
Physiology Response and Resistance Evaluation of Twenty Coconut Germplasm Resources under Low Temperature Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Assay of Relative Electrical Conductivity and Semi-Lethal Temperature
2.3. Determination of Osmotic Adjustment Substance Content
2.4. Determination of Defense Enzyme Activity
2.5. Data and Statistical Analysis
3. Results and Discussion
3.1. REC and LT50
3.2. Osmotic Adjustment Substance Content
3.2.1. MDA Content
3.2.2. SP Content
3.2.3. Pro Content
3.3. Defense Enzyme Activity
3.4. Correlation Analysis of Physiological Indexes
3.5. Principal Component Analysis of Physiological Indexes
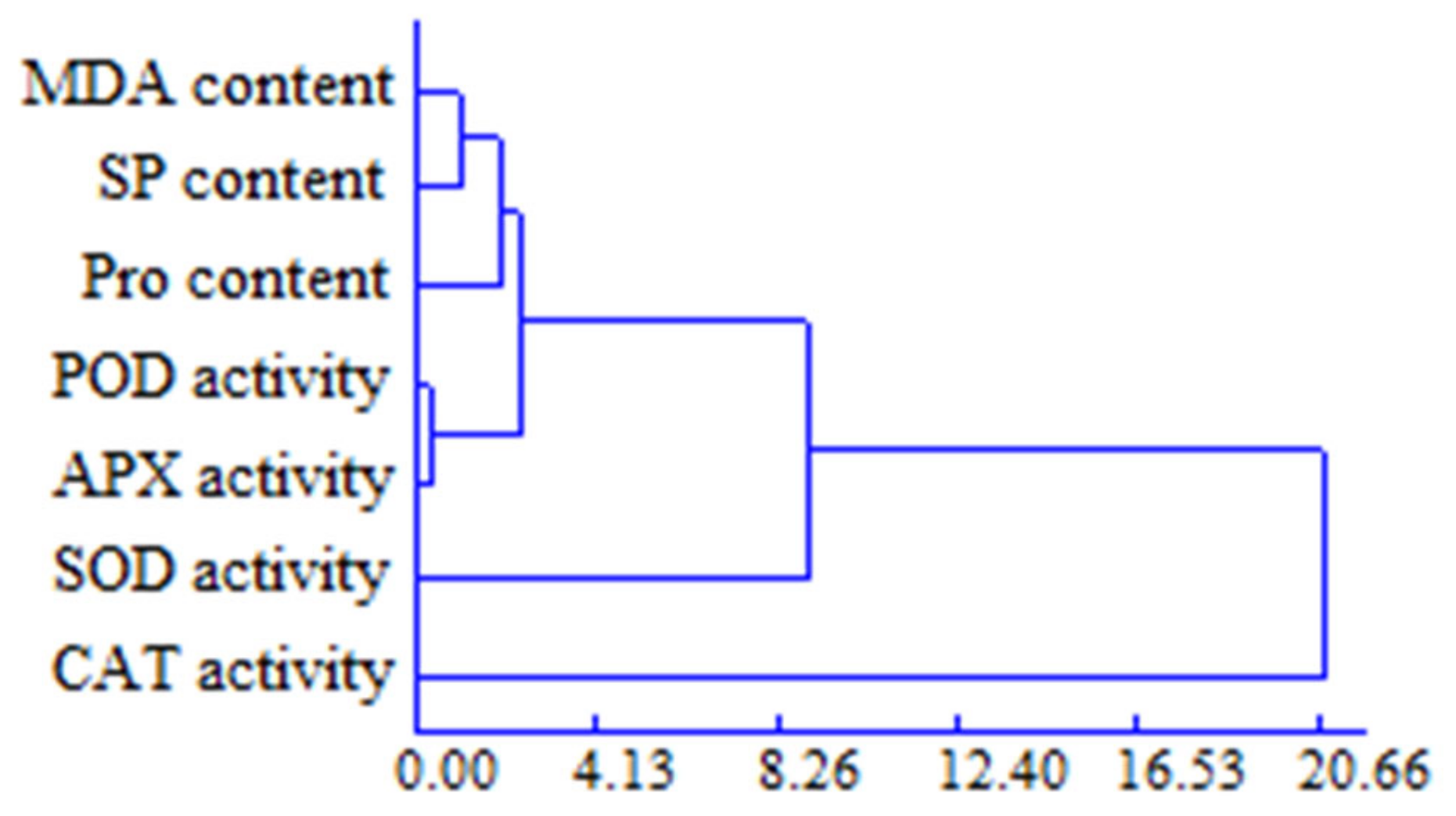
3.6. Cluster Analysis of Physiological Indexes
3.7. Comprehensive Evaluation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, M.L.; Li, J.; Wang, B.Z. Progress of comprehensive research on coconut in China. China Trop. Agric. 2007, 5, 30–31. [Google Scholar]
- Kannaian, U.P.N.; Edwin, J.B.; Rajagopal, V.; Shankar, S.N.; Srinivasan, B. Phytochemical composition and antioxidant activity of coconut cotyledon. Heliyon 2020, 6, e03411. [Google Scholar] [CrossRef]
- Freitas, N.M.; Pereira, T.N.S.; Geronimo, I.G.C.; Azevedo, A.O.N.; Ramos, S.R.R.; Pereira, M.G. Coconut genome size determined by flow cytometry: Tall versus Dwarf types. Genet. Mol. Res. 2016, 15, gmr.15017470. [Google Scholar] [CrossRef] [PubMed]
- Lédo, A.S.; Passos, E.E.M.; Fontes, H.R.; Ferreira, J.M.S.; Talamini, V.; Vendrame, W.A. Advances in coconut palm propagation. Rev. Bras. Frutic. 2019, 41, e159. [Google Scholar] [CrossRef]
- Zhang, R.N.; Zhou, B.; Xu, S.T. Discussion and consideration of plant landscape design in college campus under the background of beautiful campus: Taking Haidian campus of Hainan University as an example. Tianjin Agric. Sci. 2020, 26, 85–90. [Google Scholar]
- Maria, M.S.S.; Claudivan, F.L.; Antonia, L.R.N.; Carlos, H.C.S.; Aureliano, A.R.; Marlos, A.B.; Isabel, C.S.A.; Hans, R.G. Ecophysiology of the tall coconut growing under different coastal areas of northeastern Brazil. Agr. Water Manag. 2020, 232, 106047. [Google Scholar]
- Ruan, Z.P.; Tang, Y.J.; Zeng, M.J. Influence of drought stress on photosynthetic characteristics and activity of antioxidant enzymes of four species of palm seedlings. Chin. J. Trop. Crop. 2016, 37, 1914–1919. [Google Scholar]
- Zhang, Y.X.; Jiang, M. Physiological experiment on drought resistance of different coconut varieties. World Trop. Agric. Info. 1996, 9, 15. [Google Scholar]
- Gong, S.F.; Chen, S.T. The change of osmoregulation substance content in different varieties of coconut at the nursery stage under salt stress. Chin. J. Trop. Agric. 2018, 38, 1–5. [Google Scholar]
- Hebbar, K.B.; Neethu, P.; Sukumar, P.A.; Sujithra, M.; Santhosh, A.; Ramesh, S.V.; Niral, V.; Hareesh, G.S.; Nameer, P.O.; Prasad, P.V.V. Understanding physiology and impacts of high temperature stress on the progamic phase of coconut (Cocos nucifera L.). Plants 2020, 9, 1651. [Google Scholar] [CrossRef]
- Cao, H.X.; Lei, X.T.; Liu, Y.J.; Sun, C.X.; Zhang, R.L. Identification and comprehensive evaluation of cold resistance indexes of coconut. Guangdong Agric. Sci. 2016, 43, 49–54. [Google Scholar]
- Cao, H.X.; Yan, C.B.; Feng, M.L.; Zhang, R.L. Effects of low temperature stress on leaf nutrient content of different coconut cultivars. China Trop. Agric. 2014, 2, 77–79. [Google Scholar]
- Cao, H.X.; Huang, H.J.; Lei, X.T.; Zhang, D.P.; Zhang, R.L. Difference of the leaf anatomical structure of coconut varieties under low temperature treatments. Chin. J. Trop. Crop. 2014, 35, 2420–2425. [Google Scholar]
- Xu, W.J.; Yuan, T.; Yang, Y.G.; Yu, S.Q. The physiological response to low temperature stress of six palme plants in Nanchang. Acta Agric. Univ. Jiangxiensis 2013, 35, 1212–1216. [Google Scholar]
- Zhang, J.; Huang, G.R.; Sun, C.X.; Fan, H.K. Coconut chilling and phenology analysis of Hainan. Chin. J. Trop. Crop. 2016, 36, 50–54. [Google Scholar]
- Zhou, G.S.; Mei, F.Z.; Zhou, Z.Q.; Zhu, X.T. Comprehensive evaluation and forecast on physiological indices of waterlogging resistance of different wheat varieties. Sci. Agric. Sin. 2003, 11, 1378–1382. [Google Scholar]
- Tang, L.X.; Niu, C.; Yang, W.B. Cold damage to coconut and countermeasures in China. Chin. J. Trop. Agric. 2011, 31, 92–94. [Google Scholar]
- Liu, Z.X.; Cheng, X.Z.; Zhou, G.M.; Hou, K.H. An entropy-based evaluation model for multiple objective decision making on adzuki bean germplasm. J. Plant Genet. Res. 2011, 12, 54–58. [Google Scholar]
- Zhang, Q.; Wei, Z.W.; Yan, T.F.; Geng, X.L. Identification and evaluation of genetic diversity of agronomic traits in oat germplasm resources. Acta Agrestia Sin. 2021, 29, 309–316. [Google Scholar]
- Fan, Y.; Zhao, X.F.; Liu, X.G.; Yang, Y. Factor analysis and comprehensive assessment for agronomic traits of peanut. J. Hebei Agric. Sci. 2015, 19, 80–83. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Zhu, G.H.; Liu, Z.Q.; Zhu, P.R. Study on the determination of low temperature semi-lethal temperature of plant tissue by Logistic equation. J. Nanjing Agric. Univ. 1986, 3, 11–16. [Google Scholar]
- Wang, X.K.; Huang, J.L. Principles and Techniques of Plant Physiological Biochemical Experiment, 3rd ed.; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Guo, H.J.; Huang, Z.J.; Li, M.Q.; Hou, Z.N. Growth, ionic homeostasis, and physiological responses of cotton under different salt and alkali stresses. Sci. Rep. 2020, 10, 21844. [Google Scholar] [CrossRef]
- Jiang, L.N.; Zhang, D.J.; Song, F.; Liu, P.; Fan, T.T.; Yu, H.B.; Li, C.X. Evaluation of cold resistance of different wheat varieties based on physiological responses of leaves to low temperature at the jointing stage. Acta Ecol. Sin. 2014, 34, 4251–4261. [Google Scholar]
- Tian, Z.G.; Wang, F.; Zhang, W.N.; Liu, C.M.; Zhao, X.M. Antioxidant mechanism and lipid peroxidation patterns in leaves and petals of marigold in response to drought stress. Hortic. Environ Biote. 2012, 53, 183–192. [Google Scholar] [CrossRef]
- Cao, H.X.; Song, W.Y.; Sun, C.X.; Chen, S.T.; Tang, L.X.; Zhao, S.L. Measurement of coldness by electrical conductivity method in associated with the Logistic equation in coconut seedlings. Guihaia 2009, 29, 510–513. [Google Scholar]
- Cheng, J.H.; Zhang, M.L.; Wang, C.; Yan, S.P. Effect of low temperature stress on physiological index of four strawberry varieties. J. Northeast Agric. Sci. 2021, 46, 85–88. [Google Scholar]
- Armstrong, J.J.; Takebayashi, N.; Sformo, T.; Wolf, D.E. Cold tolerance in Arabidopsis kamchatica. Am. J. Bot. 2015, 102, 439–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, M.M.; Friesen, P.C.; Sage, R.F. Winter cold-tolerance thresholds in field-grown Miscanthus hybrid rhizomes. J. Exp. Bot. 2015, 66, 4415–4425. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Hu, Y.; Chen, B.H.; Zhu, Y.F.; Mohammed, M.D.; Sofkova, S. Physiological mechanisms of resistance to cold stress associated with 10 elite apple rootstocks. J. Integr. Agr. 2018, 17, 857–866. [Google Scholar] [CrossRef]
- Guo, H.H.; Gao, S.M.; Zhao, F.J.; Li, F.L. Effects of cold acclimation on several enzyme activities in Euonymus radicans ‘Emorald & Gold’ and its relation to semi-lethal temperature. Stud. China 2004, 1, 10–17. [Google Scholar]
- Wang, J.; Guo, J.P.; Zhang, Y.X.; Yan, X.R. Integrated transcriptomic and metabolomic analyses of yellow horn (Xanthoceras sorbifolia) in response to cold stress. PLoS ONE 2020, 15, e0236588. [Google Scholar] [CrossRef]
- Khaliq, G.; Mohamed, M.T.M.; Ghazali, H.M.; Ding, P.; Ali, A. Influence of gum arabic coating enriched with calcium chloride on physiological, biochemical and quality responses of mango (Mangifera indica L.) fruit stored under low temperature stress. Postharvest Biol. Tec. 2016, 111, 362–369. [Google Scholar] [CrossRef]
- Zhang, R.G.; Guo, X.C.; Zhang, Y.L.; Tian, C.R. Influence of modified atmosphere treatment on post-harvest reactive oxygen metabolism of pomegranate peels. Nat. Prod. Res. 2020, 34, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Guo, Q.; Li, Q.P.; Ha, Y.M.; Li, X.P.; Chen, W.X. Impact of postharvest nitric oxide treatment on antioxidant enzymes and related genes in banana fruit in response to chilling tolerance. Postharvest Biol. Tec. 2014, 92, 157–163. [Google Scholar] [CrossRef]
- Zhu, S.P.; Nong, J.F.; Luo, G.T.; Li, Q.P.; Wang, F.S.; Jiang, D.; Zhao, X.C. Varied tolerance and different responses of five citrus rootstocks to acid stress by principle component analysis and orthogonal analysis. Sci. Hortic. 2021, 278, 109853. [Google Scholar] [CrossRef]
- Sun, J.; You, X.R.; Li, L.; Peng, H.X.; Su, W.Q.; Li, C.B.; He, Q.G.; Liao, F. Effects of a phospholipase D inhibitor on postharvest enzymatic browning and oxidative stress of litchi fruit. Postharvest Biol. Tec. 2011, 62, 288–294. [Google Scholar] [CrossRef]
- Meethaworn, K.; Luckanatinwong, V.; Zhang, B.; Chen, K.S.; Siriphanich, J. Off-flavor caused by cold storage is related to induced activity of LOX and HPL in young coconut fruit. LWT-Food Sci. Technol. 2019, 114, 108329. [Google Scholar] [CrossRef]
- Meethaworn, K.; Siriphanich, J. Postharvest behavior during storage of young coconut (Cocos nucifera L.) at different temperatures. Acta Hortic. 2015, 1091, 125–131. [Google Scholar] [CrossRef]
- Phornvillay, S.; Prongprasert, N.; Wongs-Aree, C.; Uthairatanakij, A.; Srilaong, V. Physio-biochemical responses of okra (Abelmoschus esculentus) to oxidative stress under low temperature storage. Hortic. J. 2020, 89, 69–77. [Google Scholar] [CrossRef]
- Mostafa, K.M.; Shinnosuke, I.; Fuminori, T.; Ko, S.; Masaharu, K.; Taishi, U.; Kazuo, S.; Yukio, K.; Matsuo, U. Large-scale phosphoproteomic study of Arabidopsis membrane proteins reveals early signaling events in response to cold. Int. J. Mol. Sci. 2020, 21, 8631. [Google Scholar]
- Yang, H.J.; Guo, H.C. Physiological responds of potato seedlings to low temperature stress and comprehensive evaluation on their cold tolerance. Southwest China J. Agric. Sci. 2016, 29, 2560–2566. [Google Scholar]
- Bagwell, S.N.; Ricker, J.V. Antifreeze proteins: Effective adaptations of organisms for low temperature survival. Bios 2019, 90, 170–185. [Google Scholar] [CrossRef]
- Yang, Y.D.; Saand, M.A.; Abdelaal, W.B.; Zhang, J.; Wu, Y.; Li, J.; Fan, H.K.; Wang, F.Y. iTRAQ-based comparative proteomic analysis of two coconut varieties reveals aromatic coconut cold-sensitive in response to low temperature. J. Proteom. 2020, 220, 103766. [Google Scholar] [CrossRef]
- Meenakshi, G.; Navreet, K. Low temperature induced oxidative stress tolerance in oats (Avena sativa L.) genotypes. Indian J. Plant Physiol. 2018, 23, 316–324. [Google Scholar]
- Aghaee, A.; Moradi, F.; Zare-Maivan, H.; Zarinkamar, F.; Irandoost, H.P.; Sharifi, P. Physiological responses of two rice (Oryza sativa L.) genotypes to chilling stress at seedling stage. Afr. J. Biotechnol. 2011, 10, 7617–7621. [Google Scholar]
- Koç, E.; İşlek, C.; Üstün, A.S. Effect of cold on protein, proline, phenolic compounds and chlorophyll content of two pepper (Capsicum annuum L.) varieties. Gazi Univ. J. Sci. 2010, 23, 1–6. [Google Scholar]
- Wang, Y.H.; Xiong, F.; Nong, S.H.; Liao, J.R.; Xing, A.Q.; Shen, Q.; Ma, Y.C.; Fang, W.P.; Zhu, X.J. Effects of nitric oxide on the GABA, polyamines, and proline in tea (Camellia sinensis) roots under cold stress. Sci. Rep. 2020, 10, 12240. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, L.; Xu, Y.Q.; Limwachiranon, J.; Li, D.; Ban, Z.J.; Luo, Z.S. Effect of exogenous nitro oxide on chilling tolerance, polyamine, proline and gamma-aminobutyric acid and in bamboo shoots (Phyllostachys praecox f. prevernalis). J. Agr. Food Chem. 2017, 65, 5607. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Wang, M.L.; Hao, J.; Chen, X.H.; Zhang, X.C. SlMYB102 expression enhances low-temperature stress resistance in tomato plants. PeerJ 2020, 8, e10059. [Google Scholar] [CrossRef] [PubMed]
- Castañares, J.L.; Bouzo, C.A. Effect of exogenous melatonin on seed germination and seedling growth in melon (Cucumis melo L.) under salt stress. Hortic. Plant J. 2019, 5, 79–87. [Google Scholar] [CrossRef]
- Szekely-Varga, Z.; González-Orenga, S.; Cantor, M.; Jucan, D.; Boscaiu, M.; Vicente, O. Effects of drought and salinity on two commercial varieties of lavandula angustifolia mill. Plants 2020, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Wang, Q.S.; Wang, X.G.; Zhang, J.H.; Zhang, Y.C.; Zhang, X.Z.; Bie, S.; Xia, S.B. Physiological response and transcriptome analysis of cotton leaf in the two-leaf period under low temperature. Hubei Agric. Sci. 2020, 59, 169–176. [Google Scholar]
- Wang, Z.Y. Effect of low temperature stress on proline content of elderberry. Agric. Jilin 2019, 2, 60. [Google Scholar]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase–mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. R. 2015, 22, 10375–10394. [Google Scholar] [CrossRef] [PubMed]
- Eraslan, F.; Inal, A.; Savasturk, O.; Gunes, A. Changes in antioxidative system and membrane damage of lettuce in response to salinity and boron toxicity. Sci. Hortic. 2007, 114, 5–10. [Google Scholar] [CrossRef]
- Usue, P.L.; Anabel, R.; Maite, L.; Cristina, S.; Alberto, M.R.; Flavia, N.I.; Amaia, M.P. The oxidative stress caused by salinity in two barley cultivars is mitigated by elevated CO2. Physiol. Plant. 2009, 135, 29–42. [Google Scholar]
- Li, Y. Physiological responses of tomato seedlings (Lycopersicon Esculentum) to salt stress. Mod. Appl. Sci. 2009, 3, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Han, G.; Dang, Q.; Zhao, Z. Responses of antioxidation protective system of caragana korshinskii Kom.to drought stress. Acta Agrestia Sin. 2010, 18, 528–532. [Google Scholar]
- Zhang, H.M.; Jin, H.J.; Ding, X.T.; He, L.Z.; Cui, J.W.; Yu, J.Z. Physiological response and resistance evaluation of grafted cucumber seedlings with different rootstocks under temperature stress. Acta Bot. Boreali-Occident. Sin. 2019, 39, 1259–1269. [Google Scholar]
- Tian, X.H.; Jie, J.M. Physiological response of different edible lily varieties to low temperature stress. Guizhou Agric. Sci. 2020, 48, 89–93. [Google Scholar]
- Fariborz, H.; Asghar, R.; Fabián, G.; Domingo, M.R.; María, S.; Daniel, V. Susceptibility of blood orange cultivars to chilling injury based on antioxidant system and physiological and biochemical responses at different storage temperatures. Foods 2020, 9, 1609. [Google Scholar]
- Ali, S.; Khan, A.S.; Anjum, M.A.; Nawaz, A.; Hussain, S. Effect of postharvest oxalic acid application on enzymatic browning and quality of lotus (Nelumbo nucifera Gaertn.) root slices. Food Chem. 2020, 312, 126051. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Slooten, L.; Vandenbranden, S.; Rycke, R.D.; Botterman, J.; Sybesma, C.; Montagu, M.V.; Inzé, D. Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J. 1991, 10, 1723–1732. [Google Scholar] [CrossRef]
- Zhang, D.D.; Xu, X.F.; Zhang, Z.K.; Jiang, G.X.; Feng, L.Y.; Duan, X.W.; Jiang, Y.M. 6-Benzylaminopurine improves the quality of harvested litchi fruit. Postharvest Biol. Tec. 2018, 143, 137–142. [Google Scholar] [CrossRef]
- Zuo, J.H.; Chen, A.J.; Sun, A.D.; Luo, Y.B.; Zhu, B.Z. Research progress on the factors related to tomato fruit ripening and senescence. Sci. Agric. Sin. 2010, 43, 2724–2734. [Google Scholar]
- Zhu, X.M.; Shi, X.P.; Yong, D.J.; Zhang, Y.; Li, B.H.; Liang, W.X.; Wang, C.X. Induction of resistance against glomerella cingulata in apple by endophytic actinomycetes strain A-1. Plant Physiol. Comm. 2015, 51, 949–954. [Google Scholar]
- Wang, Y.S.; Luo, Z.S.; Du, R.X. Nitric oxide delays chlorophyll degradation and enhances antioxidant activity in banana fruits after cold storage. Acta Physiol. Plant. 2015, 37, 74. [Google Scholar] [CrossRef]
- Guo, Z.; Ge, B.; Han, M.L.; Tao, W.L.; Hu, Y.; Chen, G.K. Analysis of defense enzyme activities and transcriptome of clubroot-resistant and-susceptible in canola to clubroot pathogen Plasmodiophora brassicae during early infection. J. Plant Protec. 2018, 45, 290–298. [Google Scholar]
- Zhang, Q.M.; Wang, C.X.; Yong, D.J.; Li, G.F.; Dong, X.L.; Li, B.H.; Guilhermino, L. Induction of resistance mediated by an attenuated strain of valsa mali var. mali using pathogen-apple callus interaction system. Sci. World J. 2014, 2014, 201382. [Google Scholar]
- Jia, K.; Yan, H.Z.; Gao, J. Effects of exogenous MeJA on growth and antioxidant enzyme activity of turnip seedlings. Xinjiang Agric. Sci. 2020, 57, 806–813. [Google Scholar]
- Zhang, Z.K.; Huber, D.J.; Qu, H.X.; Yun, Z.; Wang, H.; Huang, Z.H.; Huang, H.; Jiang, Y.M. Enzymatic browning and antioxidant activities in harvested litchi fruit as influenced by apple polyphenols. Food Chem. 2015, 171, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Khan, A.S.; Nawaz, A.; Anjum, M.A.; Naz, S.; Ejaz, S.; Hussain, S. Aloe vera gel coating delays postharvest browning and maintains quality of harvested litchi fruit. Postharvest Biol. Tec. 2019, 157, 110960. [Google Scholar] [CrossRef]
- Jimenez, A.; Creissen, G.; Kular, B.; Firmin, J.; Robinson, S.; Verhoeyen, M.; Mullineaux, P. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 2002, 214, 751–758. [Google Scholar] [CrossRef]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 15. [Google Scholar] [CrossRef]
- Landberg, T.; Greger, M. Differences in oxidative stress in heavy metal resistant and sensitive clones of Salix viminalis. J. Plant Physiol. 2002, 159, 69–75. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Phys. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signaling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Sharma, I.; Ahmad, P. Chapter 4—Catalase: A versatile antioxidant in plants. In Oxidative Damage to Plants; Academic Press: San Diego, CA, USA, 2014; pp. 131–148. [Google Scholar]
- Caverzan, A.; Passaia, G.; Barcellos, R.S.; Werner, R.C.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Kavousi, H.R.; Karimi, M.R.; Neghab, M.G. Assessment the copper-induced changes in antioxidant defense mechanisms and copper phytoremediation potential of common mullein (Verbascum thapsus L.). Environ. Sci. Pollut. Res. Int. 2021, 28, 18070–18080. [Google Scholar] [CrossRef]
- Sun, W.H.; Wang, W.Q.; Meng, Q.W. Functional Mechanism and Enzymatic and Molecular Characteristic of Ascorbate Peroxidase in Plants. Plant Physiol. Commun. 2005, 41, 143–147. [Google Scholar]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Ali, S.; Khan, A.S.; Malik, A.U.; Anjum, M.A.; Nawaz, A.; Shah, H.M.S. Modified atmosphere packaging delays enzymatic browning and maintains quality of harvested litchi fruit during low temperature storage. Sci. Hortic. 2019, 254, 14–20. [Google Scholar] [CrossRef]
- Shah, H.M.S.; Khan, A.S.; Ali, S. Pre-storage kojic acid application delays pericarp browning and maintains antioxidant activities of litchi fruit. Postharvest Biol. Tec. 2017, 132, 154–161. [Google Scholar] [CrossRef]
- Campos, M.K.F.; Carvalho, K.; Souza, F.S.; Marur, C.J.; Pereira, L.F.P.; Filho, J.C.B.; Vieira, L.G.E. Drought tolerance and antioxidant enzymatic activity in transgenic ‘Swingle’ citrumelo plants over-accumulating proline. Environ. Exp. Bot. 2011, 72, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.L. Effects of Exogenous Salicylic Acid and Abscisic Acid on Growth and Physiological Characteristics of Medicago polymorpha under High Temperature Stress. Yangzhou Univ. 2021. [Google Scholar]
- Moieni-Korbekandi, Z.; Karimzade, G.; Sharifi, M. Cold-induced changes of proline, malondialdehyde and chlorophyll in spring canola cultivars. J. Plant Physiol. Breed. 2014, 4, 1–11. [Google Scholar]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Maji, A.T.; Shaibu, A.A. Application of principal component analysis for rice germplasm characterization and evaluation. J. Breed. Crop Sci. 2012, 4, 87–93. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.P.; Downey, G.; Rawson, A.; Warriner, K.; Gernigon, G. Application of principal component and hierarchical cluster analysis to classify fruits and vegetables commonly consumed in Ireland based on in vitro antioxidant activity. J. Food Compos. Anal. 2010, 24, 250–256. [Google Scholar] [CrossRef]
- Liu, W.; Guo, H.J.; Qin, J.P.; Luo, Y.D.; Zheng, L.Z. Quality control of Vernonia patula Merr based upon multi-component determination and principal component analysis. Chin. J. Hosp. Pharm. 2021, 41, 135–138. [Google Scholar]
- Kong, D.W.; Chen, D.Q.; Zhou, L.Q.; Wang, Y.P.; Li, S.G. The principle components analysis of several agronomic and yield traits of rice. Chin. Agric. Sci. Bull. 2005, 21, 117–119. [Google Scholar]
- Suzana, M.; Zulkifli, Y.; Marhalil, M.; Rajanaidu, N.; Meilina, O.A. Principal component and cluster analyses on Tanzania oil palm Elaeis guineensis JACQ. germplasm. J. Oil Palm Res. 2020, 32, 24–33. [Google Scholar]
- Tang, Z.C. The accumulation of free proline and its roles in water-stressed sorghum seedlings. Physiol. Mol. Biol. Pla. 1989, 1, 105–110. [Google Scholar]
- Jiang, M.Y.; Guo, S.C.; Zhang, X.M. Antioxidant effects of proline accumulation in rice seedlings under oxidative stress. Plant Physiol. J. 1997, 4, 347–352. [Google Scholar]
- Zhang, J.; Zhang, F.; Tian, J.X.; Zhang, G.Y.; Wang, J.S.; Li, H.Z. Response of cucubita pepo seedlings to low temperature and weak light and evaluation index screening. North. Hortic. 2021, 4, 1–9. [Google Scholar]
- Biglari, F.; AlKarkhi, A.F.M.; Easa, A.M. Cluster analysis of antioxidant compounds in dates (Phoenix dactylifera): Effect of long-term cold storage. Food Chem. 2008, 112, 998–1001. [Google Scholar] [CrossRef]
- Saraçli, S.; Doğan, N.; Doğan, İ. Comparison of hierarchical cluster analysis methods by cophenetic correlation. J. Inequal. Appl. 2013, 1, 1–8. [Google Scholar] [CrossRef]
- Ling, F.; Jiao, J.; Li, C.Z.; Jin, Q.X.; Zhao, M.L. Physiological response and comprehensive evaluation of cold resistance under cold stress for different varieties of olea europaea. Acta Bot. Boreali-Occident. Sin. 2015, 35, 508–515. [Google Scholar]
- Li, Q.Y.; Lu, B.; Zhao, J.W.; Li, H.; Li, Y.; Miao, S.Y.; Lu, B.S. Physiological response and cold resistance evaluation of different Pyrus calleryan varieties under low temperature stress. J. NW For. Univ. 2020, 48, 86–94. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resource Number | Place of Origin | Type |
|---|---|---|
| C1 | China | tall coconut |
| C2 | China | tall coconut |
| C3 | China | tall coconut |
| C4 | China | tall coconut |
| C5 | China | tall coconut |
| C6 | Malaysia | dwarf coconut |
| C7 | China | tall coconut |
| C8 | China | dwarf coconut |
| C9 | Vietnam | dwarf coconut |
| C10 | China | dwarf coconut |
| C11 | China | tall coconut |
| C12 | Vietnam | dwarf coconut |
| C13 | China | dwarf coconut |
| C14 | China | tall coconut |
| C15 | Malaysia | dwarf coconut |
| C16 | Thailand | dwarf coconut |
| C17 | Thailand | dwarf coconut |
| C18 | China | tall coconut |
| C19 | China | tall coconut |
| C20 | Vietnam | dwarf coconut |
| Correlation Coefficient | MDA Content | SP Content | Pro Content | SOD Activity | POD Activity | CAT Activity | APX Activity |
|---|---|---|---|---|---|---|---|
| MDA content | 1.00 | ||||||
| SP content | 0.18 | 1.00 | |||||
| Pro content | 0.01 | 0.76 ** | 1.00 | ||||
| SOD activity | 0.73 ** | 0.37 ** | 0.16 | 1.00 | |||
| POD activity | 0.81 ** | 0.23 * | 0.11 | 0.80 ** | 1.00 | ||
| CAT activity | 0.74 ** | −0.02 | −0.19 | 0.70 ** | 0.80 ** | 1.00 | |
| APX activity | 0.41 ** | 0.57 ** | 0.57 ** | 0.59 ** | 0.59 ** | 0.29 ** | 1.00 |
| Physiological Indexes | Principal Component 1 | Principal Component 2 | Principal Component 3 |
|---|---|---|---|
| MDA content | 0.43 | −0.23 | 0.31 |
| SP content | 0.24 | 0.54 | 0.55 |
| Pro content | 0.16 | 0.62 | 0.05 |
| SOD activity | 0.46 | −0.07 | 0.09 |
| POD activity | 0.47 | −0.16 | −0.15 |
| CAT activity | 0.39 | −0.37 | 0.10 |
| APX activity | 0.38 | 0.31 | −0.75 |
| Contribution rate/% | 54.52 | 28.91 | 5.43 |
| Cumulative contribution rate/% | 54.52 | 83.43 | 88.86 |
| Coconut Resources | MDA Content | SP Content | Pro Content | SOD Activity | POD Activity | CAT Activity | APX Activity | Mean Membership Function | Rank |
|---|---|---|---|---|---|---|---|---|---|
| C1 | 0.14 | 0.34 | 0.03 | 2.45 | 0.40 | 0.24 | 0.19 | 0.54 | 13 |
| C2 | 0.25 | 1.28 | 0.82 | 2.96 | 1.25 | 0.58 | 1.19 | 1.19 | 2 |
| C3 | 0.02 | 1.19 | 0.33 | 1.73 | 2.56 | 0.84 | 0.90 | 1.08 | 4 |
| C4 | 0.09 | 0.17 | 0.11 | 1.04 | 0.24 | 0.96 | 0.38 | 0.43 | 17 |
| C5 | 0.08 | 0.02 | 0.08 | 2.28 | 0.35 | 0.24 | 0.16 | 0.46 | 16 |
| C6 | 0.93 | 0.50 | 0.11 | 3.50 | 1.11 | 0.18 | 0.53 | 0.98 | 6 |
| C7 | 0.45 | 0.52 | 0.19 | 2.46 | 3.00 | 0.11 | 1.27 | 1.14 | 3 |
| C8 | 0.46 | 0.77 | 0.28 | 0.10 | 0.68 | 1.61 | 0.12 | 0.57 | 11 |
| C9 | 0.01 | 0.19 | 0.06 | 2.43 | 0.45 | 0.44 | 0.26 | 0.55 | 12 |
| C10 | 1.13 | 0.54 | 1.01 | 2.90 | 1.59 | 0.96 | 0.25 | 1.20 | 1 |
| C11 | 0.16 | 0.54 | 0.39 | 1.75 | 2.66 | 0.43 | 0.12 | 0.86 | 8 |
| C12 | 0.18 | 0.05 | 0.17 | 1.02 | 0.10 | 0.09 | 0.31 | 0.27 | 19 |
| C13 | 0.02 | 0.48 | 0.10 | 2.23 | 0.34 | 0.35 | 0.07 | 0.51 | 15 |
| C14 | 1.06 | 0.29 | 0.09 | 2.86 | 1.32 | 0.53 | 0.94 | 1.01 | 5 |
| C15 | 0.32 | 0.37 | 0.01 | 1.43 | 2.84 | 0.51 | 0.72 | 0.89 | 7 |
| C16 | 0.33 | 0.11 | 0.33 | 0.47 | 0.37 | 0.57 | 0.13 | 0.33 | 18 |
| C17 | 0.00 | 0.24 | 0.06 | 2.37 | 0.40 | 0.46 | 0.13 | 0.52 | 14 |
| C18 | 0.83 | 0.28 | 0.34 | 1.65 | 0.72 | 0.07 | 0.60 | 0.64 | 10 |
| C19 | 0.30 | 0.33 | 0.08 | 0.89 | 3.05 | 0.26 | 0.49 | 0.77 | 9 |
| C20 | 0.16 | 0.01 | 0.21 | 0.99 | 0.21 | 0.15 | 0.10 | 0.26 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Zhang, R.; Yuan, Z.; Cao, H.; Martin, J.J.J. Physiology Response and Resistance Evaluation of Twenty Coconut Germplasm Resources under Low Temperature Stress. Horticulturae 2021, 7, 234. https://doi.org/10.3390/horticulturae7080234
Sun C, Zhang R, Yuan Z, Cao H, Martin JJJ. Physiology Response and Resistance Evaluation of Twenty Coconut Germplasm Resources under Low Temperature Stress. Horticulturae. 2021; 7(8):234. https://doi.org/10.3390/horticulturae7080234
Chicago/Turabian StyleSun, Chengxu, Ruining Zhang, Zhouyu Yuan, Hongxing Cao, and Jerome Jeyakumar John Martin. 2021. "Physiology Response and Resistance Evaluation of Twenty Coconut Germplasm Resources under Low Temperature Stress" Horticulturae 7, no. 8: 234. https://doi.org/10.3390/horticulturae7080234
APA StyleSun, C., Zhang, R., Yuan, Z., Cao, H., & Martin, J. J. J. (2021). Physiology Response and Resistance Evaluation of Twenty Coconut Germplasm Resources under Low Temperature Stress. Horticulturae, 7(8), 234. https://doi.org/10.3390/horticulturae7080234