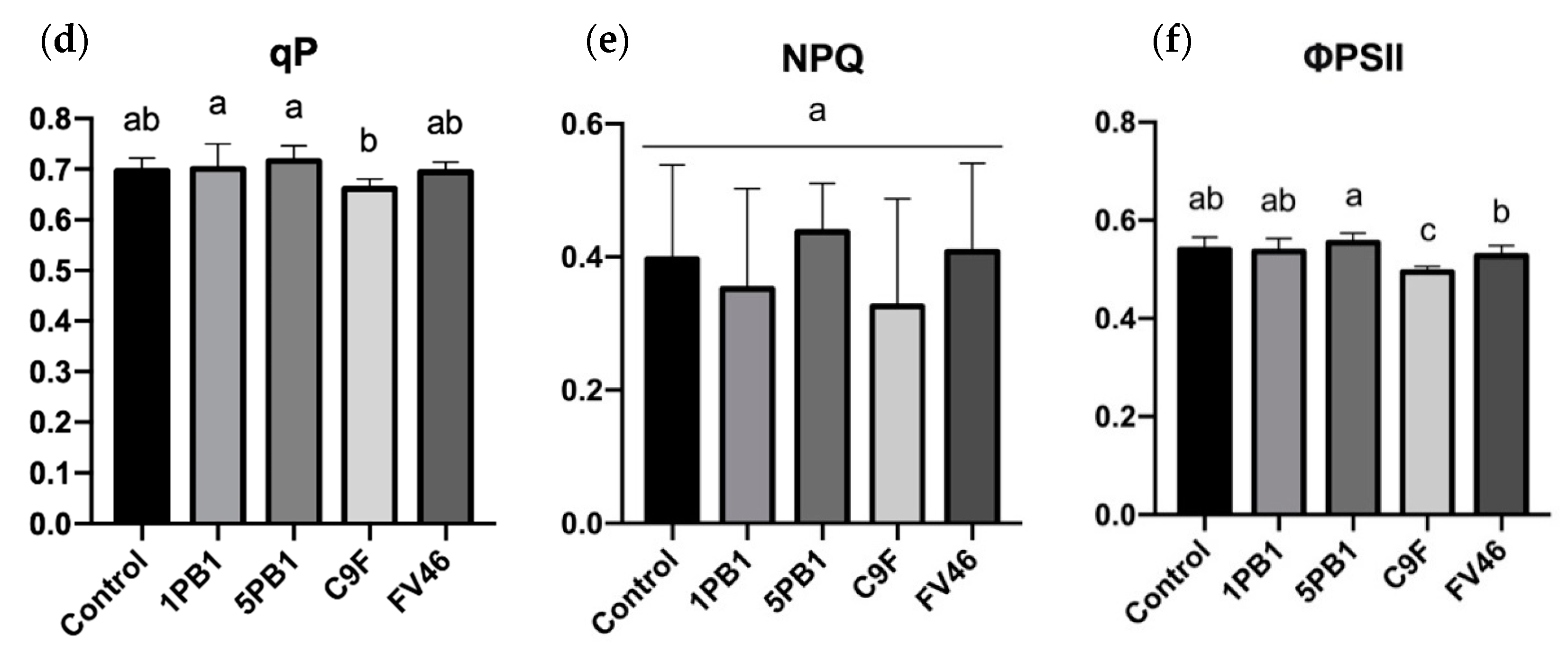
3.5. Gas Exchange
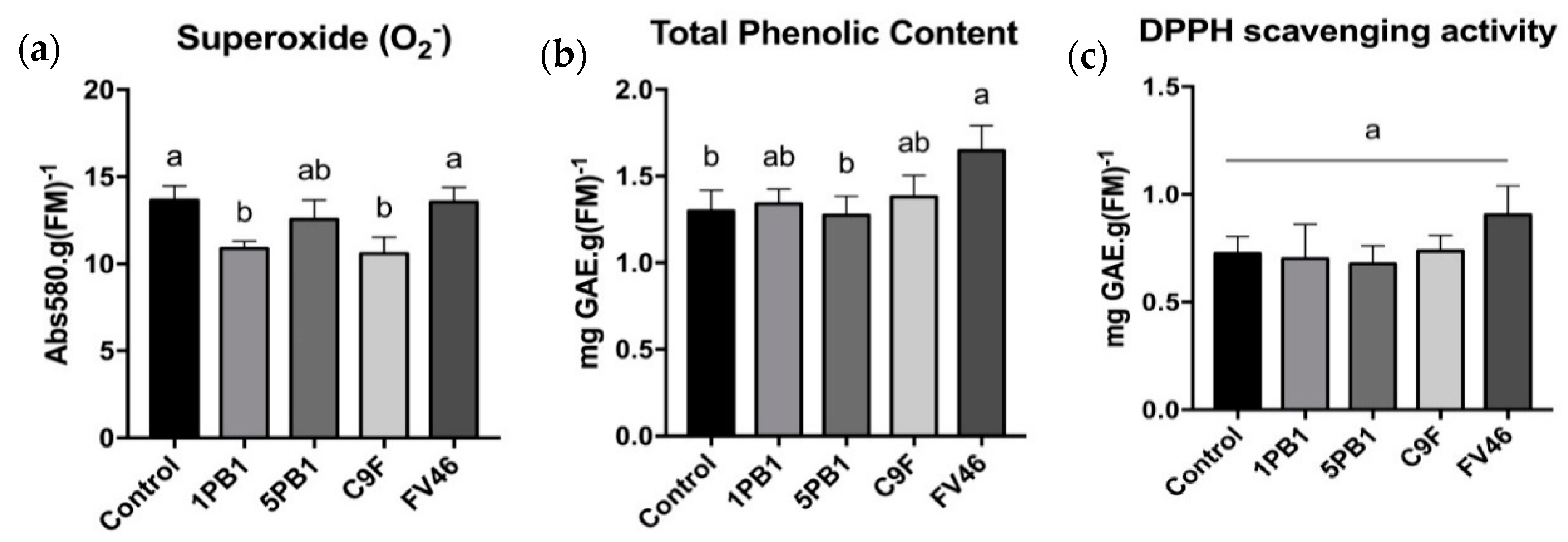
3.6. Superoxide Content and Non-Enzymatic Antioxidant Activity
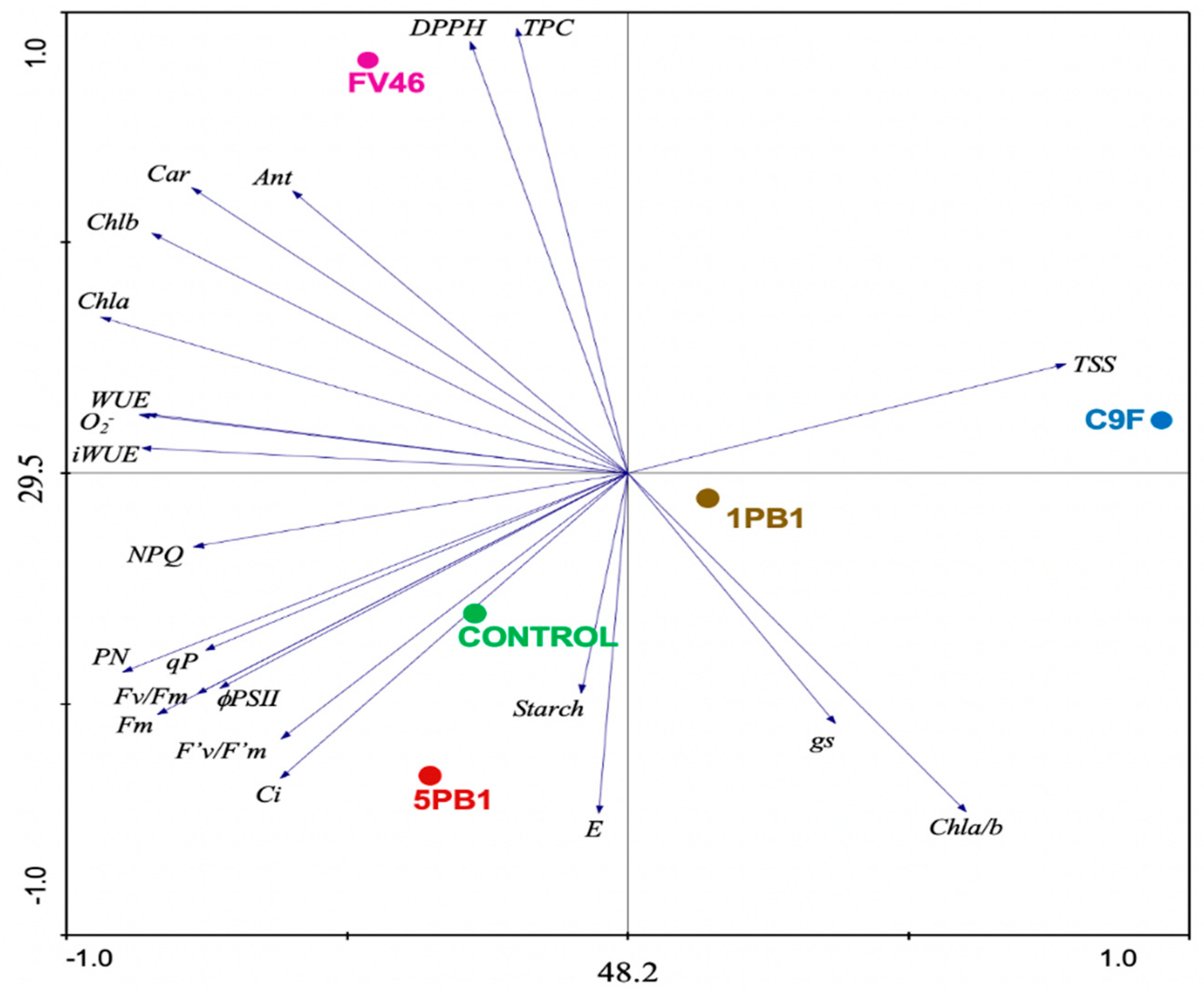
3.7. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brown, K.H.; Jameton, A.L. Public health implications of urban agriculture. J. Public Health Policy 2000, 21, 20–39. [Google Scholar] [CrossRef] [PubMed]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inadequate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Yadav, A.N. (Eds.) Natural Bioactive Products in Sustainable Agriculture; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- Basu, S.; Rabara, R.; Negi, S. Towards a better greener future-an alternative strategy using biofertilizers. I: Plant growth promoting bacteria. Plant Gene 2017, 12, 43–49. [Google Scholar] [CrossRef]
- Chandra, D.; Barh, A.; Sharma, I.P. Plant growth promoting bacteria: A gateway to sustainable agriculture. In Microbial Biotechnology in Environmental Monitoring and Cleanup; IGI Global: Hershey, PA, USA, 2018; pp. 318–338. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.-M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y.; et al. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid (IAA) production trait, a useful screening to select endophytic and rhizosphere competent bacteria for rice growth promoting agents. MethodsX 2015, 2, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Johri, B.N. Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron limiting conditions. Microbiol. Res. 2003, 158, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Gull, M.; Hafeez, F.Y. Characterization of siderophore producing bacterial strain Pseudomonas fluorescens Mst 8.2 as plant growth promoting and biocontrol agent in wheat. Afr. J. Microbiol. Res. 2012, 6, 6308–6318. [Google Scholar] [CrossRef] [Green Version]
- Yadegari, M.; Rahmani, H.A.; Noormohammadi, G.; Ayneband, A. Plant growth promoting rhizobacteria increase growth, yield and nitrogen fixation in Phaseolus vulgaris. J. Plant Nutr. 2010, 33, 1733–1743. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Samineni, S. Nitrogen fixation, plant growth and yield enhancements by diazotrophic growth-promoting bacteria in two cultivars of chickpea (Cicer arietinum L.). Biocatal. Agric. Biotechnol. 2017, 11, 116–123. [Google Scholar] [CrossRef]
- Kang, S.-M.; Joo, G.-J.; Hamayun, M.; Na, C.-I.; Shin, D.-H.; Kim, H.Y.; Hong, J.-K.; Lee, I.-J. Gibberellin production and phosphate solubilization by newly isolated strain of Acinetobacter calcoaceticus and its effect on plant growth. Biotechnol. Lett. 2008, 31, 277–281. [Google Scholar] [CrossRef]
- Midekssa, M.J.; Löscher, C.R.; Schmitz, R.A.; Assefa, F. Phosphate solubilization and multiple plant growth promoting properties of rhizobacteria isolated from chickpea (Cicer aeritinum L.) producing areas of Ethiopia. Afr. J. Biotechnol. 2016, 15, 1899–1912. [Google Scholar] [CrossRef] [Green Version]
- Mnif, I.; Ghribi, D. Potential of bacterial derived biopesticides in pest management. Crop Prot. 2015, 77, 52–64. [Google Scholar] [CrossRef]
- Mnif, I.; Hammami, I.; Triki, M.A.; Azabou, M.C.; Ellouze-Chaabouni, S.; Ghribi, D. Antifungal efficiency of a lipopeptide biosurfactant derived from Bacillus subtilis SPB1 versus the phytopathogenic fungus, Fusarium solani. Environ. Sci. Pollut. Res. 2015, 22, 18137–18147. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Marahiel, M.A. Siderophore-Based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2015, 71, 413–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. Plant growth-promoting microbial-mediated induced systemic resistance in plants: Induction, mechanism, and expression. In Microbial-Mediated Induced Systemic Resistance in Plants; Springer: Singapore, 2016; pp. 213–226. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; García-Estévez, I.; García-Fraile, P.; Escribano-Bailón, M.T.; Rivas, R. Increase in phenolic compounds of Coriandrum sativum L. after the application of a Bacillus halotolerans biofertilizer. J. Sci. Food Agric. 2020, 100, 2742–2749. [Google Scholar] [CrossRef] [PubMed]
- Danish, S.; Zafar-ul-Hye, M. Co-Application of ACC-deaminase producing PGPR and timber-waste biochar improves pigments formation, growth and yield of wheat under drought stress. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Lehvävirta, S.; Timonen, S.; Kasurinen, J.; Niemikapee, J.; Valkonen, J.P.T. Species-specific synergistic effects of two plant growth—promoting microbes on green roof plant biomass and photosynthetic efficiency. PLoS ONE 2018, 13, e0209432. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lou, K.; Li, C. Growth and photosynthetic efficiency promotion of sugar beet (Beta vulgaris L.) by endophytic bacteria. Photosynth. Res. 2010, 105, 5–13. [Google Scholar] [CrossRef]
- Stefan, M.; Munteanu, N.; Stoleru, V.; Mihasan, M.; Hritcu, L. Seed inoculation with plant growth promoting rhizobacteria enhances photosynthesis and yield of runner bean (Phaseolus coccineus L.). Sci. Hortic. 2013, 151, 22–29. [Google Scholar] [CrossRef]
- Wang, C.-J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.-D.; Liu, H.-X.; Wang, Y.; Guo, J.-H. Induction of drought tolerance in cucumber plants by a consortium of three plant growth-promoting Rhizobacterium strains. PLoS ONE 2012, 7, e52565. [Google Scholar] [CrossRef] [Green Version]
- Vimal, S.R.; Patel, V.K.; Singh, J.S. Plant growth promoting Curtobacterium albidum strain SRV4: An agriculturally important microbe to alleviate salinity stress in paddy plants. Ecol. Indic. 2018, 105, 553–562. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R. Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.S.; Lee, K.D. Plant growth promoting rhizobacteria effect on antioxidant status, photosynthesis, mineral uptake and growth of lettuce under soil salinity. Res. J. Agric. Biol. Sci. 2005, 1, 210–215. [Google Scholar]
- Instituto Português da Atmosfera e do Mar. Boletim Climatológico Mensal—Julho de 2020. 2020. Available online: http://www.ipma.pt/pt/media/noticias/documentos/2020/resumo_clima_mar2020.pdf (accessed on 10 October 2020).
- Instituto Português da Atmosfera e do Mar. Resumo Climatológico Março de 2020. 2020. Available online: https://www.ipma.pt/pt/media/noticias/news.detail.jsp?f=/pt/media/noticias/textos/resumo-clima-julho-20.html (accessed on 10 October 2020).
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Osaki, M.; Shinano, T.; Tadano, T. Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci. Plant Nutr. 1991, 37, 117–128. [Google Scholar] [CrossRef]
- Dias, M.C.; Santos, C.; Pinto, G.; Silva, A.M.; Silva, S. Titanium dioxide nanoparticles impaired both photochemical and non-photochemical phases of photosynthesis in wheat. Protoplasma 2019, 256, 69–78. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Harkat-Madouri, L.; Asma, B.; Madani, K.; Said, Z.B.-O.S.; Rigou, P.; Grenier, D.; Allaloua, H.; Remini, H.; Adjaouda, A.; Boulekbache-Makhlouf, L. Chemical composition, antibacterial and antioxidant activities of essential oil of Eucalyptus globulus from Algeria. Ind. Crop. Prod. 2015, 78, 148–153. [Google Scholar] [CrossRef]
- Gajewska, E.; Sklodowska, M. Effect of nickel on ROS content and antioxidative enzyme activities in wheat leaves. BioMetals 2007, 20, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: Feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.K.; McCouch, S. Getting to the roots of it: Genetic and hormonal control of root architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef] [Green Version]
- Cabra Cendales, T.; Rodríguez González, C.A.; Villota Cuásquer, C.P.; Tapasco Alzate, O.A.; Hernández Rodríguez, A. Bacillus effect on the germination and growth of tomato seedlings (Solanum lycopersicum L.). Acta Biol. Colomb. 2017, 22, 37. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Duraivadivel, P.; Hariprasad, P.; Niranjana, S.R. A novel split-pot bioassay to screen indole acetic acid producing rhizobacteria for the improvement of plant growth in tomato (Solanum lycopersicum L.). Sci. Hortic. 2017, 224, 351–357. [Google Scholar] [CrossRef]
- Zhori, A.; Meco, M.; Brandl, H.; Bachofen, R. In situ chlorophyll fluorescence kinetics as a tool to quantify effects on photosynthesis in Euphorbia cyparissias by a parasitic infection of the rust fungus Uromyces pisi. BMC Res. Notes 2015, 8, 698. [Google Scholar] [CrossRef] [Green Version]
- Yobo, K.S.; Laing, M.D.; Hunter, C.H. Effects of single and combined inoculations of selected Trichoderma and Bacillus isolates on growth of dry bean and biological control of Rhizoctonia solani damping-off. Afr. J. Biotechnol. 2011, 10, 8746–8756. [Google Scholar]
- Anusaraporn, S.; Autarmat, S.; Treesubsuntorn, C.; Thiravetyan, P. Application Bacillus sp. N7 to enhance ozone tolerance of various Oryza sativa in vegetative phase: Possible mechanism and rice productivity. Biocatal. Agric. Biotechnol. 2020, 25, 101591. [Google Scholar] [CrossRef]
- de Castro, G.L.S.; Rêgo, M.C.F.; Silvestre, W.V.D.; Batista, T.F.V.; da Silva, G.B. Açaí palm seedling growth promotion by rhizobacteria inoculation. Braz. J. Microbiol. 2020, 51, 205–216. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Gao, J.; Pan, S.; Wang, G. Bacillus subtilis-regulation of stomatal movement and instantaneous water use efficiency in Vicia faba. Plant Growth Regul. 2015, 78, 43–55. [Google Scholar] [CrossRef]
- Jain, L.; Jajoo, A. Protection of PSI and PSII complexes of wheat from toxic effect of anthracene by Bacillus subtilis (NCIM 5594). Photosynth. Res. 2020, 146, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Vaikuntapu, P.R.; Dutta, S.; Samudrala, R.B.; Rao, V.R.V.N.; Kalam, S.; Podile, A.R. Preferential promotion of Lycopersicon esculentum (Tomato) Growth by plant growth promoting bacteria associated with tomato. Indian J. Microbiol. 2014, 54, 403–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaniego-Gámez, B.Y.; Garruña, R.; Tun-Suárez, J.M.; Kantun-Can, J.; Reyes-Ramírez, A.; Cervantes-Díaz, L. Bacillus spp. inoculation improves photosystem II efficiency and enhances photosynthesis in pepper plants. Chil. J. Agric. Res. 2016, 76, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Akram, W.; Aslam, H.; Ahmad, S.R.; Anjum, T.; Yasin, N.A.; Khan, W.U.; Ahmad, A.; Guo, J.; Wu, T.; Luo, W.; et al. Bacillus megaterium strain A12 ameliorates salinity stress in tomato plants through multiple mechanisms. J. Plant Interact. 2019, 14, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.; Amaresan, N.; Patel, P.; Jinal, H.N.; Krishnamurthy, R. Isolation and characterization of Bacillus spp. endowed with multifarious plant growth-promoting traits and their potential effect on tomato (Lycopersicon esculentum) seedlings. Arab. J. Sci. Eng. 2020, 45, 4579–4587. [Google Scholar] [CrossRef]
- Morais, M.C.; Mucha, Â.; Ferreira, H.; Gonçalves, B.; Bacelar, E.; Marques, G. Comparative study of plant growth-promoting bacteria on the physiology, growth and fruit quality of strawberry. J. Sci. Food Agric. 2019, 99, 5341–5349. [Google Scholar] [CrossRef] [PubMed]
- Gagné-Bourque, F.; Mayer, B.F.; Charron, J.-B.; Vali, H.; Bertrand, A.; Jabaji, S. Accelerated growth rate and increased drought stress resilience of the model grass Brachypodium distachyon Colonized by Bacillus subtilis B26. PLoS ONE 2015, 10, e0130456. [Google Scholar] [CrossRef]
- Li, J.; Guan, Y.; Yuan, L.; Hou, J.; Wang, C.; Liu, F.; Yang, Y.; Lu, Z.; Chen, G.; Zhu, S. Effects of exogenous IAA in regulating photosynthetic capacity, carbohydrate metabolism and yield of Zizania latifolia. Sci. Hortic. 2019, 253, 276–285. [Google Scholar] [CrossRef]
- Liu, P.; Ewis, H.E.; Huang, Y.J.; Lu, C.D.; Tai, P.C.; Weber, I.T. Structure of Bacillus subtilis superoxide dismutase. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2007, 63, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, I.; Egamberdieva, D. Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Prakash, D.; Singh, B.N.; Upadhyay, G. Antioxidant and free radical scavenging activities of phenols from onion (Allium cepa). Food Chem. 2007, 102, 1389–1393. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 1PB1 | 5PB1 | C9F | FV46 | |
|---|---|---|---|---|---|
| Shoot length | 3.237 ± 0.491 a | 3.147 ± 0.527 a | 3.082 ± 0.513 a | 3.100 ± 0.489 a | 3.124 ± 0.491 a |
| Shoot FW | 0.0609 ± 0.0139 a | 0.0558 ± 0.0007 a | 0.0561 ± 0.0097 a | 0.0572 ± 0.0054 a | 0.0568 ± 0.0118 a |
| Shoot DW | 0.00256 ± 0.00053 a | 0.00252 ± 0.00011 a | 0.00251 ± 0.00045 a | 0.00253 ± 0.00047 a | 0.00257 ± 0.00056 a |
| Root length | 9.163 ± 1.761 a | 6.211 ± 1.195 c | 7.382 ± 1.261 bc | 8.271 ± 2.074 ab | 8.082 ± 1.694 ab |
| Root FW | 0.012 ± 0.005 a | 0.012 ± 0.006 a | 0.010 ± 0.006 a | 0.013 ± 0.003 a | 0.010 ± 0.004 a |
| Root DW | 0.00053 ± 0.00005 a | 0.00054 ± 0.00011 a | 0.00049 ± 0.00012 a | 0.00063 ± 0.00018 a | 0.00056 ± 0.00002 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Santos, M.; Mariz-Ponte, N.; Dias, M.C.; Moura, L.; Marques, G.; Santos, C. Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum. Horticulturae 2021, 7, 24. https://doi.org/10.3390/horticulturae7020024
Costa-Santos M, Mariz-Ponte N, Dias MC, Moura L, Marques G, Santos C. Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum. Horticulturae. 2021; 7(2):24. https://doi.org/10.3390/horticulturae7020024
Chicago/Turabian StyleCosta-Santos, Marino, Nuno Mariz-Ponte, Maria Celeste Dias, Luísa Moura, Guilhermina Marques, and Conceição Santos. 2021. "Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum" Horticulturae 7, no. 2: 24. https://doi.org/10.3390/horticulturae7020024
APA StyleCosta-Santos, M., Mariz-Ponte, N., Dias, M. C., Moura, L., Marques, G., & Santos, C. (2021). Effect of Bacillus spp. and Brevibacillus sp. on the Photosynthesis and Redox Status of Solanum lycopersicum. Horticulturae, 7(2), 24. https://doi.org/10.3390/horticulturae7020024