
Evaluating the Drought Endurance of Landscaping Ground Cover Plants in a Roof Top Model



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Environmental Conditions
2.3. Plant Longevity
2.4. Physiological Responses
2.5. Plant Health
2.6. Color Appearance
2.7. Data Analysis
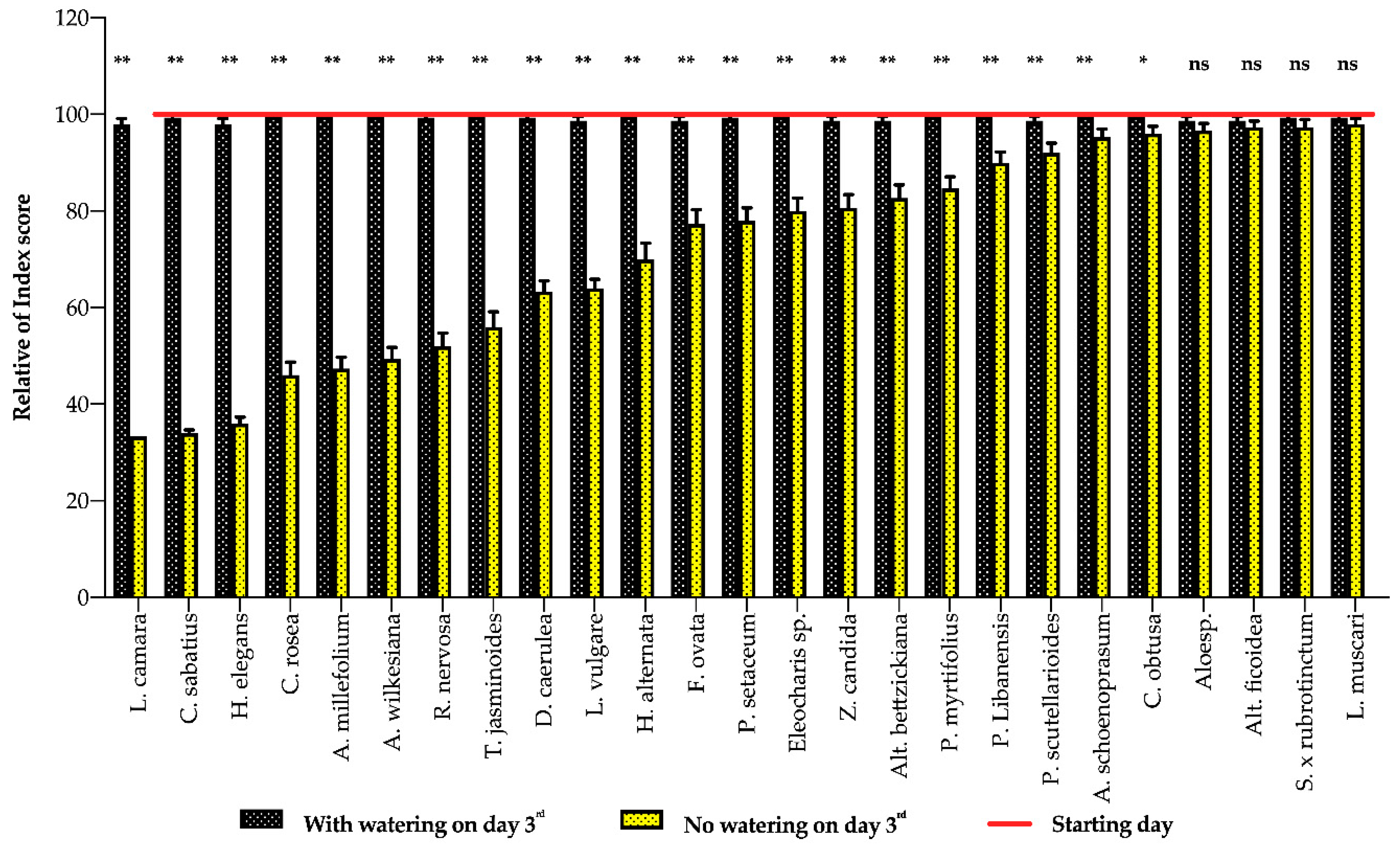
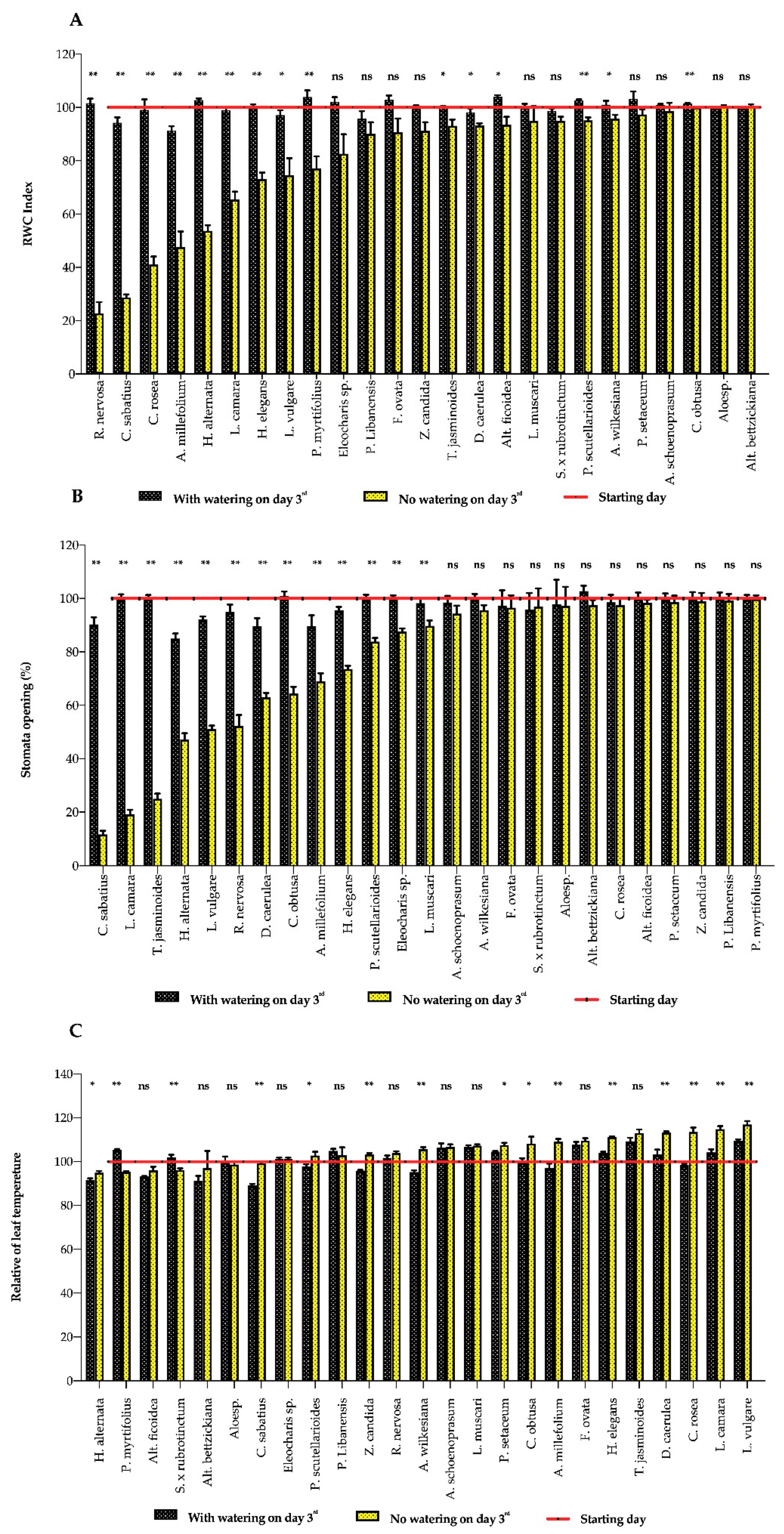
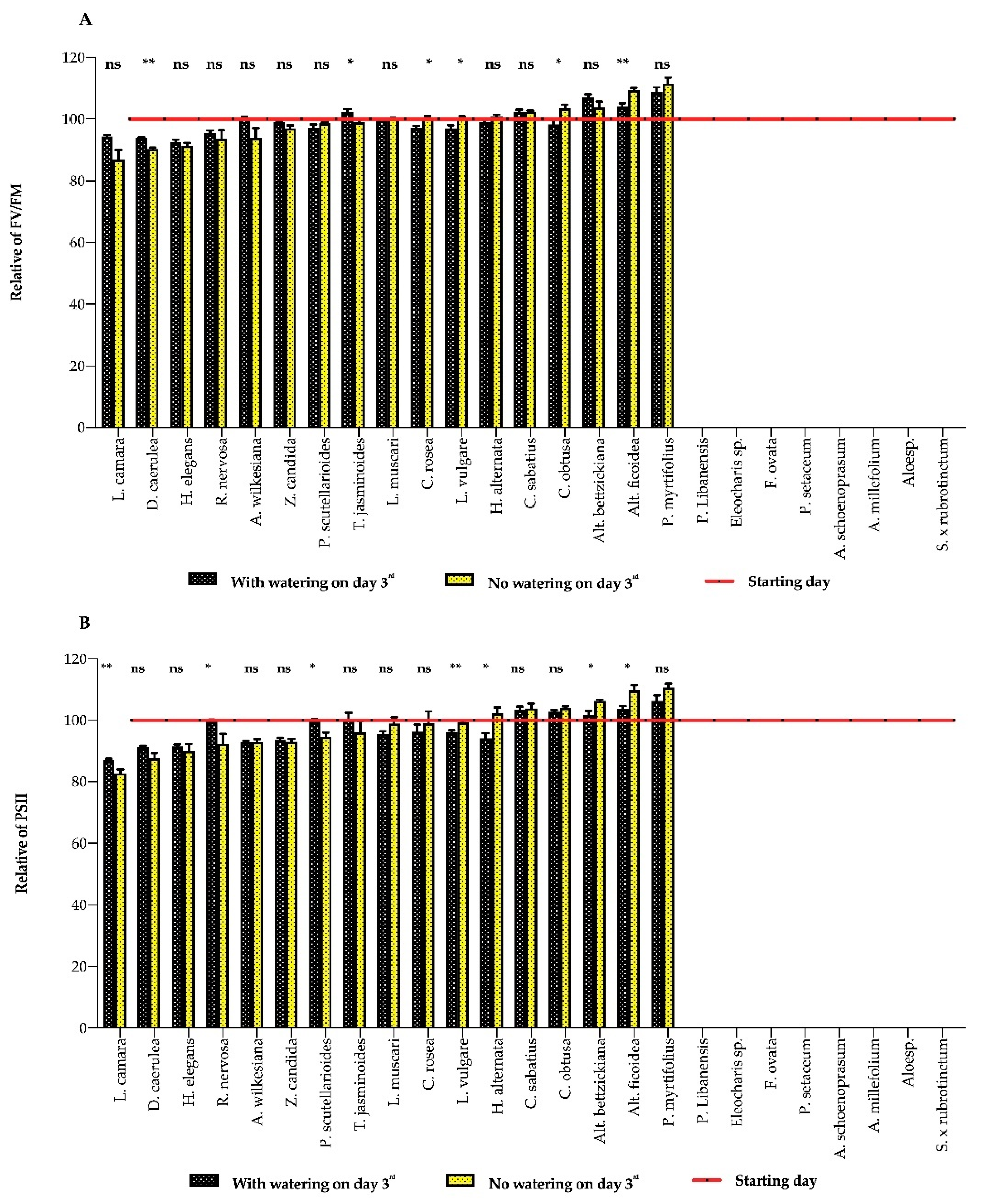
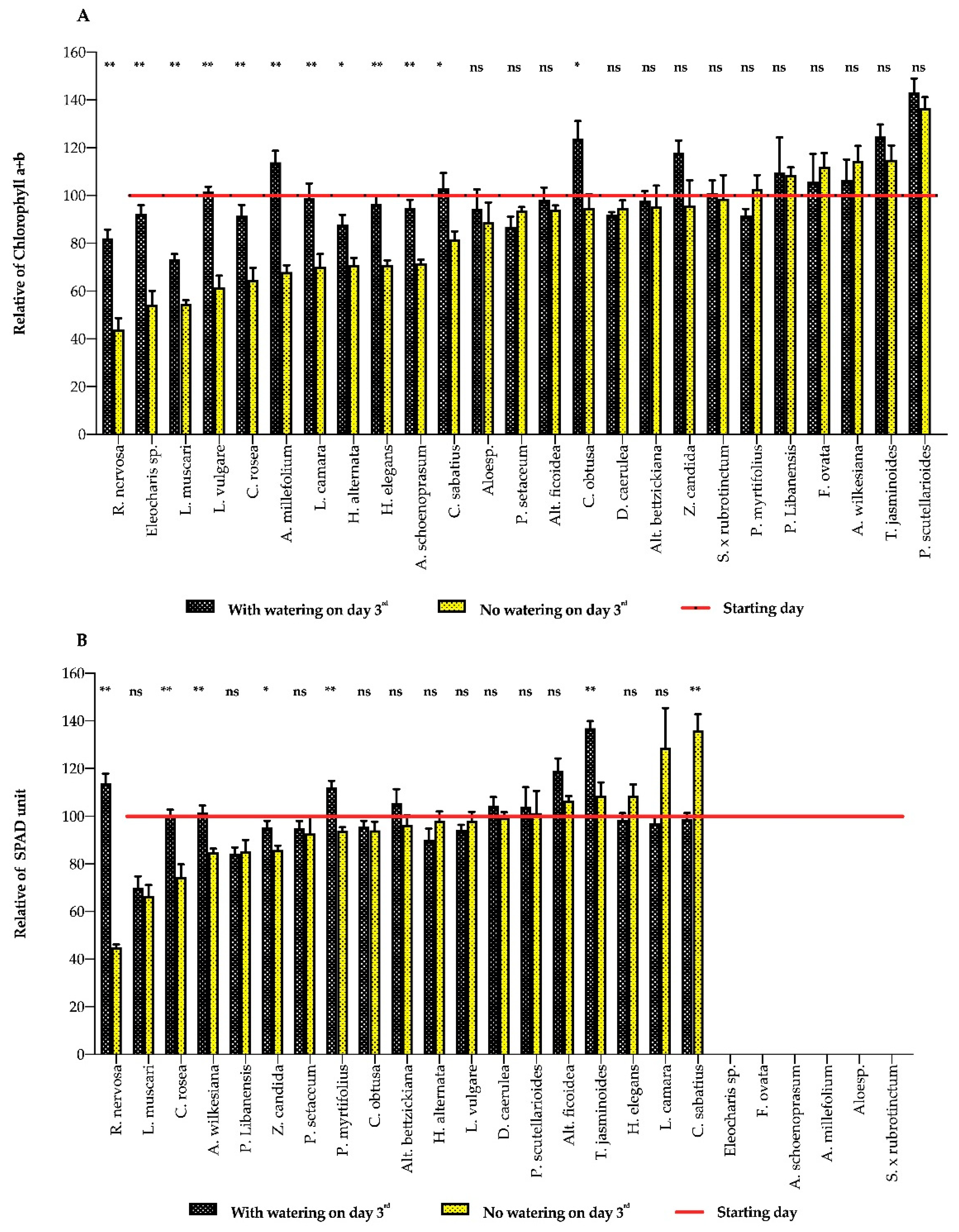
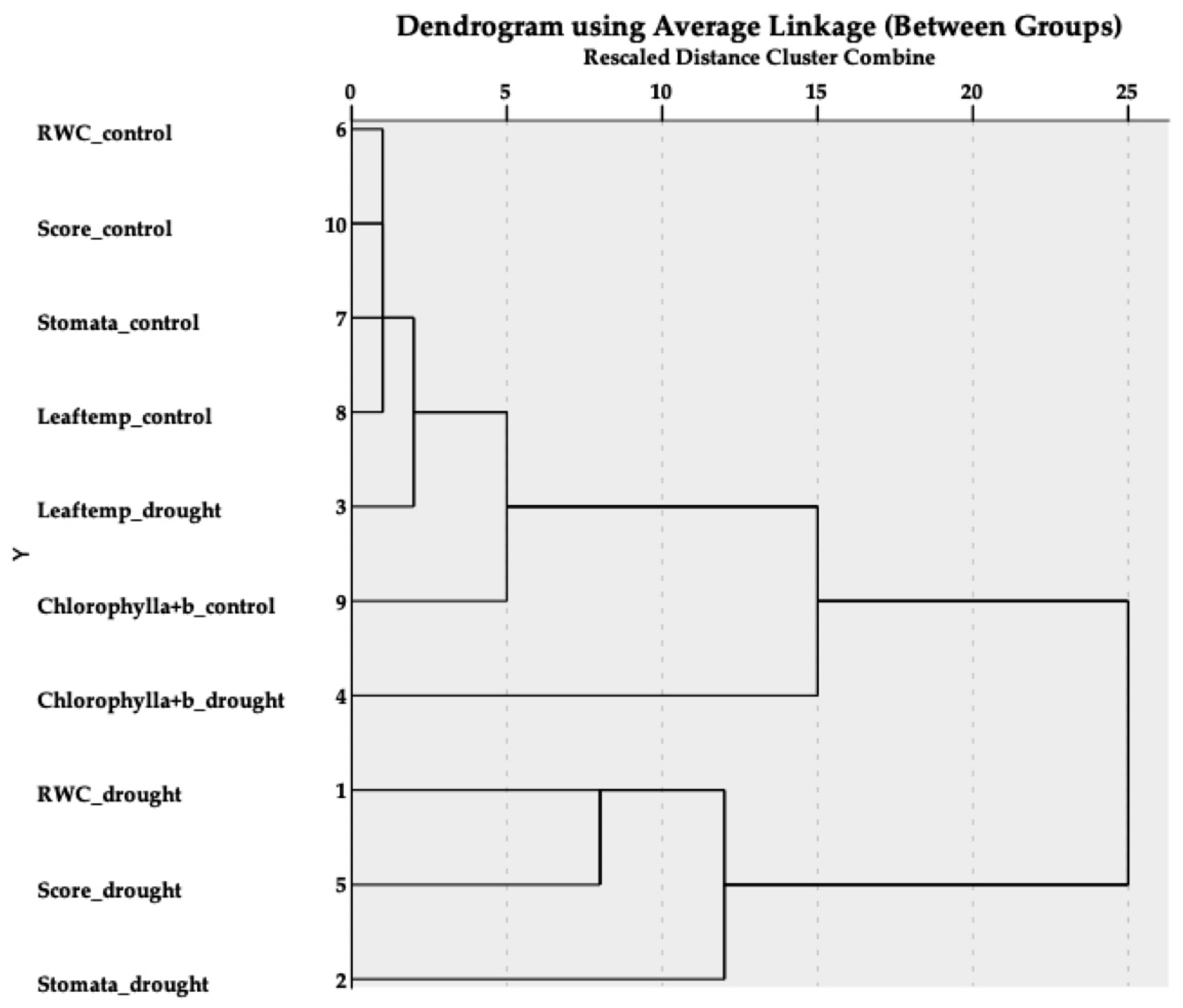
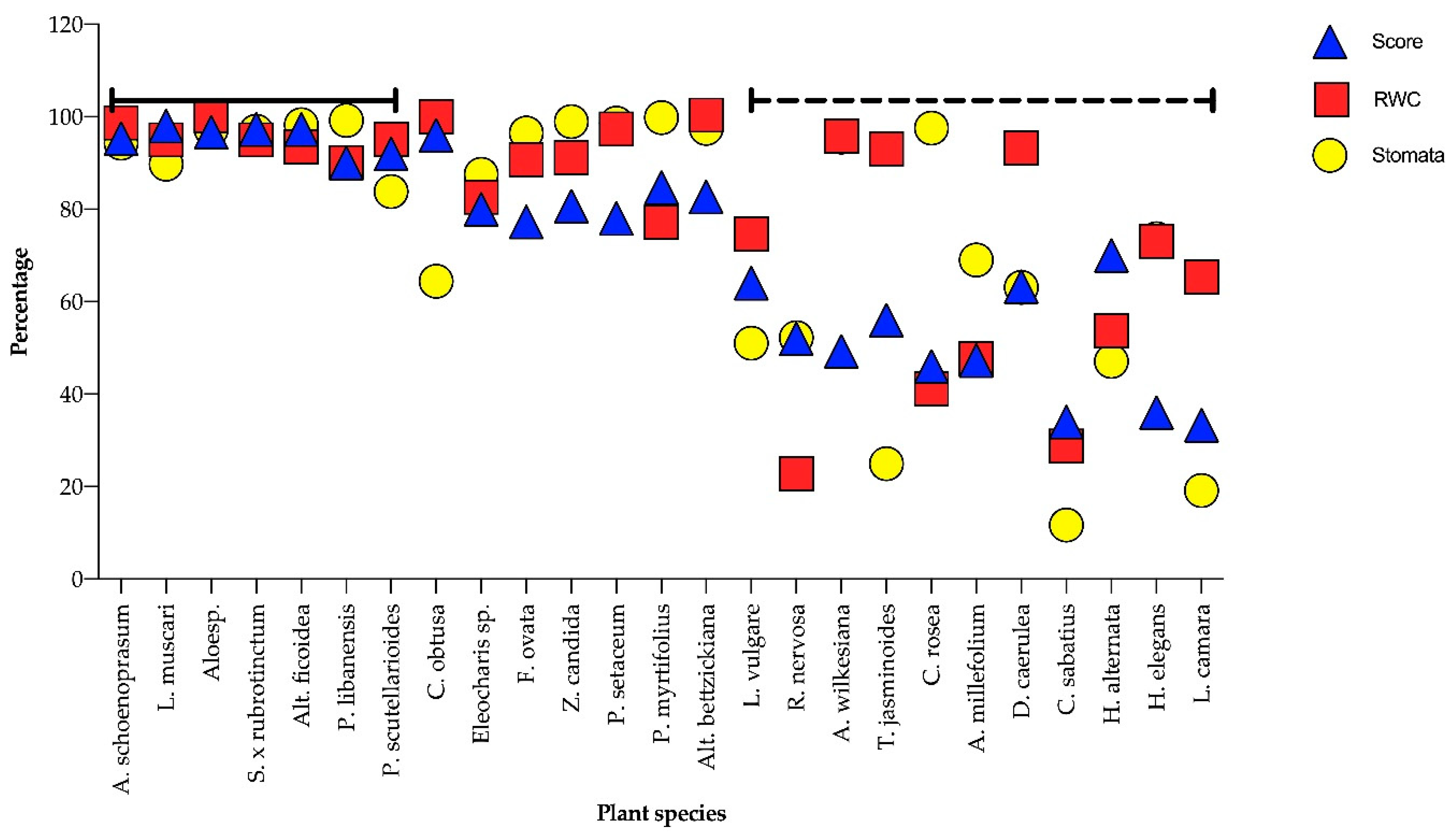
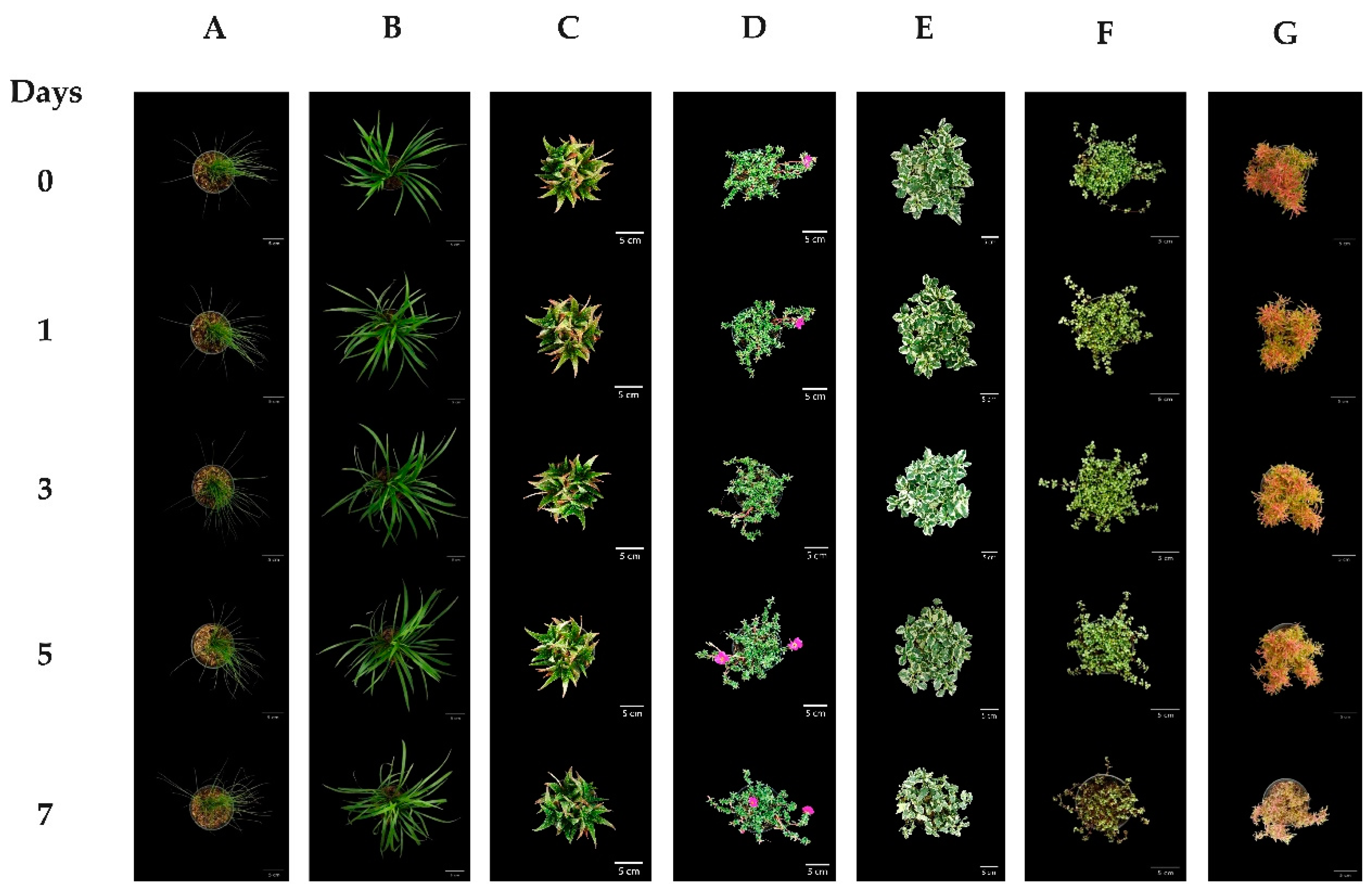
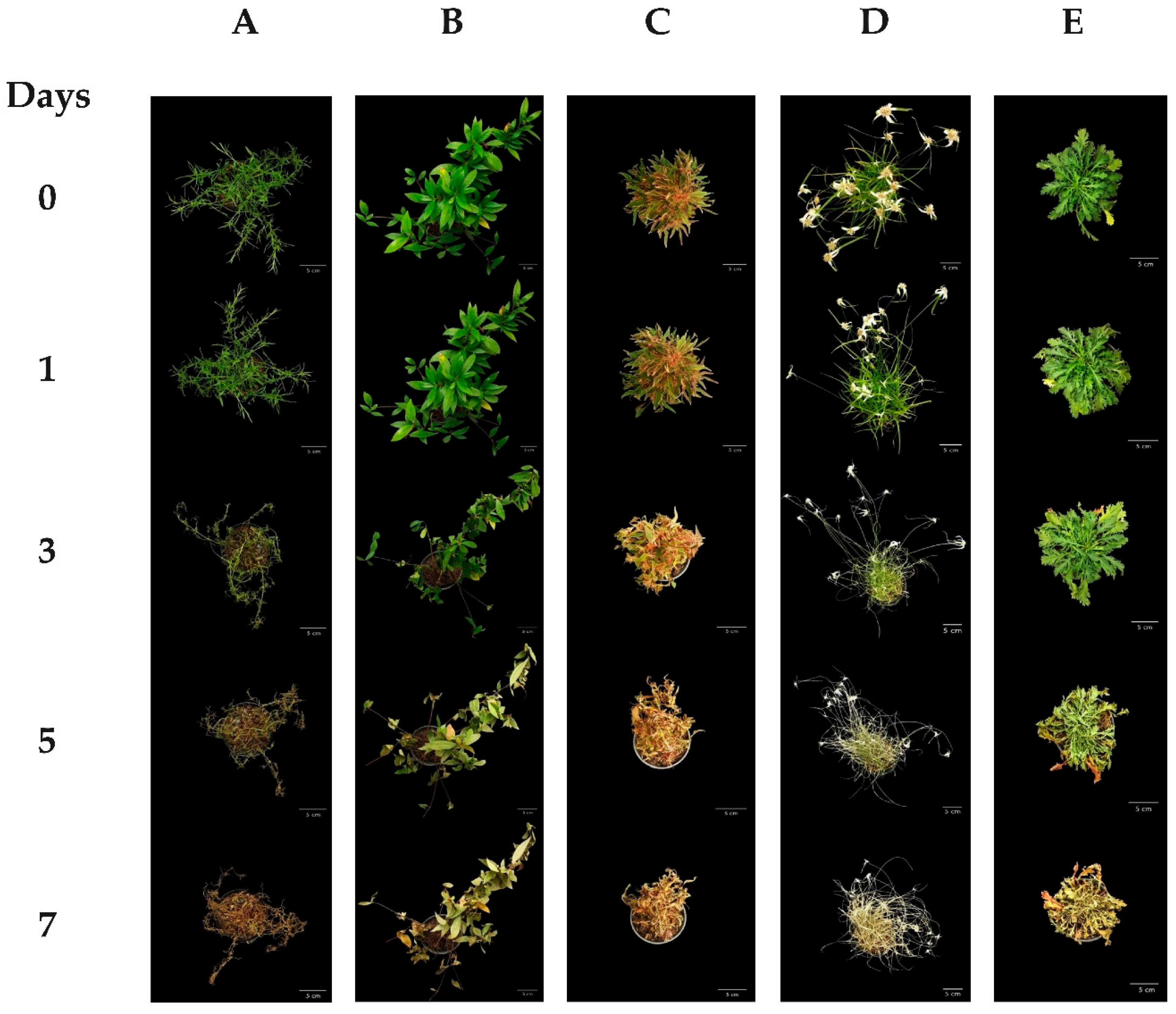
3. Results
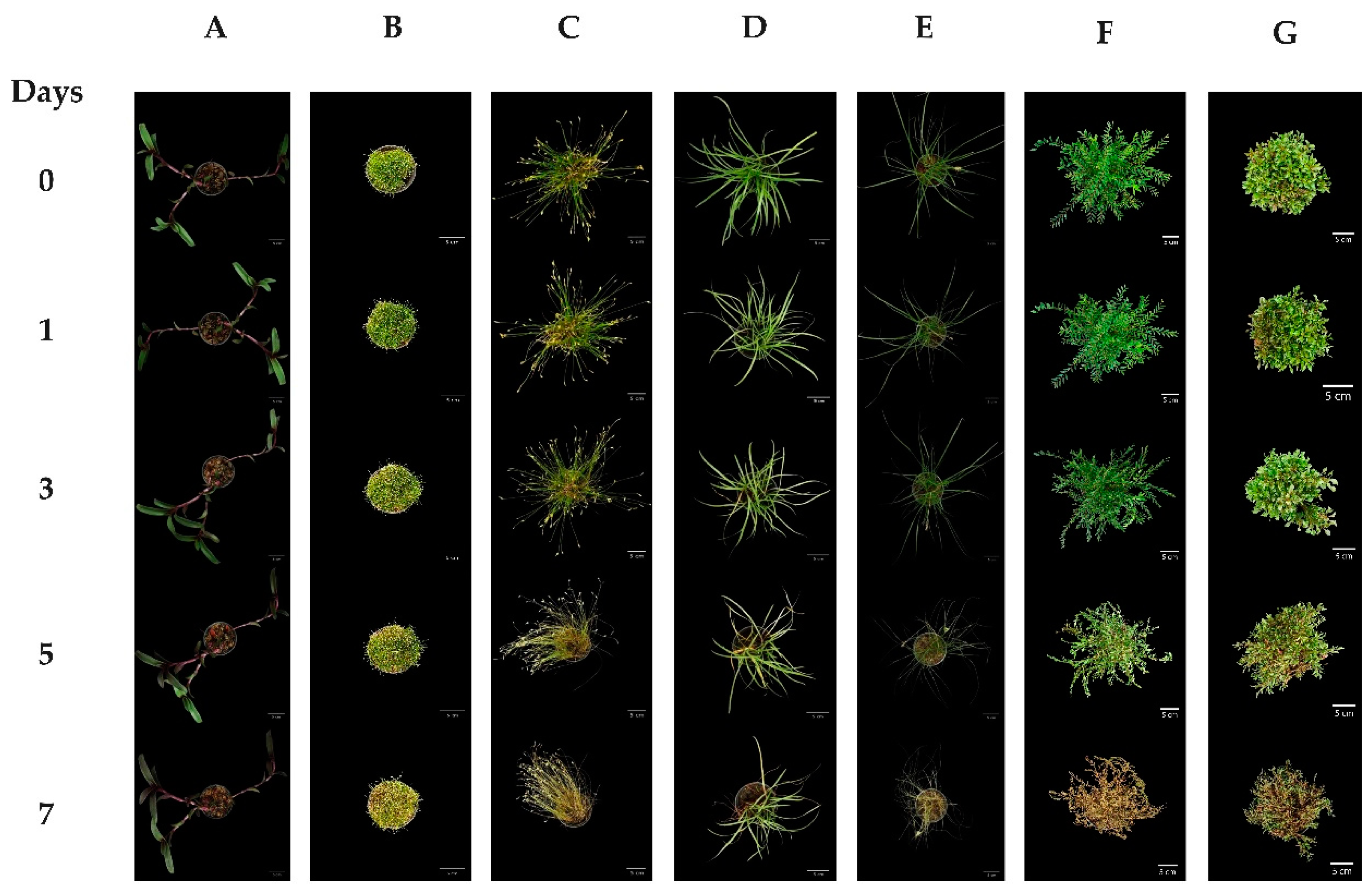
3.1. Plant Response to Water Deficits
3.2. Indicator Implements
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Scientific Name | Days after the Start of the Drought | |||
|---|---|---|---|---|---|
| 1 | 3 | 5 | 7 | ||
| 1 | Hemigraphis alternata (Burm.f.) T. Anderson (Syn.) (Strobilanthes alternata (Burm.f.) Moylan ex J.R.I. Wood) | 2.86 * | 1.86 * | 1.38 * | 1.00 * |
| 2 | Alternanthera bettzickiana (Regel) G. Nicholson | 2.78 * | 2.48 * | 1.56 * | 1.00 * |
| 3 | Alternanthera ficoidea (L.) P. Beauv. | 2.98 | 2.92 | 2.62 * | 1.50 * |
| 4 | Allium schoenoprasum L. | 2.94 | 2.86 * | 2.70 * | 2.12 * |
| 5 | Zephyranthes candida (Lindl.) Herb. | 2.76 * | 2.42 * | 1.54 * | 1.36 * |
| 6 | Trachelospermum jasminoides (Lindl.) Lem. | 2.96 | 1.68 * | 1.08 * | 1.02 * |
| 7 | Liriope muscari (Decne.) LH. Bailey | 3.00 | 2.94 | 2.86 | 2.56 * |
| 8 | Aloe sp. | 2.94 | 2.90 | 2.90 | 2.90 |
| 9 | Dianella caerulea Sims | 2.88 * | 1.90 * | 1.42 * | 1.14 * |
| 10 | Achillea millefolium L. | 2.78 * | 1.42 * | 1.02 * | 1.00 * |
| 11 | Coreopsis rosea Nutt. | 2.70 * | 1.38 * | 1.02 * | 1.00 * |
| 12 | Leucanthemum vulgare (Vaill.) Lam. | 2.72 * | 1.92 * | 1.12 * | 1.00 * |
| 13 | Cyanotis obtusa (Trimen) Trimen (Syn.) | 2.94 | 2.88 * | 2.74 * | 2.48 * |
| 14 | Convolvulus sabatius Viv. | 2.16 * | 1.00 * | 1.00 * | 1.00 * |
| 15 | Sedum x rubrotinctum R.T. Clausen | 2.96 | 2.94 | 2.94 | 2.94 |
| 16 | Fimbristylis ovata (Burm.f.) J.Kern | 2.78 * | 2.32 * | 1.30 * | 1.10 * |
| 17 | Rhynchospora nervosa (Vahl) Boeckeler | 2.48 * | 1.56 * | 1.06 * | 1.00 * |
| 18 | Eleocharis sp. | 2.90 * | 2.40 * | 1.84 * | 1.14 * |
| 19 | Acalypha wilkesiana Müll.Arg. | 2.84 * | 1.48 * | 1.00 * | 1.00 * |
| 20 | Plectranthus scutellarioides (L.) R.Br. | 2.90 * | 2.76 * | 2.22 * | 1.04 * |
| 21 | Heterocentron elegans Kuntze | 1.44 * | 1.08 * | 1.00 * | 1.00 * |
| 22 | Phyllanthus myrtifolius (Wight) Müll.Arg. | 2.98 | 2.54 * | 1.42 * | 1.00 * |
| 23 | Pennisetum setaceum (Forssk.) Chiov. (Syn.) (Cenchrus setaceus (Forssk.) Morrone.) | 2.64 * | 2.34 * | 1.30 * | 1.02 * |
| 24 | Pilea libanensis Urb. | 2.88 * | 2.70 * | 2.36 * | 1.04 * |
| 25 | Lantana camara L. | 1.52 * | 1.00 * | 1.00 * | 1.00 * |




References
- Jongtanom, Y.; Kositanont, C.; Baulert, S. Temporal variations of urban heat island intensity in three major cities, Thailand. Mod. Appl. Sci. 2011, 5, 105. [Google Scholar] [CrossRef] [Green Version]
- Getter, K.L.; Rowe, D.B. The Role of Extensive Green Roofs in Sustainable Development. HortScience 2006, 41, 1276. [Google Scholar] [CrossRef] [Green Version]
- Fathi, M.; Masnavi, M. Assessing environmental aesthetics of roadside vegetation and scenic beauty of highway landscape: Preferences and perception of motorists. Int. J. Environ. Res. 2014, 8, 941–952. [Google Scholar]
- Klett, J.E.; Wilson, C.R. Xeriscaping: Ground Cover Plants. Ph.D. Thesis, Colorado State University Extension, Fort Collin, CO, USA, 2007. [Google Scholar]
- Oraee, A.; Tehranifar, A. Evaluating the potential drought tolerance of pansy through its physiological and biochemical responses to drought and recovery periods. Sci. Hortic. 2020, 265, 109225. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dichio, B.; Xiloyannis, C.; Sofo, A.; Montanaro, G. Osmotic regulation in leaves and roots of olive trees during a water deficit and rewatering. Tree Physiol. 2006, 26, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juan, M.; Rivero, R.M.; Romero, L.; Ruiz, J.M. Evaluation of some nutritional and biochemical indicators in selecting salt-resistant tomato cultivars. Environ. Exp. Bot. 2005, 54, 193–201. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Hu, Y.; Yakout, G.M.; Awad, A.M.; Hafiz, S.E.; Schmidhalter, U. Evaluating salt tolerance of wheat genotypes using multiple parameters. Eur. J. Agron. 2005, 22, 243–253. [Google Scholar] [CrossRef]
- Getter, K.L.; Rowe, D.B. Selecting Plants for Extensive Green Roofs in the United States; Michigan State University Extension: E. Lansing, MI, USA, 2008. [Google Scholar]
- Ornamental Plant, Landscape, Exotic Plant, Rare Plant. Available online: http://docnursery.com/home.html (accessed on 17 January 2021).
- Hemigraphis Alternata. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?taxonid=275316 (accessed on 21 October 2020).
- Alternanthera Ficoidea. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?taxonid=275668 (accessed on 21 October 2020).
- Allium Schoenoprasum. Available online: https://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=j270 (accessed on 21 October 2020).
- Zephyranthes Candida. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=c747 (accessed on 21 October 2020).
- Trachelospermum Jasminoides. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=a155 (accessed on 21 October 2020).
- Liriope Muscari. Available online: https://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=l100 (accessed on 21 October 2020).
- Aloe Vera. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=b628 (accessed on 21 October 2020).
- Dianella Caerulea—Blue Flax Lily. Available online: https://www.gardeningwithangus.com.au/dianella-caerulea/ (accessed on 22 October 2020).
- Achillea Millefolium. Available online: https://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=b282 (accessed on 21 October 2020).
- Coreopsis rosea. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=s150 (accessed on 21 October 2020).
- Leucanthemum vulgare ‘Maikonigin’ MAY QUEEN. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?taxonid=250182&isprofile=0& (accessed on 21 October 2020).
- Cyanotis Obtusa Trimen. Available online: https://www.nparks.gov.sg/florafaunaweb/flora/6/9/6924 (accessed on 22 October 2020).
- Convolvulus Sabatius. Available online: https://www.perennials.com/plants/convolvulus-sabatius.html (accessed on 22 October 2020).
- Sedum x Rubrotinctum. Available online: https://www.rhs.org.uk/Plants/16987/i-Sedum-i-times;-i-rubrotinctum-i/Details (accessed on 22 October 2020).
- Eleocharis sp. (montevidensis)—Tropica Aquarium Plants. Available online: https://www.birstall.co.uk/products/tropica-eleocharis-sp-montevidensis-.html (accessed on 22 October 2020).
- Acalypha Wilkesiana. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=a478 (accessed on 21 October 2020).
- Plectranthus Scutellarioides. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=a547 (accessed on 21 October 2020).
- Heterocentron Elegans. Available online: http://www.surinenglish.com/lifestyle/201801/05/heterocentron-elegans-20180105105108-v.html (accessed on 22 October 2020).
- Pennisetum Setaceum ‘Rubrum’. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=c257 (accessed on 21 October 2020).
- Lantana Camara. Available online: https://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=a523 (accessed on 21 October 2020).
- Meetam, M.; Sripintusorn, N.; Songnuan, W.; Siriwattanakul, U.; Pichakum, A. Assessment of physiological parameters to determine drought tolerance of plants for extensive green roof architecture in tropical areas. Urban For Urban Green 2020, 56, 126874. [Google Scholar] [CrossRef]
- Turner, N.C. Techniques and experimental approaches for the measurement of plant water status. Plant Soil 1981, 58, 339–366. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric analysis of chlorophylls and carotenoids from commonly grown fern species by using various extracting solvents. Res. J. Chem. Sci. 2014, 4, 63–69. [Google Scholar]
- Dolferus, R. To grow or not to grow: A stressful decision for plants. Plant Sci 2014, 229, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Duca, D.; Glick, B.R. Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 2017, 410, 335–356. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, X.; Ma, Y.; Qing, Y.; Wang, H.; Huang, X. The highly drought-tolerant pitaya (Hylocereus undatus) is a non-facultative CAM plant under both well-watered and drought conditions. J. Hortic. Sci. Biotechnol. 2019, 94, 643–652. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha-um, S.; Boriboonkaset, T.; Pichakum, A.; Kirdmanee, C. Multivariate physiological indices for salt tolerance classification in indica rice (Oryza sativa L. spp. indica). Gen. Appl. Plant Physiol. 2009, 35, 75–87. [Google Scholar]
- Boonanunt, S.; Krisanapook, K.; Boonprakob, U.; Pichakum, A.; Phavaphutanon, L. Suitable criteria for drought-tolerant peach rootstocks grown in northern Thailand. Maejo Int. J. Sci. 2014, 8, 190–197. [Google Scholar] [CrossRef]
- Broussard, M.C. A Horticultural Study of Liriope and Ophiopogon: Nomenclature, Morphology and Culture; Lousiana State University and Agricultural and Mechanical College: Baton Rouge, LA, USA, 2007. [Google Scholar]
- Domenghini, J.C.; Bremer, D.J.; Fry, J.D.; Davis, G.L. Prolonged Drought and Recovery Responses of Kentucky Bluegrass and Ornamental Groundcovers. HortScience 2013, 48, 1209. [Google Scholar] [CrossRef]
- Greeshma, G.M.; Murugan, K. Desiccation tolerance in Artillery Plant (Pilea microphylla (L.) Liebm): A search. Int. Res. J. Environ. Sci. 2015, 4, 26–32. [Google Scholar]
- Tan, P.Y.; Sia, A. A pilot green roof research project in Singapore. In Proceedings of the Third Annual Greening Rooftops for Sustainable Communities Conference, Awards and Trade Show, Washington, DC, USA, 4 May 2005. [Google Scholar]
- Wadugodapitiya, A.; Weeratunga, V.; de Goonatilake, S.A.; Chandranimal, D.; Perera, N.; Asela, C. Insights into the Biodiversity of the Sampur Area in Trincomalee; IUCN, International Union for Conservation of Nature: Columbo, Sri Lanka, 2013; p. 15. [Google Scholar]







| No | Scientific Name | Landscaping Features/Reference |
|---|---|---|
| 1 | Hemigraphis alternata (Burm.f.) T. Anderson (Syn.) (Strobilanthes alternata (Burm.f.) Moylan ex J.R.I. Wood) | Low maintenance [13] |
| 2 | Alternanthera bettzickiana (Regel) G. Nicholson | - |
| 3 | Alternanthera ficoidea (L.) P. Beauv. | Low maintenance [14] |
| 4 | Allium schoenoprasum L. | Low maintenance [15] |
| 5 | Zephyranthes candida (Lindl.) Herb. | Medium maintenance [16] |
| 6 | Trachelospermum jasminoides (Lindl.) Lem. | Medium maintenance and heavy shade tolerance [17] |
| 7 | Liriope muscari (Decne.) LH. Bailey | Drought and air pollution tolerance [18] |
| 8 | Aloe sp. | Low maintenance and drought tolerance [19] |
| 9 | Dianella caerulea Sims | Low maintenance and drought tolerance [20] |
| 10 | Achillea millefolium L. | Heat and drought tolerance [21] |
| 11 | Coreopsis rosea Nutt. | Drought tolerance [22] |
| 12 | Leucanthemum vulgare (Vaill.) Lam. | Drought tolerance [23] |
| 13 | Cyanotis obtusa (Trimen) Trimen (Syn.) | Drought tolerance [24] |
| 14 | Convolvulus sabatius Viv. | Drought tolerance [25] |
| 15 | Sedum x rubrotinctum R.T. Clausen | Drought tolerance [26] |
| 16 | Fimbristylis ovata (Burm.f.) J.Kern | - |
| 17 | Rhynchospora nervosa (Vahl) Boeckeler | - |
| 18 | Eleocharis sp. | Low maintenance [27] |
| 19 | Acalypha wilkesiana Müll.Arg. | Low maintenance [28] |
| 20 | Plectranthus scutellarioides (L.) R.Br. | Low maintenance [29] |
| 21 | Heterocentron elegans Kuntze | Partial drought tolerance [30] |
| 22 | Phyllanthus myrtifolius (Wight) Müll.Arg. | - |
| 23 | Pennisetum setaceum (Forssk.) Chiov. (Syn.)(Cenchrus setaceus (Forssk.) Morrone.) | Low maintenance and air pollution tolerance [31] |
| 24 | Pilea libanensis Urb. | - |
| 25 | Lantana camara L. | Low maintenance and drought tolerance [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pichakum, N.; Pichakum, A. Evaluating the Drought Endurance of Landscaping Ground Cover Plants in a Roof Top Model. Horticulturae 2021, 7, 31. https://doi.org/10.3390/horticulturae7020031
Pichakum N, Pichakum A. Evaluating the Drought Endurance of Landscaping Ground Cover Plants in a Roof Top Model. Horticulturae. 2021; 7(2):31. https://doi.org/10.3390/horticulturae7020031
Chicago/Turabian StylePichakum, Nath, and Aussanee Pichakum. 2021. "Evaluating the Drought Endurance of Landscaping Ground Cover Plants in a Roof Top Model" Horticulturae 7, no. 2: 31. https://doi.org/10.3390/horticulturae7020031
APA StylePichakum, N., & Pichakum, A. (2021). Evaluating the Drought Endurance of Landscaping Ground Cover Plants in a Roof Top Model. Horticulturae, 7(2), 31. https://doi.org/10.3390/horticulturae7020031