
Monitoring Stem Water Potential with an Embedded Microtensiometer to Inform Irrigation Scheduling in Fruit Crops
{kind=link}
{kind=link}
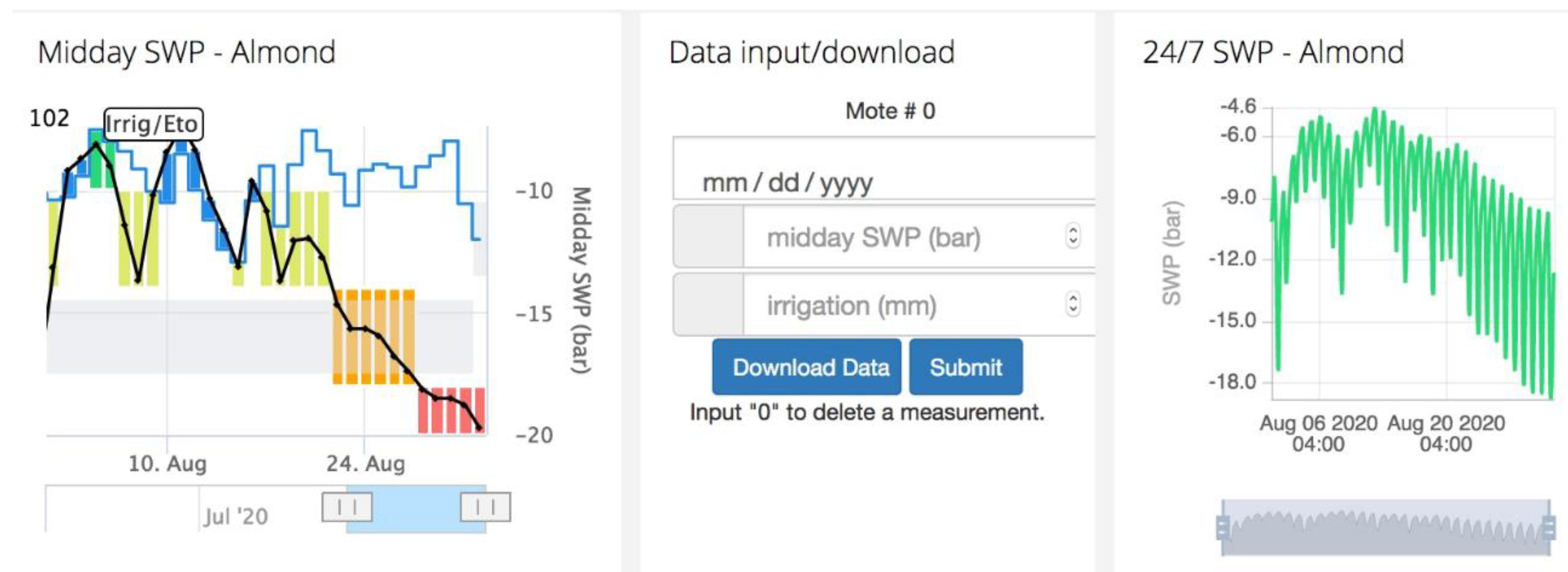{kind=link}
{kind=link}
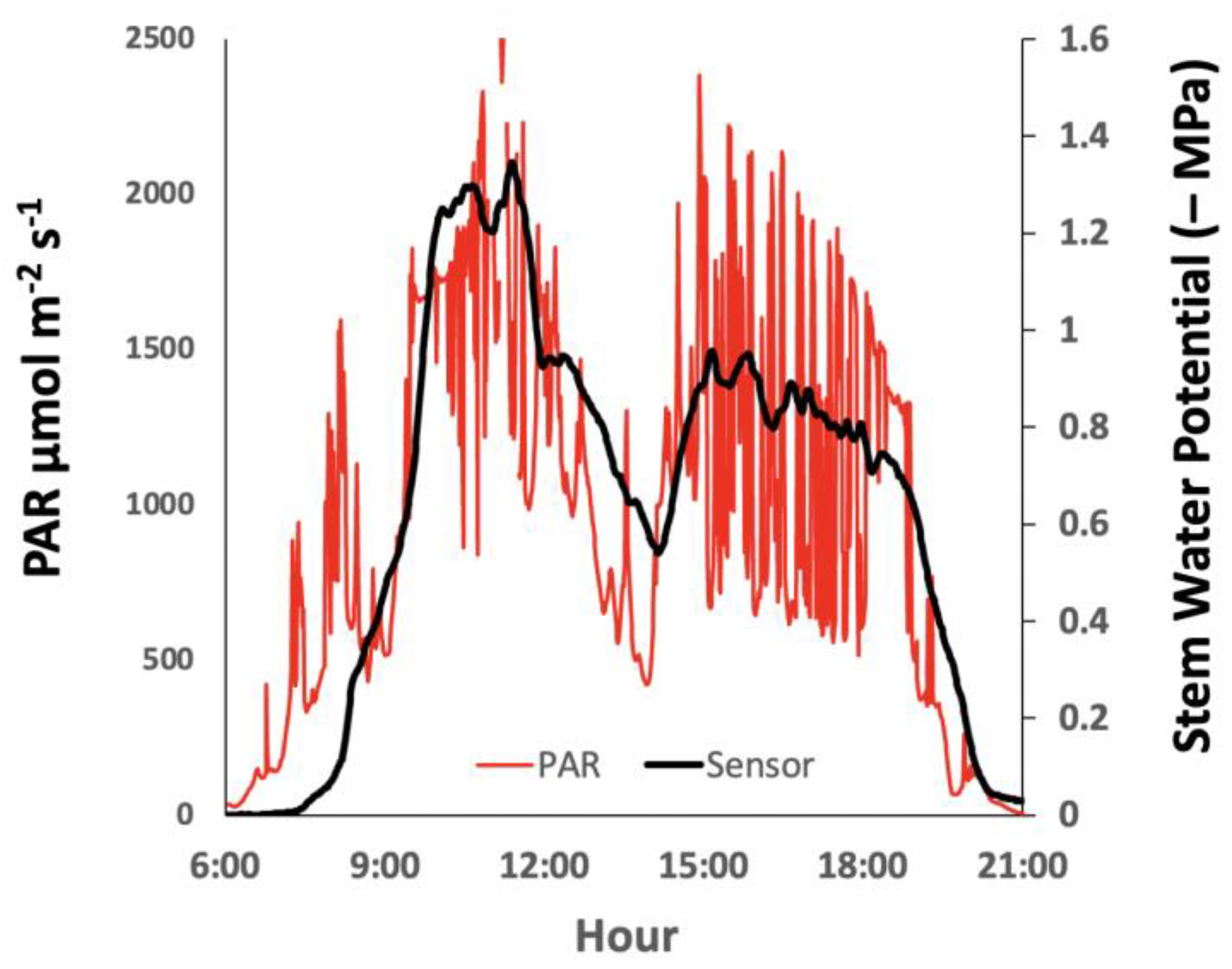{kind=link}
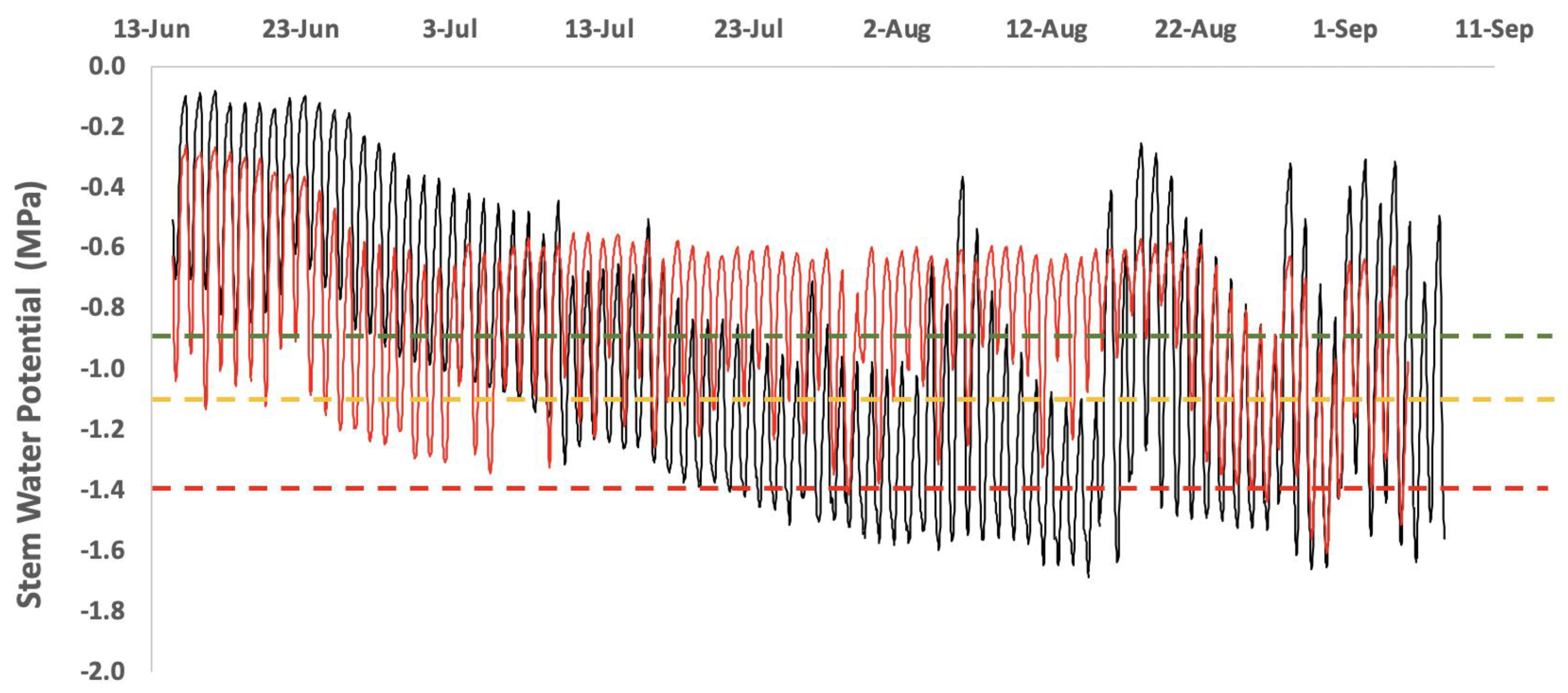{kind=link}
{kind=link}
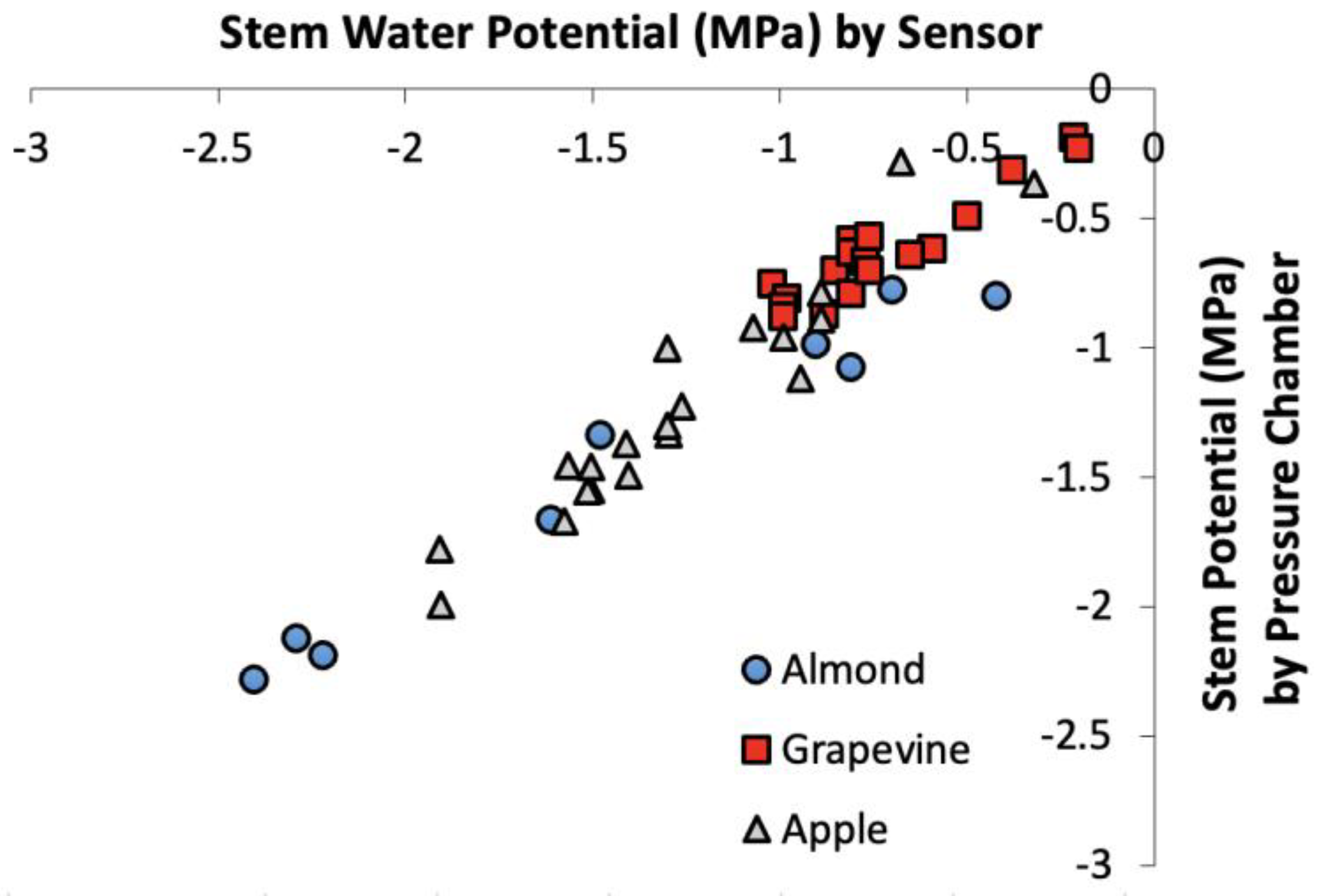{kind=link}
{kind=link}
Abstract
:1. Introduction

Plant Measurements to Estimate or Measure Water Status
- capable of direct measurement of plant water potential;
- accurate (0.01 MPa or better) and stable over the full physiologically relevant range of plant water potential (0 to −3 MPa or lower);
- produces continuous, real-time measurements over months to years;
- simple to install and operate with minimal sensitivity to temperature variations and contaminants;
- as small as possible to allow precise spatial measurements and to minimize disruption of tissue upon embedding;
- reasonable cost to manufacture and deploy; many sensors may be needed to resolve spatial variations;
- compatible with wireless networks for real-time data collection and spatial integration.
2. Materials and Methods
3. Results
4. Discussion
General Limitations of Plant Sensors
5. Conclusions and New Opportunities
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fereres, E.; Goldhamer, D.A. Deciduous fruit and nut trees. In Irrigation of Agricultural Crops; Stewart, B.A., Nielsen, D.R., Eds.; ASA-CSSA-SSSA: Madison, WI, USA, 1990; pp. 987–1017. [Google Scholar]
- Williams, L.E.; Matthews, M.A. Grapevine. In Irrigation of Agricultural Crops; Stewart, B.A., Nielsen, D.R., Eds.; ASA-CSSA-SSSA: Madison, WI, USA, 1990; pp. 1019–1055. [Google Scholar]
- Lakso, A.N. Water relations of apples. In Apples, Botany, Production and Uses; Ferree, D.C., Warrington, I.A., Eds.; CAB International: Wallingford, UK, 2003; pp. 167–194. [Google Scholar] [CrossRef]
- Powell, D.B.B. Some effects of water stress on the growth and development of apple trees. J. Hortic. Sci. 1976, 51, 75–90. [Google Scholar] [CrossRef]
- Guilpart, N.; Roux, S.; Gary, C.; Metay, A. The trade-off between grape yield and grapevine susceptibility to powdery mildew and grey mould depends on inter-annual variations in water stress. Agric. Forest Meteorol. 2017, 234, 203–211. [Google Scholar] [CrossRef]
- Atkinson, D. The distribution and effectiveness of the roots of tree crops. Hortic. Rev. 1980, 2, 424–490. [Google Scholar]
- Smart, D.R.; Schwass, E.; Shaulis, N.J.; Lakso, A.N.; Morano, L. Grapevine rooting patterns: A comprehensive analysis and a review. Amer. J. Enol. Vitic. 2006, 57, 89–104. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Vanden Heuvel, J.E. Influence of grapevine training systems on vine growth and fruit composition: A review. Amer. J. Enol. Vitic. 2009, 60, 251–268. [Google Scholar] [CrossRef]
- Musacchi, S.; Greene, D. Innovations in apple tree cultivation to manage crop load and ripening. In Achieving Sustainable Cultivation of Apples; Evans, K., Ed.; Burleigh Dodds Science Publ.: Cambridge, UK, 2017; pp. 195–237. [Google Scholar] [CrossRef]
- Fazio, G. Genetics, Breeding, and Genomics of Apple Rootstocks. In The Apple Genome; Korban, S.S., Ed.; Compendium of Plant Genomes; Springer: Cham, Switzerland, 2021; pp. 105–130. [Google Scholar] [CrossRef]
- Naor, A. Irrigation scheduling and evaluation of tree water status in deciduous orchards. Hortic. Rev. 2006, 32, 111–165. [Google Scholar] [CrossRef]
- Jones, H.G.; Lakso, A.N.; Syvertsen, J.P. Physiological control of water status in Temperate and subtropical fruit trees. Hortic. Rev. 1985, 7, 301–344. [Google Scholar]
- Jones, H.G. Plants and Microclimate—A Quantitative Approach to Environmental Plant Physiology, 3rd ed.; Cambridge Univ. Press: Cambridge, UK, 2014. [Google Scholar]
- Lakso, A.N.; Intrigliolo, D.S. Plant-based sensing for irrigation management in the field. Acta Hortic. 2022, 1335, 247–262. [Google Scholar] [CrossRef]
- Scholasch, T. Improving winegrowing with sap flow driven irrigation—A 10-year review. Acta Hortic. 2018, 1222, 155–168. [Google Scholar] [CrossRef]
- Vandegehuchte, M.W.; Steppe, K. Sap-flux density measurement methods: Working principles and applicability. Funct. Plant Biol. 2013, 40, 213–223. [Google Scholar] [CrossRef]
- McElrone, A.J.; Shapland, T.M.; Calderon, A.; Fitzmaurice, L.; Paw, U.K.T.; Snyder, R.L. Surface Renewal: An advanced micrometeorological method for measuring and processing field-scale energy flux density data. J. Visualized Exp. 2013, 82, e50666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tule Technologies Website. Available online: https://tule.ag/ (accessed on 17 October 2022).
- Basile, B.; Girona, J.; Behboudian, M.H.; Mata, M.; Rosello, J.; Ferre, M.; Marsal, J. Responses of “Chardonnay” to deficit irrigation applied at different phenological stages: Vine growth, must composition, and wine quality. Irrig. Sci. 2012, 30, 397–406. [Google Scholar] [CrossRef]
- López-López, M.; Espadafor, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Water use of irrigated almond trees when subjected to water deficits. Agric. Water Manage 2018, 195, 84–93. [Google Scholar] [CrossRef]
- Galindo, A.; Collado-González, J.; Griñán, I.; Corell, M.; Centeno, A.; Martín-Palomo, M.J.; Girón, I.F.; Rodríguez, P.; Cruz, Z.N.; Memmi, H.; et al. Deficit irrigation and emerging fruit crops as a strategy to save water in Mediterranean semiarid agrosystems. Agric. Water Manag. 2018, 202, 311–324. [Google Scholar] [CrossRef]
- Scholasch, T.; Rienth, M. Review of water deficit mediated changes in vine and berry physiology; Consequences for the optimization of irrigation strategies. Oeno One 2019, 3, 423–444. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exper. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Lakso, A.N.; Corelli Grappadelli, L.; Barnard, J.; Goffinet, M.C. An expolinear model of the growth pattern of the apple fruit. J. Hortic. Sci. 1995, 70, 389–394. [Google Scholar] [CrossRef]
- Famiani, F.; Bonghi, C.; Chen, Z.-H.; Drincovich, M.F.; Farinelli, D.; Lara, M.V.; Proietti, S.; Rosati, A.; Vizzotto, G.; Walker, R.P. Stone fruits: Growth and nitrogen and organic acid metabolism in the fruits and seeds—A review. Front. Plant Sci. 2020, 11, 572601. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Barbe, J.-C.; Darriet, P.; Geffroy, O.; Gomès, E.; Guillaumie, S.; Helwi, P.; Laboyrie, J.; Lytra, G.; Le Menn, N.; et al. Recent advancements in understanding the terroir effect on aromas in grapes and wines. Oeno One 2020, 4, 985–1006. [Google Scholar] [CrossRef]
- Rienth, M.; Vigneron, N.; Darriet, P.; Sweetman, C.; Burbidge, C.; Bonghi, C.; Walker, R.P.; Famiani, F.; Castellarin, S.D. Grape berry secondary metabolites and their modulation by abiotic factors in a climate change scenario—A review. Front. Plant Sci. 2021, 12, 643258. [Google Scholar] [CrossRef]
- Baram, S.; Couvreur, V.; Harter, T.; Read, M.; Brown, P.H.; Hopmans, J.W.; Smart, D.R. Assessment of orchard N losses to groundwater with a vadose zone monitoring network. Agric. Water Man. 2016, 172, 83–95. [Google Scholar] [CrossRef]
- Gautam, D.; Pagay, V. A Review of current and potential applications of remote sensing to study the water status of horticultural crops. Agronomy 2020, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Tosin, R.; Martins, R.; Pocas, I.; Cunha, M. Canopy VIS-NIR spectroscopy and self-learning artificial intelligence for a generalised model of predawn leaf water potential in Vitis vinifera. Biosyst. Engineering 2022, 219, 235–258. [Google Scholar] [CrossRef]
- Girón, I.F.; Corell, M.; Martín-Palomo, M.J.; Galindo, A.; Torrecillas, A.; Moreno, F.; Moriana, A. Limitations and usefulness of maximum daily shrinkage (MDS) and trunk growth rate (TGR) indicators in the irrigation scheduling of table olive trees. Agric. Water Manage. 2016, 164, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Scalisi, A.; Bresilla, K.; Simões Grilo, F. Continuous determination of fruit tree water-status by plant-based sensors. Italus Hortus 2017, 24, 39–50. [Google Scholar] [CrossRef]
- Scholander, P.F.; Hammel, H.T.; Hemingsen, E.A.; Bradstreet, E.D. Hydrostatic pressure and osmotic potential of leaves of mangrove and some other plants. Proc. Natl. Acad. Sci. USA 1964, 52, 119. [Google Scholar] [CrossRef] [Green Version]
- Scholander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap pressure in vascular plants. Science 1965, 148, 339. [Google Scholar] [CrossRef]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Shackel, K. A plant-based approach to deficit irrigation in trees and vines. HortScience 2011, 46, 173–177. [Google Scholar] [CrossRef]
- Patakas, A.; Noitsakis, B.; Chouzouri, A. Optimisation of irrigation water use in grapevines using the relationship between transpiration and plant water status. Agric. Ecosyst. Environ. 2005, 106, 253–259. [Google Scholar] [CrossRef]
- Naor, A. Midday stem water potential as a plant water stress indicator for irrigation scheduling in fruit trees. Acta Hortic. 2000, 537, 447–454. [Google Scholar] [CrossRef]
- Chone, X.; Van Leeuwen, C.; Dubordieu, D.; Gaudillere, J.P. Stem water potential is a sensitive integrator of grapevine water status. Ann. Botany 2001, 87, 477–483. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, C.; Pieri, P.; Vivin, P. Comparison of three operational tools for the assessment of vine water status, stem water potential, carbon isotope discrimination measured on grape sugar and water balance. In Methodologies and Results in Grapevine Research; Delrot, S., Medrano, H., Or, E., Bavaresco, L., Grando, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 87–106. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Miranda, C.; Marín, D.; Sesma, B.; Intrigliolo, D.S.; Mirás-Avalos, J.M.; Escalona, J.M.; Montoro, A.; de Herralde, F.; Baeza, P.; et al. Discrimination ability of leaf and stem water potential at different times of the day through a meta-analysis in grapevine (Vitis vinifera L.). Agric. Water Manag. 2019, 221, 202–210. [Google Scholar] [CrossRef]
- Tian, T.; Schreiner, R.P. Appropriate time to measure leaf and stem water potential in north-south oriented, vertically shoot-positioned vineyards. Amer. J. Enol. Vitic. 2021, 72, 64–72. [Google Scholar] [CrossRef]
- Fernández, J.E. Plant-based sensing to monitor water stress, applicability to commercial orchards. Agric. Water Manage. 2014, 142, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Pagay, V.; Santiago, M.; Sessoms, D.A.; Huber, E.J.; Vincent, O.; Pharkya, A.; Corso, T.N.; Lakso, A.N.; Stroock, A.D. A microtensiometer capable of measuring water potentials below -10 MPa. Lab on a Chip 2014, 14, 2806–2817. [Google Scholar] [CrossRef]
- Black, W.L.; Santiago, M.; Zhu, S.; Stroock, A.D. Ex situ and in situ measurement of water activity with a mems tensiometer. Analytical Chem. 2019, 92, 716–723. [Google Scholar] [CrossRef]
- Blanco, V.; Kalcsits, L. Microtensiometers accurately measure stem water potential in woody perennials. Plants 2021, 10, 2780. [Google Scholar] [CrossRef]
- Lakso, A.N.; Zhu, S.; Santiago, S.; Shackel, K.; Volkov, V.; Stroock, A.D. A microtensiometer sensor to continuously monitor stem water potentials in woody plants - design and field testing. Acta Hortic. 2022, 1335, 317–324. [Google Scholar] [CrossRef]
- Pagay, V. Evaluating a novel microtensiometer for continuous trunk water potential measurements in field-grown irrigated grapevines. Irrig. Sci. 2022, 40, 45–54. [Google Scholar] [CrossRef]
- Olien, W.C.; Lakso, A.N. Effect of rootstock on apple (Malus domestica) tree water relations. Physiol. Plant. 1986, 67, 421–430. [Google Scholar] [CrossRef]
- Forster, M.A. How significant is nocturnal sap flow? Tree Physiol. 2014, 34, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameglio, T.; Archer, P.; Cohen, M.; Valancogne, C.; Daudet, F.-A.; Dayau, S.; Cruiziat, P. Significance and limits in the use of predawn leaf water potential for tree irrigation. Plant Soil 1999, 207, 155–167. [Google Scholar] [CrossRef]
- Shackel, K.; Moriana, A.; Marino, G.; Corell, M.; Pérez-López, D.; Martin-Palomo, M.J.; Caruso, T.; Marra, F.P.; Agüero Alcaras, L.M.; Milliron, L.; et al. Establishing a reference baseline for midday stem water potential in olive and its use for plant-based irrigation management. Front. Plant Sci. 2021, 12, 791711. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, C.R.; McElrone, A.J.; Choat, B.; Lee, E.F.; Shackel, K.A.; Matthews, M.A. In vivo visualizations of drought-induced embolism spread in Vitis vinifera. Plant Physiol. 2013, 161, 1820–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landsberg, J.J.; Beadle, C.L.; Biscoe, P.V.; Butler, D.R.; Davidson, B.; Incoll, L.D.; James, D.B.; Jarvis, P.G.; Martin, P.J.; Neilson, R.E.; et al. Diurnal energy, water and CO2 exchanges in an apple (Malus pumila) orchard. J. Appl. Ecol. 1975, 12, 659–684. [Google Scholar] [CrossRef]
- Levin, A.D. Re-evaluating pressure chamber methods of water status determination in field-grown grapevine (Vitis spp.). Agric. Water Manag. 2019, 221, 422–429. [Google Scholar] [CrossRef]
- Damm, A.; Paul-Limoges, E.; Haghighi, E.; Simmer, C.; Morsdorf, F.; Schneider, F.D.; van der Tol, C.; Migliavacca, M.; Rascher, U. Remote sensing of plant-water relations: An overview and future perspectives. J. Plant Physiol. 2018, 227, 3–19. [Google Scholar] [CrossRef]
- Yu, R.; Zaccaria, D.; Kisekka, I.; Kurtural, S.K. Soil apparent electrical conductivity and must carbon isotope ratio provide indication of plant water status in wine grape vineyards. Prec. Agric. 2021, 22, 1333–1352. [Google Scholar] [CrossRef]
- Brillante, L.; Martínez-Lüscher, J.; Yu, R.; Kurtural, S.K. Carbon isotope discrimination (δ13C) of grape musts is a reliable tool for zoning and the physiological ground-truthing of sensor maps in precision viticulture. Front. Environ. Sci. 2020, 8, 561477. [Google Scholar] [CrossRef]
- Rosenthal, R. The “File Drawer Problem” and Tolerance for Null Results. Psychol. Bull. 1979, 86, 638–641. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics, 2nd ed.; Springer: New York, NY, USA, 1998; 286p. [Google Scholar]
- Lakso, A.N. Comparing water use efficiency of apples and grapes-physiological and morphological aspects. Acta Hortic. 2014, 1038, 67–72. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Perulli, G.D.; Manfrini, L.; Zibordi, M.; Lopez Velasco, G.; Anconelli, S.; Pierpaoli, E.; Corelli Grappadelli, L.; Morandi, B. Time of irrigation affects vine water relations and the daily patterns of leaf gas exchanges and vascular flows to kiwifruit (Actinidia deliciosa Chev.). Agric. Water Manag. 2016, 166, 101–110. [Google Scholar] [CrossRef]
- Scalisi, A.; O’Connell, M.G.; Turpin, S.R.; Lo Bianco, R. Diurnal irrigation timing affects fruit growth in late-ripening nectarines. Acta Hortic. 2021, 1314, 61–68. [Google Scholar] [CrossRef]
- Lakso, A.N. Challenges to interpreting internal and external factors limiting apple fruit growth, set and abscission. Acta Hortic. 2022, 1342, 317–328. [Google Scholar] [CrossRef]
- Yu, R.; Brillante, L.; Torres, N.; Kurtural, S.K. Proximal sensing of vineyard soil and canopy vegetation for determining vineyard spatial variability in plant physiology and berry chemistry. Oeno One 2021, 55, 315–333. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lakso, A.N.; Santiago, M.; Stroock, A.D. Monitoring Stem Water Potential with an Embedded Microtensiometer to Inform Irrigation Scheduling in Fruit Crops. Horticulturae 2022, 8, 1207. https://doi.org/10.3390/horticulturae8121207
Lakso AN, Santiago M, Stroock AD. Monitoring Stem Water Potential with an Embedded Microtensiometer to Inform Irrigation Scheduling in Fruit Crops. Horticulturae. 2022; 8(12):1207. https://doi.org/10.3390/horticulturae8121207
Chicago/Turabian StyleLakso, Alan N., Michael Santiago, and Abraham D. Stroock. 2022. "Monitoring Stem Water Potential with an Embedded Microtensiometer to Inform Irrigation Scheduling in Fruit Crops" Horticulturae 8, no. 12: 1207. https://doi.org/10.3390/horticulturae8121207
APA StyleLakso, A. N., Santiago, M., & Stroock, A. D. (2022). Monitoring Stem Water Potential with an Embedded Microtensiometer to Inform Irrigation Scheduling in Fruit Crops. Horticulturae, 8(12), 1207. https://doi.org/10.3390/horticulturae8121207