

Physiological Responses of Lettuce (Lactuca sativa L.) to Soil Contamination with Pb

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Pb Treatments
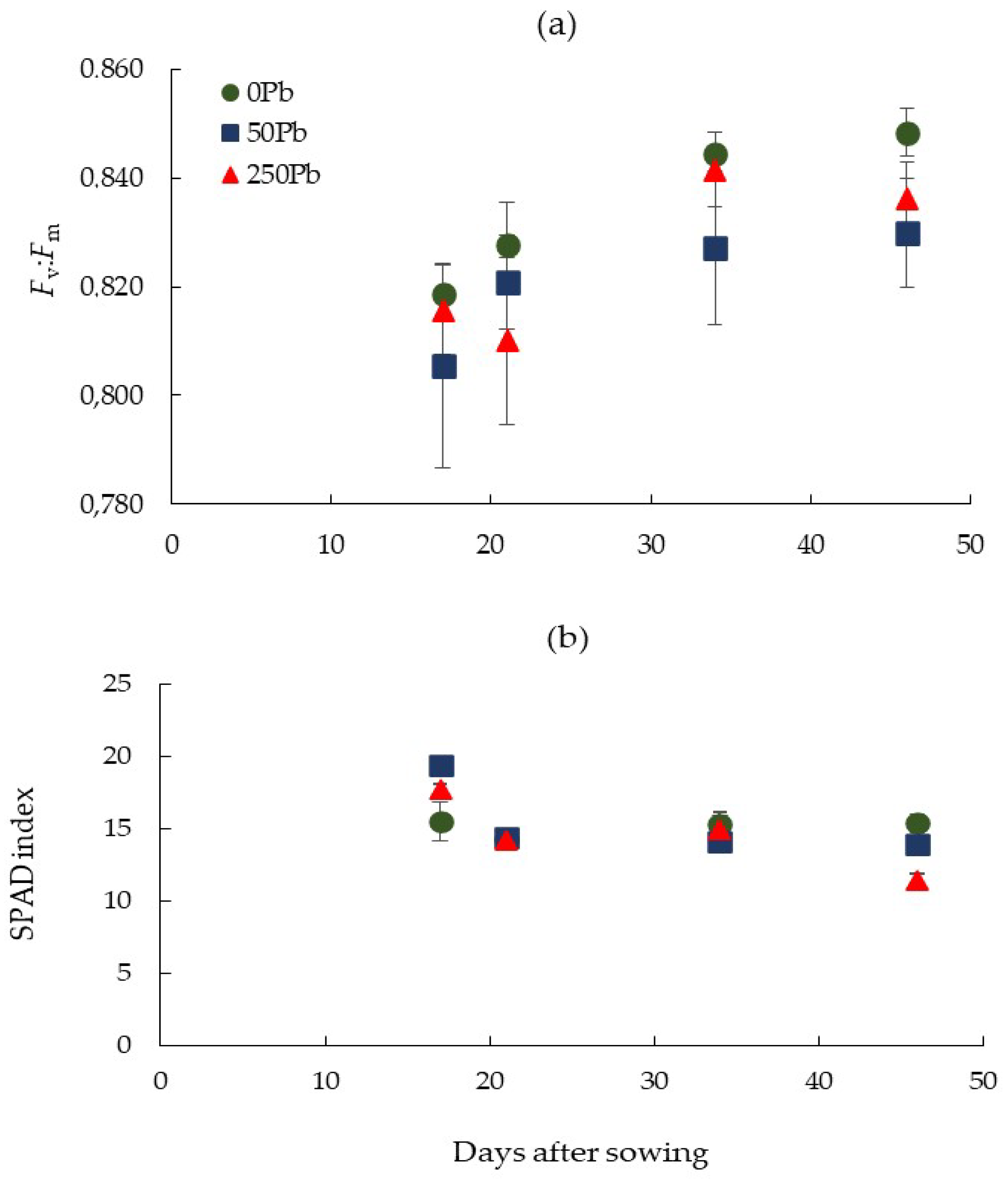
2.2. Chlorophyll Fluorescence and Chlorophyll Content
2.3. CO2 Gas Exchange
2.4. Leaf Relative Water Content
2.5. Mitochondrial Respiratory Pathway
2.6. Plant Growth Parameters
2.7. Leaf and Root Pb Content
2.8. Statistical Analysis
3. Results
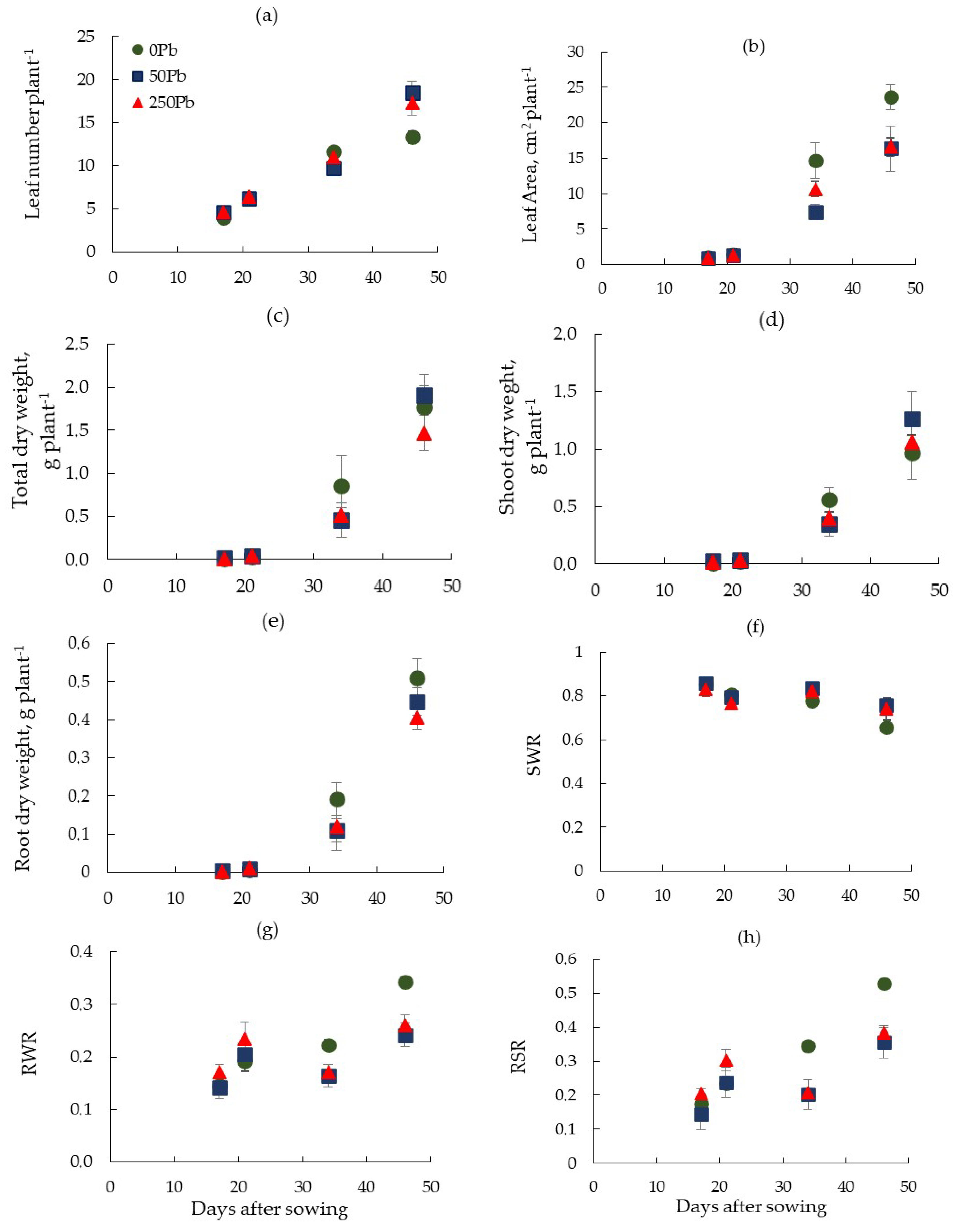
3.1. Plant Growth
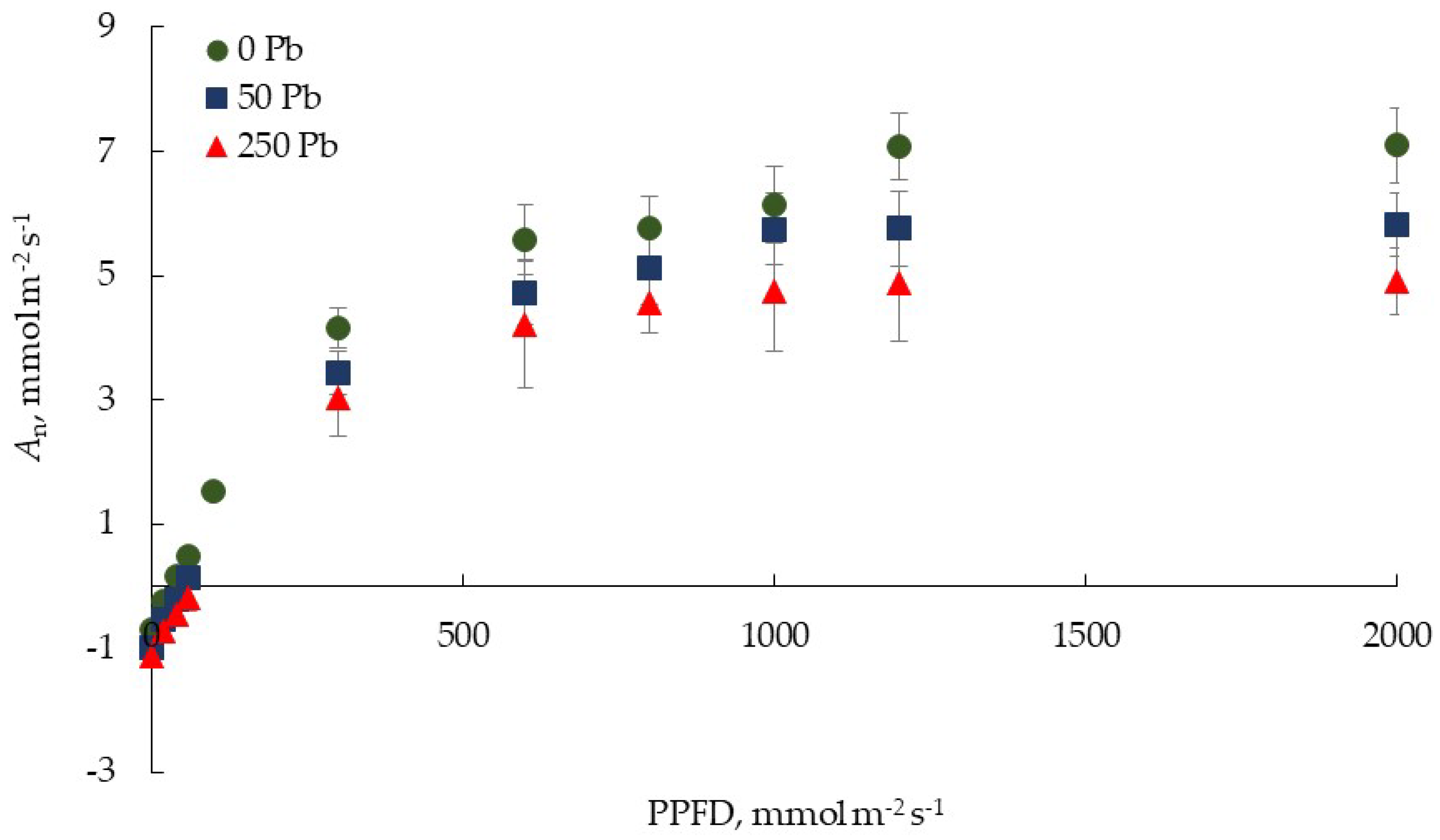
3.2. Effect of Pb on Leaf Gas Exchange of Lettuce
3.3. Pb Content and Accumulation in Shoot and Root of Lettuce
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, Y.; Deng, G.; Guo, H.; Yang, M.; Yang, Q. Accumulation and sub cellular distribution of lead (Pb) in industrial hemp grown in Pb contaminated soil. Ind. Crops Prod. 2021, 161, 113220. [Google Scholar] [CrossRef]
- Nikiforova, E.M.; Kosheleva, N.E. Dynamics of Contamination of Urban Soils with Lead in the Eastern District of Moscow. Eurasian Soil Sci. 2007, 40, 880–892. [Google Scholar] [CrossRef]
- Glazovskaya, M.A. Criteria of Soil Classification According to Lead Contamination Hazard. Pochvovedenie 1994, 4, 110–120. [Google Scholar]
- Markus, J.; McBratney, A.B. A review of the contamination of soil with lead II. Spatial distribution and risk assessment of soil lead. Environ. Int. 2001, 27, 399–411. [Google Scholar] [CrossRef]
- Kumar, B.; Smita, K.; Flores, L.C. Plant mediated detoxification of mercury and lead. Arab. J. Chem. 2017, 10, 2335–2342. [Google Scholar] [CrossRef] [Green Version]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef] [PubMed]
- Capelo, A.; Santos, C.; Loureiro, S.; Pedro, M.A. Phytotoxicity of lead on Lactuca sativa: Effects On growth, mineral nutrition, photosynthetic activity and oxidant metabolism. Fresenius Environ. Bull. 2012, 21, 450–459. [Google Scholar]
- Fahr, M.; Laplaze, L.; Bendaou, N.; Hocher, V.; El Mzibri, M.; Bogusz, D.; Smouni, A. Effect of lead on root growth. Front. Plant Sci. 2013, 4, 175. [Google Scholar] [CrossRef] [Green Version]
- Collin, S.; Baskar, A.; Geevarghese, D.M.; Ali, M.N.V.S.; Bahubali, P.; Choudhary, R.; Lvov, V.; Tovar, G.I.; Senatov, F.; Koppala, S.; et al. Bioaccumulation of lead (Pb) and its effects in plants: A review. J. Hazard. Mater. 2022, 3, 100064. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Stevens, R.G.; Creissen, G.P.; Mullineaux, P.M. Cloning and characterisation of a cytosolic glutathione reductase cDNA from pea (Pisum sativum L.) and its expression in response to stress. Plant Mol. Biol. 1997, 35, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Chugh, L.K.; Sawhney, S.K. Photosynthetic activities of pisum sativum seedlings grown in presence of cadmium. Plant Physiol. Biochem. 1999, 37, 297–303. [Google Scholar] [CrossRef]
- Sabrine, H.; Boutheina, D.; Lassad, C.; Kheder, M.B.; Bousetta, H. Photosynthesis and growth responses of pea Pisum sativum L. Under heavy metals stress. J. Environ. Sci. 2009, 21, 1552–1556. [Google Scholar]
- Zhou, M.; Gong, X.; Ying, W.; Chao, L.; Hong, M.; Wang, L.; Fashui, H. Cerium relieves the inhibition of chlorophyll biosynthesis of maize caused by magnesium deficiency. Biol. Trace Elem. Res. 2011, 143, 468–477. [Google Scholar] [CrossRef]
- Shu, X.; Yin, L.Y.; Zhang, Q.F.; Wang, W.B. Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of photosynthesis to different concentrations of heavy metals in Davidia involucrata. PLoS ONE 2020, 15, e0228563. [Google Scholar]
- Fu, W.G.; Wang, F.K. Effects of high soil lead concentration on photosynthetic gas exchange and chlorophyll fluorescence in Brassica chinensis L. Plant Soil Environ. 2015, 61, 316–321. [Google Scholar]
- Koeppe, D.E.; Miller, R.J. Lead effects on corn mitochondria respiration. Science 1970, 167, 1376–1377. [Google Scholar] [CrossRef]
- Romanowska, E.; Igamberdiev, A.U.; Parys, E.; Gardeström, P. Stimulation of respiration by Pb2π in detached leaves and mitochondria of C3 and C4 plants. Physiol. Plant. 2002, 116, 148–154. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Distribution of cadmium, lead, nickel, and strontium in imbibing maize caryopses. Russ. J. Plant Physiol. 2005, 52, 565–569. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 1998. [Google Scholar]
- Tangahu, B.V.; Abdullah, S.R.S.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M. A Review on HeavyMetals (As, Pb, and Hg) Uptake by Plants through Phytoremediation. Int. J. Chem. Eng. 2011, 20, 939161. [Google Scholar]
- Gupta, D.K.; Huang, H.G.; Corpas, F.J. Lead tolerance in plants: Strategies for phytoremediation. Environ. Sci. Pollut. Res. Int. 2013, 20, 2150–2161. [Google Scholar] [CrossRef]
- Đurdecic, B.; Lisjak, M.; Stosic, M.; Engler, M.; Popovic, B. Influence of Pb and Cu Toxicity on Lettuce Photosynthetic Pigments and Dry Matter Accumulation, 7th ed.; Alps-Adria Scientific Workshop: Stara Lesna, Slovakia, 2008; pp. 1951–1954. [Google Scholar]
- Silva, S.; Pinto, G.; Santos, C. Low doses of Pb affected Lactuca sativa photosynthetic performance. Photosynthetica 2017, 55, 50–57. [Google Scholar]
- Zhang, K.; Yuan, J.; Kong, W.; Yang, Z. Genotype variations in cadmium and lead accumulations of leafy lettuce (Lactuca sativa L.) and screening for pollution-safe cultivars for food safety. Environ. Sci. Process. Impacts 2013, 15, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Silva, P.; Oliveira, H.; Gaivão, I.; Matos, M.; Pinto-Carnide, O.; Santos, C. Pb low doses induced genotoxicity in Lactuca sativa plants. Plant Physiol. Biochem. 2017, 112, 109–116. [Google Scholar] [CrossRef]
- Brengi, S.H.M.; Abouelsaad, I.A.A. The Combined Use of Beneficial Soil Microorganisms Enhanced the Growth and Efficiently Reduced Lead Content in Leaves of Lettuce (Lactuca sativa L.) Plant under Lead Stress. Alex. J. Agric. Sci. 2019, 64, 41–51. [Google Scholar]
- Garmash, E.V.; Golovko, T.K. CO2 gas exchange and growth in Rhaponticum carthamoides under the condition of middle taiga subzone of Northeastern Europe: Dependence of photosynthesis and respiration on environmental factors. Russ. J. Plant Physiol. 1997, 44, 737–745. [Google Scholar]
- Kok, B. A critical consideration of the quantum yield of Chlorella-photosynthesis. Enzymologia 1948, 13, 1–5. [Google Scholar]
- Farquhar, G.D.; von Caemmerer, S. Modelling of photosynthetic response to environmental conditions. In Physiological Plant Ecology II; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Encyclopedia of Plant Physiology, Volume 12B/B; Water Relations and Carbon Assimilation; Springer: Berlin/Heidelberg, Germany, 1982; Volume 12, pp. 549–587. [Google Scholar]
- González, L.; González-Vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Reigosa Roger, M.J., Ed.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2001; pp. 207–212. [Google Scholar]
- Ikkonen, E.N.; Grabelnykh, O.I.; Sherudilo, E.G.; Shibaeva, T.G. Salicylhydroxamic Acid-Resistant and Sensitive Components of Respiration in Chilling-Sensitive Plants Subjected to a Daily Short-Term Temperature Drop. Russ. J. Plant Physiol. 2020, 67, 60–67. [Google Scholar] [CrossRef]
- Campbell, C.; Atkinson, L.; Zaragoza-Castells, J.; Lundmark, M.; Atkin, O.; Hurry, V. Acclimation of photosynthesis and respiration is asynchronous in response to changes in temperature regardless of plant functional group. New Phytol. 2007, 176, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Çimrin, K.M.; Turan, M.; Kapur, B. Effect of elemental sulphur on heavy metals solubility and remediation by plants in calcareous soils. Fresenius Environ. Bull. 2007, 16, 1113–1120. [Google Scholar]
- Hung, N.M.; Hiep, N.V.; Dung, B.N.; Hai, N.X. Lead Accumulation in different parts of okra plant (Abelmoschus esculentus). ARPN J. Agric. Biol. Sci. 2014, 9, 190–194. [Google Scholar]
- Cai, X.; Jiang, M.; Liao, J.; Yang, Y.; Li, N.; Cheng, Q.; Li, X.; Song, H.; Luo, Z.; Liu, S. Biomass allocation strategies and Pb-enrichment characteristics of six dwarf bamboos under soil Pb stress. Ecotoxicol. Environ. Saf. 2021, 207, 111500. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.R.; McDonald, G.K.; Rengel, Z. Zn fertilization improves water use efficiency, grain yield and seed Zn content in chickpea. Plant Soil 2003, 249, 389–400. [Google Scholar] [CrossRef]
- Mahaffey, K.R. Environmental lead toxicity: Nutrition as a component of intervention. Environ. Health Perspect. 1990, 89, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.; Björkman, O. Quantum yields for CO2 uptake in C3 and C4 plants. Dependence on temperature, CO2 and O2 concentration. Plant Physiol. 1977, 59, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, E.S.; Krauss, K.W. Photosynthetic light response of flooded cherrybark oak (Quercus pagoda) seedlings grown in two light regimes. Tree Physiol. 2001, 21, 1103–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savitch, L.V.; Leonardos, E.D.; Krol, M.; Jansson, S.; Grodzinski, B.N.; Huner, P.A.; Öquist, G. Two different strategies for light utilization in photosynthesis in relation to growth and cold acclimation. Plant Cell Environ. 2002, 25, 761–771. [Google Scholar] [CrossRef]
- Sofronova, V.E.; Dymova, O.V.; Golovko, T.K.; Chepalov, V.A.; Petrov, K.A. Adaptive changes in pigment complex of Pinus sylvestris needles upon cold acclimation. Russ. J. Plant Physiol. 2016, 63, 433–442. [Google Scholar] [CrossRef]
- Ahmed, Z.F.R.; Alnuaimi, A.K.H.; Askri, A.; Tzortzakis, N. Evaluation of Lettuce (Lactuca sativa L.) Production under Hydroponic System: Nutrient Solution Derived from Fish Waste vs. Inorganic Nutrient Solution. Horticulturae 2021, 7, 292. [Google Scholar] [CrossRef]
- Opeolu, B.O.; Adenuga, P.A.; Ndakidemi Olujimi, O.O. Assessment of phyto–toxicity potential of lead on tomato (Lycopersicon esculentum L) planted on contaminated soils. Int. J. Nat. Sci. 2010, 5, 68–73. [Google Scholar]
- Ahmed, A.; Tajmir–Riahi, H.A. Interaction of toxic metal ions Cd2+, Hg2+ and Pb with light–harvesting proteins of chloroplast thylakoid membranes. An FTIR spectroscopic study. J. Inorg. Biochem. 1993, 50, 235–243. [Google Scholar] [CrossRef]
- Nwugo, C.C.; Huerta, A.J. Efects of silicon nutrition on cadmium uptake, growth and photosynthesis of rice plants exposed to lowlevel cadmium. Plant Soil 2008, 311, 73–86. [Google Scholar] [CrossRef]
- Tränkner, M.; Jákli, B.; Tavakol, E.; Geilfus, C.-M.; Cakmak, I.; Dittert, K.; Senbayram, M. Magnesium deficiency decreases biomass water-use efficiency and increases leaf water-use efficiency and oxidative stress in barley plants. Plant Soil 2016, 406, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Bruhn, D.; Hurry, V.M.; Tjoelker, M.G. The hot and the cold: Unraveling the variable response of plant respiration to temperature. Funct. Plant Biol. 2005, 32, 87–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semikhatova, O.A. Evaluation of plant adaptation potential by assessing dark respiration. Russ. J. Plant Physiol. 1998, 45, 122–128. [Google Scholar]
- Ikkonen, E.N.; Shibaeva, T.K.; Titov, A.F. Influence of Daily Short-Term Temperature Drops on Respiration to Photosynthesis Ratio in Chilling-Sensitive Plants. Russ. J. Plant Physiol. 2018, 65, 94–99. [Google Scholar] [CrossRef]
- Ayub, G.; Zaragoza-Castells, J.; Griffin, K.L.; Atkin, O.K. Leaf respiration in darkness and in the light under pre-industrial, current and elevated atmospheric CO2 concentrations. Plant Sci. 2014, 226, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Crous, K.Y.; Wallin, G.; Atkin, O.K.; Uddling, J.; Ekenstam, A. Acclimation of light and dark respiration to experimental and seasonal warming are mediated by changes in leaf nitrogen in Eucalyptus globules. Tree Physiol. 2017, 37, 1069–1083. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, M.H.; Ogaya, R.; Barbeta, A.; Penuelas, J.; Zaragoza-Castells, J.; Atkin, O.K.; Valladares, F.; Gimeno, T.E.; Pias, B.; Griffin, K.L. Light inhibition of foliar respiration in response to soil water availability and seasonal changes in temperature in Mediterranean holm oak (Quercus ilex) forest. Funct. Plant Biol. 2017, 44, 1178–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heskel, M.A.; Tang, J. Environmental controls onlight inhibition of respiration and leaf and canopy daytimecarbon exchange in a temperate deciduous forest. Tree Physiol. 2018, 38, 1886–1902. [Google Scholar] [CrossRef] [Green Version]
- Ikkonen, E.N.; Shibaeva, T.G.; Sherudilo, E.G.; Titov, A.F. Response of winter wheat seedlings respiration to long-term cold exposure and short-term daily temperature drops. Russ. J. Plant Physiol. 2020, 67, 538–544. [Google Scholar] [CrossRef]
- Ikkonen, E.N.; Shibaeva, T.G.; Sherudilo, E.G.; Titov, A.F. Effect of Continuous Lighting on Mitochondrial Respiration in Solanacea Plants. Russ. J. Plant Physiol. 2022, in press.
- Heskel, M.A.; Atkin, O.K.; Turnbull, M.H.; Griffin, K.L. Bringing the Kok effect to light: A review on the integration of daytime respiration and net ecosystem exchange. Ecosphere 2013, 4, 1–14. [Google Scholar] [CrossRef]
- Griffin, K.L.; Turnbull, M.H. Light saturated RuBP oxygenation by Rubisco is a robust predictor of light inhibition of respiration in Triticum aestivum L. Plant Biol. 2013, 15, 755–775. [Google Scholar] [CrossRef] [PubMed]
- Tcherkez, G.; Gauthier, P.; Buckley, T.N.; Busch, F.A.; Barbour, M.M.; Bruhn, D.; Heskel, M.A.; Gong, X.Y.; Crous, K.Y.; Griffin, K.; et al. Leaf day respiration: Low CO2 flux but high significance for metabolism and carbon balance. New Phytol. 2017, 216, 986–1001. [Google Scholar] [CrossRef]
- Atkin, O.K.; Scheurwater, I.; Pons, T.L. Respiration as a percentage of daily photosynthesis in whole plants is homeostatic at moderate, but not high growth temperatures. New Phytol. 2007, 174, 367–380. [Google Scholar] [CrossRef]
- Yamori, W.; Noguchi, K.; Hikosaka, K.; Terashima, I. Cold tolerant crop species have greater temperature homeostasis of leaf respiration and photosynthesis than cold-sensitive species. Plant Cell Physiol. 2009, 50, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Rakhmankulova, Z.F. Physiological aspects of photosynthesis–respiration interrelation. Russ. J. Plant Physiol. 2019, 66, 365–374. [Google Scholar] [CrossRef]
- Atkin, O.K.; Scheurwater, I.; Pons, T.L. High thermal acclimation potential of both photosynthesis and respiration in two lowland Plantago species in contrast to an alpine congeneric. Glob. Chang. Biol. 2006, 12, 500–515. [Google Scholar] [CrossRef]
- Ayub, G.; Smith, R.A.; Tissue, D.T.; Atkin, O.K. Impacts of drought on leaf respiration in darkness and light in Eucalyptus saligna exposed to industrial-age atmospheric CO2 and growth temperature. New Phytol. 2011, 190, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Seregin, I.; Ivanov, V. Physiological aspects of cadmium and lead toxic effects on higher plants. Russ. J. Plant Physiol. 2001, 48, 606–630. [Google Scholar] [CrossRef]
- Kumar, G.; Singh, R.P.; Sushila, R. Nitrate assimilation and biomass production in Sesamum indicum L. cv-HT-1 seedlings in a Pb enriched environment. Water Air Soil Pollut. 1993, 66, 163–171. [Google Scholar] [CrossRef]
- Piechalak, A.; Tomaszewska, B.; Baralkiewicz, D.; Malecka, A. Accumulation and detoxification of lead ions in legumes. Phytochemistry 2002, 60, 153–162. [Google Scholar] [CrossRef]
- Feng, H.; Qian, Y.; Gallagher, F.; Wu, M.; Zhang, W.; Yu, L.; Zhu, Q.; Zhang, K.; Liu, C.-J.; Tappero, R. Lead accumulation and association with Fe on Typha latifolia root from an urban brownfield site. Environ. Sci. Pollut. Res. Int. 2013, 20, 3743–3750. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 0 Pb | 50 Pb | 250 Pb | p |
|---|---|---|---|---|
| LMA, g m−2 | 19.3 ± 0.8 b | 23.5 ± 0.6 a | 22.9 ± 0.7 a | <0.05 * |
| Tr, mmol m−2 s−1 | 1.06 ± 0.09 a | 0.93 ± 0.10 a | 0.94 ± 0.13 a | 0.631 ns |
| qs, mmol m−2 s−1 | 81 ± 9 a | 69 ± 7 a | 89 ± 10 a | 0.532 ns |
| An1200[area], μmol m−2 s−1 | 7.1 ± 0.6 a | 5.8 ± 0.6 ab | 4.9 ± 0.9 b | <0.05 * |
| An1200[mass], nmol g−1DW s−1 | 367 ± 28 a | 245 ± 26 b | 231 ± 41 b | <0.05 * |
| J, μmol m−2 s−1 | 47 ± 4 a | 46 ± 4 a | 43 ± 6 a | 0.738 ns |
| νo, μmol m−2 s−1 | 1.9 ± 0.3 a | 2.1 ± 0.1 a | 1.9 ± 0.3 a | 0.797 ns |
| νc, μmol m−2 s−1 | 8.9 ± 0.6 a | 8.7 ± 0.7 a | 8.1 ± 1.0 a | 0.744 ns |
| Ci:Ca | 0.59 ± 0.04 a | 0.61 ± 0.02 a | 0.62 ± 0.07 a | 0.807 ns |
| PWUE, μmolCO2 mmol−1H2O | 6.8 ± 0.7 a | 6.4 ± 0.6 a | 6.4 ± 0.9 a | 0.762 ns |
| α, μmolCO2 μmol−1 quant | 0.018 ± 0.001 a | 0.016 ± 0.001 ab | 0.014 ± 0.002 b | <0.05 * |
| RWC, % | 71 ± 2 ab | 69 ± 2 b | 76 ± 1 a | <0.05 * |
| LCP, μmol m−2 s−1 | 33 ± 5 c | 54 ± 6 b | 78 ± 15 a | <0.01 ** |
| Rl[area], μmol m−2 s−1 | 0.58 ± 0.08 b | 0.86 ± 0.08 ab | 1.00 ± 0.17 a | <0.01 ** |
| Rl[mass], nmol g−1DW s−1 | 30 ± 4 b | 37 ± 3 ab | 43 ± 6 a | <0.01 ** |
| Rd[area], μmol m−2 s−1 | 0.71 ± 0.07 b | 0.99 ± 0.08 ab | 1.14 ± 0.18 a | <0.01 ** |
| Rd[mass], nmol g−1DW s−1 | 37 ± 4 b | 42 ± 3 ab | 50 ± 4 a | <0.01 ** |
| Rl:Rd | 0.81 ± 0.04 a | 0.87 ± 0.014 a | 0.87 ± 0.015 a | 0.959 ns |
| Ag[area], μmol m−2 s−1 | 7.6 ± 0.5 a | 6.6 ± 0.5 ab | 5.9 ± 0.8 b | <0.05 * |
| Rl:Ag | 0.07 ± 0.01 c | 0.14 ± 0.02 b | 0.20 ± 0.05 a | <0.001 *** |
| Rd:Ag | 0.09 ± 0.01 c | 0.16 ± 0.02 b | 0.22 ± 0.06 a | <0.001 *** |
| Vt, μmolO2 g−1DW h−1 | 100 ± 15 a | 106 ± 9 a | 96 ± 7 a | 0.987 ns |
| VSHAM-res, μmolO2 g−1DW h−1 | 45 ± 9 a | 45 ± 7 a | 47 ± 4 a | 0.998 ns |
| VSHAM-sen:Vt | 51 ± 7 a | 55 ± 9 a | 51 ± 3 a | 0.968 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikkonen, E.; Kaznina, N. Physiological Responses of Lettuce (Lactuca sativa L.) to Soil Contamination with Pb. Horticulturae 2022, 8, 951. https://doi.org/10.3390/horticulturae8100951
Ikkonen E, Kaznina N. Physiological Responses of Lettuce (Lactuca sativa L.) to Soil Contamination with Pb. Horticulturae. 2022; 8(10):951. https://doi.org/10.3390/horticulturae8100951
Chicago/Turabian StyleIkkonen, Elena, and Natalia Kaznina. 2022. "Physiological Responses of Lettuce (Lactuca sativa L.) to Soil Contamination with Pb" Horticulturae 8, no. 10: 951. https://doi.org/10.3390/horticulturae8100951
APA StyleIkkonen, E., & Kaznina, N. (2022). Physiological Responses of Lettuce (Lactuca sativa L.) to Soil Contamination with Pb. Horticulturae, 8(10), 951. https://doi.org/10.3390/horticulturae8100951