
Transcriptional Effects of Rootstock on Scion after Drought: A Case Study of Using MdGH3 RNAi as the Rootstock

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction and Sequencing Procedures
2.3. RNA-Seq Analysis
2.4. Mobile mRNA Identification by RNA-Seq
3. Results
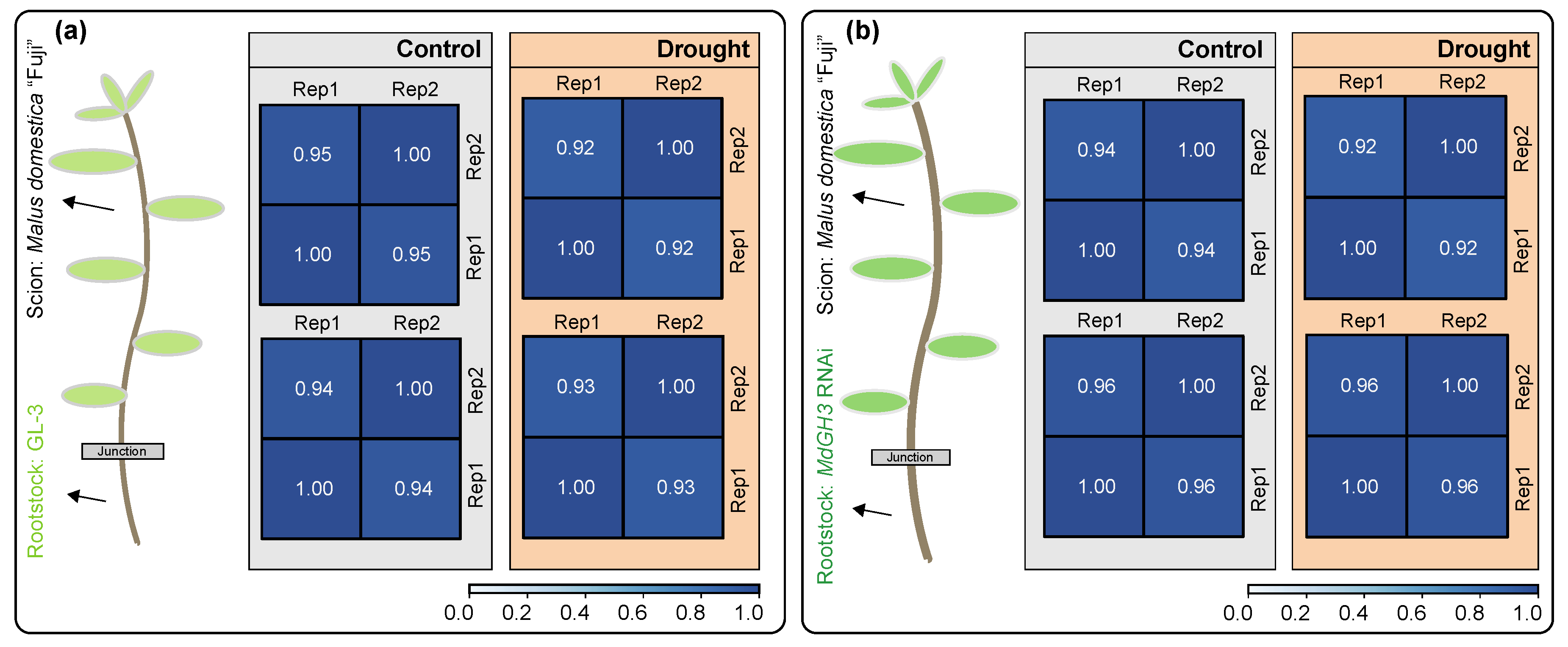
3.1. Phenotypic Comparison and Transcriptome of Grafted Plants Using MdGH3 RNAi and GL-3 Plants as Rootstocks
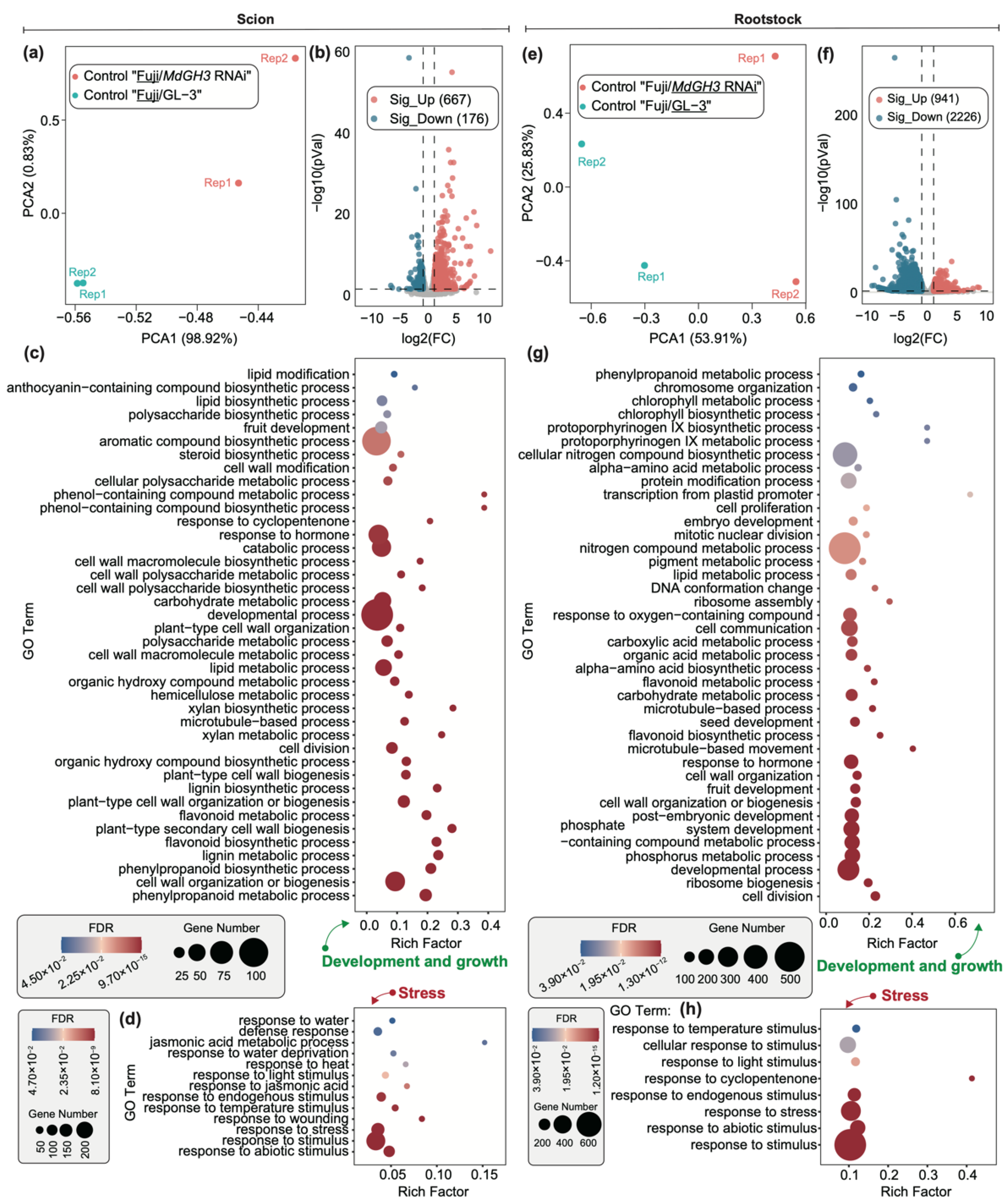
3.2. Differentially Expressed Genes (DEGs) in Scion and Rootstock under Control Conditions
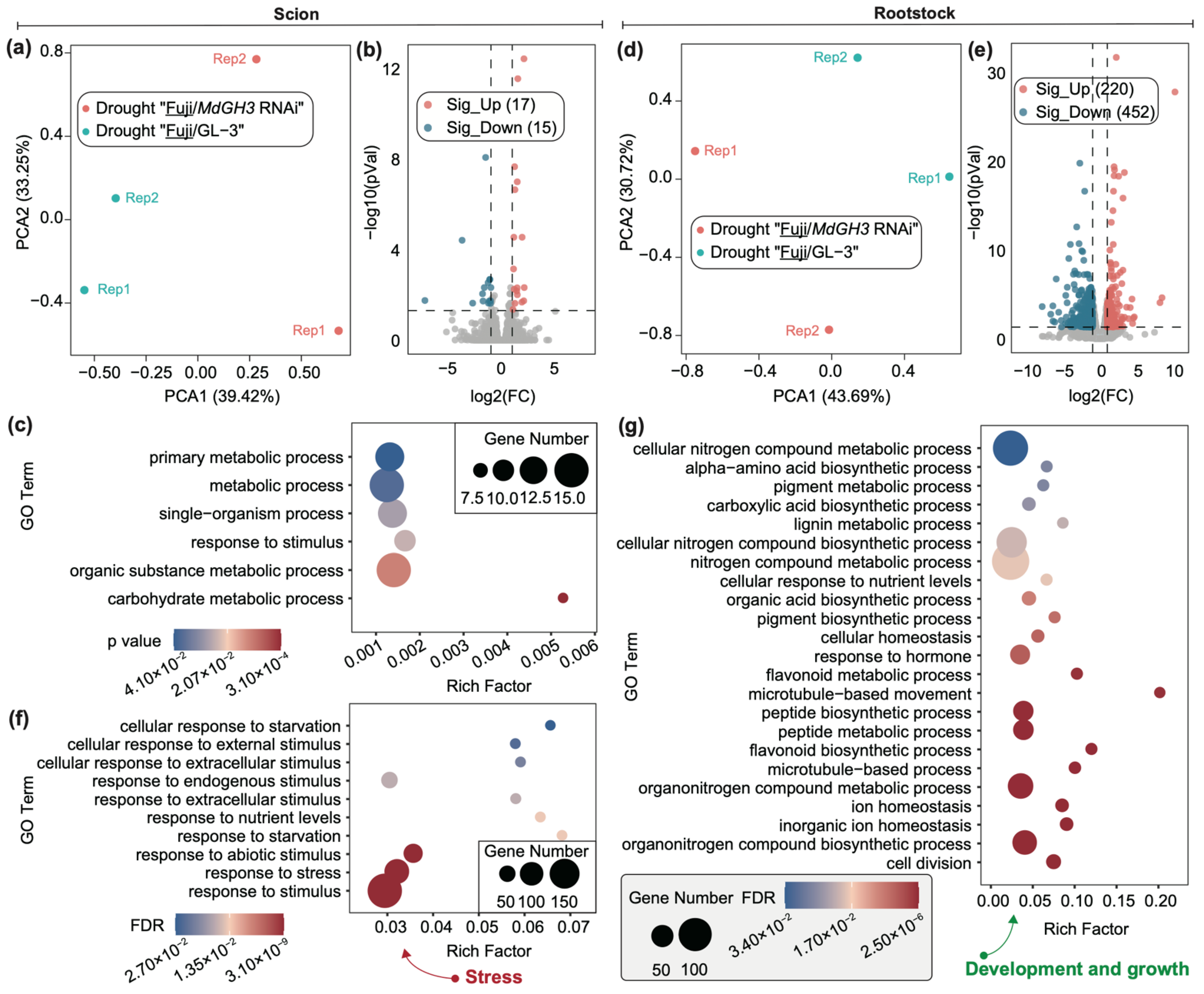
3.3. DEGs in Drought-Treated Scion and Rootstock
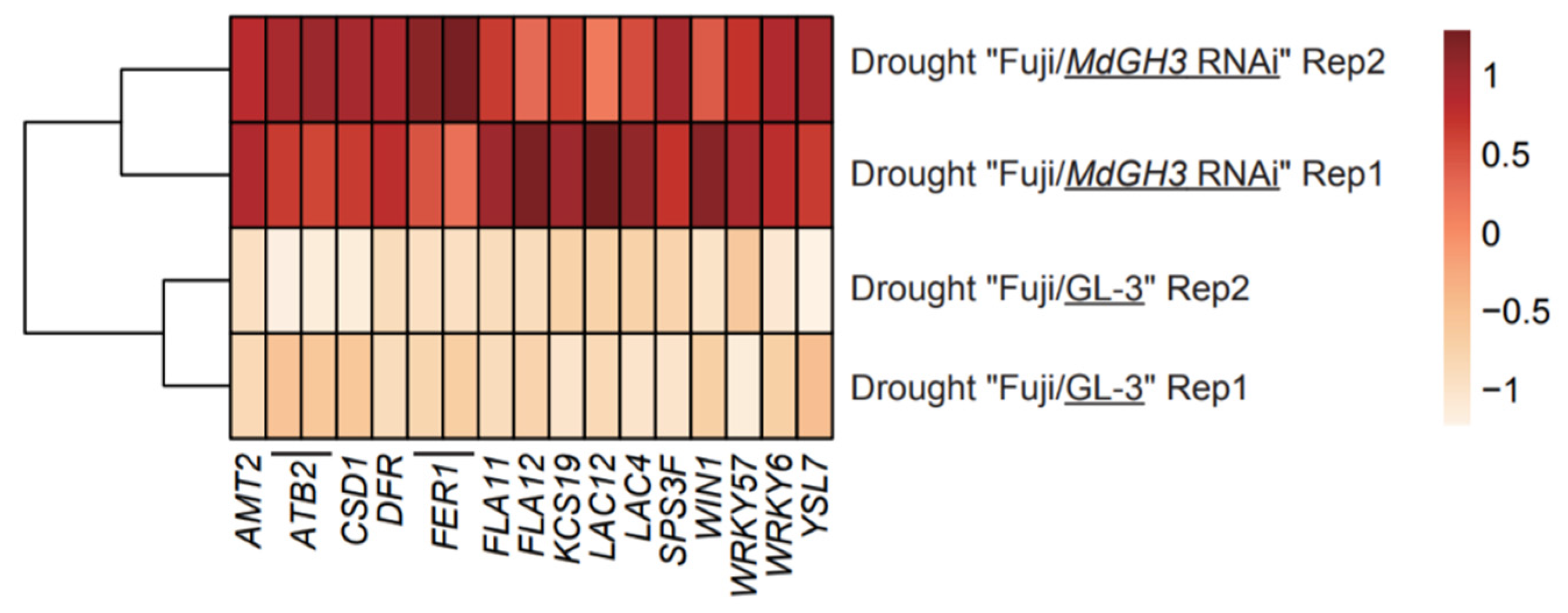
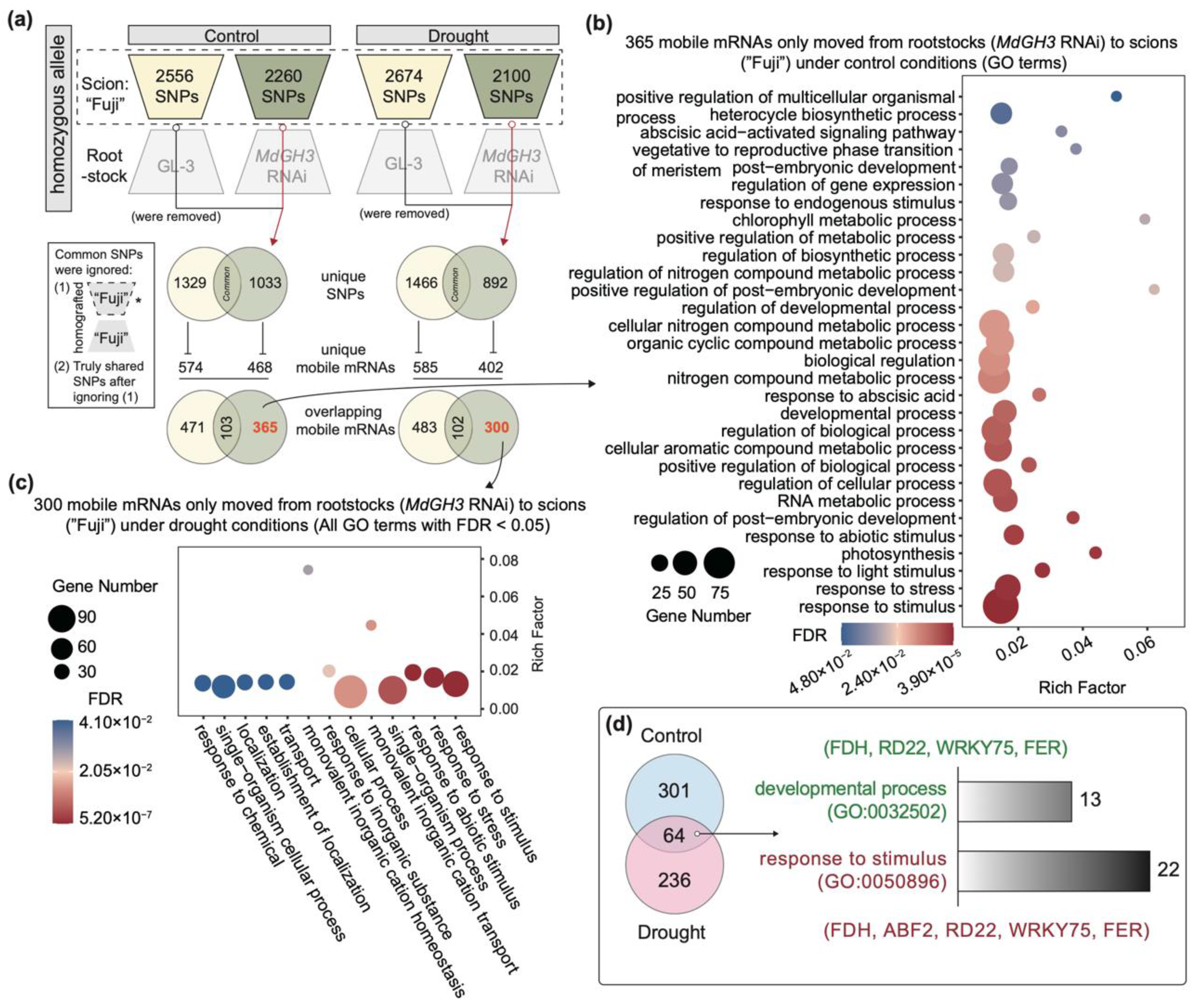
3.4. Identification of Mobile mRNAs Moving from MdGH3 RNAi Rootstock to Scion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Hou, N.; Liu, Z.; He, J. Profiling of N6-Methyladenosine (M6A) Modification Landscape in Response to Drought Stress in Apple (Malus Prunifolia (Willd.) Borkh). Plants 2021, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Tikkanen, M.; Grieco, M.; Aro, E.-M. Novel Insights into Plant Light-Harvesting Complex II Phosphorylation and ‘State Transitions’. Trends Plant Sci. 2011, 16, 126–131. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Allakhverdiev, S.I.; Murata, N. Protein Synthesis Is the Primary Target of Reactive Oxygen Species in the Photoinhibition of Photosystem II. Physiol. Plant. 2011, 142, 35–46. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of Photosynthesis during Abiotic Stress-Induced Photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Shigeoka, S. Understanding Oxidative Stress and Antioxidant Functions to Enhance Photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, L.; He, J.; Li, X.; Hou, N.; Guo, J.; Niu, C.; Li, C.; Liu, S.; Xu, J.; et al. Chromosome-scale Reference Genome Provides Insights into the Genetic Origin and Grafting-mediated Stress Tolerance of Malus Prunifolia. Plant Biotechnol. J. 2022, 20, 1015–1017. [Google Scholar] [CrossRef]
- Jiang, L.; Shen, W.; Liu, C.; Tahir, M.M.; Li, X.; Zhou, S.; Ma, F.; Guan, Q. Engineering Drought-Tolerant Apple by Knocking down Six GH3 Genes and Potential Application of Transgenic Apple as a Rootstock. Hortic. Res. 2022, 9, uhac122. [Google Scholar] [CrossRef]
- Tzfira, T. Agrobacterium T-DNA Integration: Molecules and Models. Trends Genet. 2004, 20, 375–383. [Google Scholar] [CrossRef]
- Terns, R.M.; Terns, M.P. CRISPR-Based Technologies: Prokaryotic Defense Weapons Repurposed. Trends Genet. 2014, 30, 111–118. [Google Scholar] [CrossRef]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuong, U.T.; Iswanto, A.B.B.; Nguyen, Q.; Kang, H.; Lee, J.; Moon, J.; Kim, S.H. Engineering Plant Immune Circuit: Walking to the Bright Future with a Novel Toolbox. Plant Biotechnol. J. 2022, pbi.13916. [Google Scholar] [CrossRef] [PubMed]
- Khawar, K.M.; Onarici, S.; Ozel, C.A.; Aasim, M.; Bakhsh, A.; Rao, A.Q. Plant Biotechnology. Sci. World J. 2013, 2013, 736731. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Pessina, S.; Lenzi, L.; Perazzolli, M.; Campa, M.; Dalla Costa, L.; Urso, S.; Valè, G.; Salamini, F.; Velasco, R.; Malnoy, M. Knockdown of MLO Genes Reduces Susceptibility to Powdery Mildew in Grapevine. Hortic. Res. 2016, 3, 16016. [Google Scholar] [CrossRef] [Green Version]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering Canker-Resistant Plants through CRISPR/Cas9-Targeted Editing of the Susceptibility Gene CsLOB1 Promoter in Citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [Green Version]
- Östin, A.; Kowalyczk, M.; Bhalerao, R.P.; Sandberg, G. Metabolism of Indole-3-Acetic Acid in Arabidopsis1. Plant Physiol. 1998, 118, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, M.; Sandberg, G. Quantitative Analysis of Indole-3-Acetic Acid Metabolites in Arabidopsis. Plant Physiol. 2001, 127, 1845–1853. [Google Scholar] [CrossRef]
- Brunoni, F.; Collani, S.; Casanova-Sáez, R.; Šimura, J.; Karady, M.; Schmid, M.; Ljung, K.; Bellini, C. Conifers Exhibit a Characteristic Inactivation of Auxin to Maintain Tissue Homeostasis. New Phytol. 2020, 226, 1753–1765. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Wu, N.; Fu, J.; Wang, S.; Li, X.; Xiao, J.; Xiong, L. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice. J. Exp. Bot. 2012, 63, 6467–6480. [Google Scholar] [CrossRef]
- Kirungu, J.N.; Magwanga, R.O.; Lu, P.; Cai, X.; Zhou, Z.; Wang, X.; Peng, R.; Wang, K.; Liu, F. Functional Characterization of Gh_A08G1120 (GH3.5) Gene Reveal Their Significant Role in Enhancing Drought and Salt Stress Tolerance in Cotton. BMC Genet 2019, 20, 62. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Zhao, K.; Lei, H.; Shen, X.; Liu, Y.; Liao, X.; Li, T. Genome-Wide Analysis of the GH3 Family in Apple (Malus × Domestica). BMC Genom. 2013, 14, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Wang, Y.; Feng, C.; Wei, Y.; Peng, X.; Guo, X.; Guo, X.; Zhai, Z.; Li, J.; Shen, X.; et al. Overexpression of MsGH3.5 Inhibits Shoot and Root Development through the Auxin and Cytokinin Pathways in Apple Plants. Plant J. 2020, 103, 166–183. [Google Scholar] [CrossRef]
- Harada, T. Grafting and RNA Transport via Phloem Tissue in Horticultural Plants. Sci. Hortic. 2010, 125, 545–550. [Google Scholar] [CrossRef]
- Kehr, J.; Kragler, F. Long Distance RNA Movement. New Phytol. 2018, 218, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H. RNA Transport: Delivering the Message. Nat. Plants 2015, 1, 15038. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, D.; Liu, C.; Shen, W.; He, J.; Yue, Q.; Niu, C.; Yang, F.; Li, X.; Shen, X.; et al. MdGH3.6 Is Targeted by MdMYB94 and Plays a Negative Role in Apple Water-deficit Stress Tolerance. Plant J. 2022, 109, 1271–1289. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, K.; Kan, Z.; Dang, H.; Feng, S.; Yang, Y.; Li, L.; Nou, H.; Xu, L.; Wang, X.; et al. The regulatory module MdBT2–MdMYB88/MdMYB124–MdNRTs regulates nitrogen usage in apple. Plant Physiol. 2021, 185, 1924–1942. [Google Scholar] [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A Simple and Efficient Method for Isolating RNA from Pine Trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Fiancette, R.; Finlay, C.M.; Willis, C.; Bevington, S.L.; Soley, J.; Ng, S.T.H.; Baker, S.M.; Andrews, S.; Hepworth, M.R.; Withers, D.R. Reciprocal Transcription Factor Networks Govern Tissue-Resident ILC3 Subset Function and Identity. Nat. Immunol. 2021, 22, 1245–1255. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Daccord, N.; Celton, J.-M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; Van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-Quality de Novo Assembly of the Apple Genome and Methylome Dynamics of Early Fruit Development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lawrence, M.; Huber, W.; Pagès, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. Software for Computing and Annotating Genomic Ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 2012, 1, 726. [Google Scholar]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, F.; Ryan, D.P.; Grüning, B.; Bhardwaj, V.; Kilpert, F.; Richter, A.S.; Heyne, S.; Dündar, F.; Manke, T. deepTools2: A next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 2016, 44, W160–W165. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Koboldt, D.C.; Chen, K.; Wylie, T.; Larson, D.E.; McLellan, M.D.; Mardis, E.R.; Weinstock, G.M.; Wilson, R.K.; Ding, L. VarScan: Variant Detection in Massively Parallel Sequencing of Individual and Pooled Samples. Bioinformatics 2009, 25, 2283–2285. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Kim, J.H.; Kwon, J.; Jeong, C.Y.; Lee, W.; Lee, D.; Hong, S.-W.; Lee, H. Characterization of Arabidopsis Thaliana FLAVONOL SYNTHASE 1 (FLS1) -Overexpression Plants in Response to Abiotic Stress. Plant Physiol. Biochem. 2016, 103, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.-R.; Yu, Y.-H.; Ni, P.-Y.; Zhang, G.-H.; Guo, D.-L. Genome-Wide Identification of Small Heat-Shock Protein (HSP20) Gene Family in Grape and Expression Profile during Berry Development. BMC Plant Biol. 2019, 19, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neta-Sharir, I.; Isaacson, T.; Lurie, S.; Weiss, D. Dual Role for Tomato Heat Shock Protein 21: Protecting Photosystem II from Oxidative Stress and Promoting Color Changes during Fruit Maturation. Plant Cell 2005, 17, 1829–1838. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Liu, J.-H.; Ma, X.; Zhai, Y.-F.; Gong, Z.-H.; Lu, M.-H. Genome-Wide Analysis of the Hsp70 Family Genes in Pepper (Capsicum Annuum L.) and Functional Identification of CaHsp70-2 Involvement in Heat Stress. Plant Sci. 2016, 252, 246–256. [Google Scholar] [CrossRef]
- Abbas, M.; Peszlen, I.; Shi, R.; Kim, H.; Katahira, R.; Kafle, K.; Xiang, Z.; Huang, X.; Min, D.; Mohamadamin, M.; et al. Involvement of CesA4, CesA7-A/B and CesA8-A/B in Secondary Wall Formation in Populus Trichocarpa Wood. Tree Physiol. 2020, 40, 73–89. [Google Scholar] [CrossRef]
- Zhang, Z.-B.; Zhu, J.; Gao, J.-F.; Wang, C.; Li, H.; Li, H.; Zhang, H.-Q.; Zhang, S.; Wang, D.-M.; Wang, Q.-X.; et al. Transcription Factor AtMYB103 Is Required for Anther Development by Regulating Tapetum Development, Callose Dissolution and Exine Formation in Arabidopsis: Molecular Cloning and Functional Analysis of AtMYB103. Plant J. 2007, 52, 528–538. [Google Scholar] [CrossRef]
- Stracke, R.; Jahns, O.; Keck, M.; Tohge, T.; Niehaus, K.; Fernie, A.R.; Weisshaar, B. Analysis of PRODUCTION OF FLAVONOL GLYCOSIDES-dependent Flavonol Glycoside Accumulation in Arabidopsis Thaliana Plants Reveals MYB11-, MYB12- and MYB111-independent Flavonol Glycoside Accumulation. New Phytol. 2010, 188, 985–1000. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the Anthocyanin Biosynthetic Pathway by the TTG1/BHLH/Myb Transcriptional Complex in Arabidopsis Seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fu, Q.; Chen, L.; Huang, W.; Yu, D. Arabidopsis Thaliana WRKY25, WRKY26, and WRKY33 Coordinate Induction of Plant Thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar] [CrossRef]
- Ishizaki, T.; Maruyama, K.; Obara, M.; Fukutani, A.; Yamaguchi-Shinozaki, K.; Ito, Y.; Kumashiro, T. Expression of Arabidopsis DREB1C Improves Survival, Growth, and Yield of Upland New Rice for Africa (NERICA) under Drought. Mol. Breed. 2013, 31, 255–264. [Google Scholar] [CrossRef]
- Li, F.; Min, D.; Ren, C.; Dong, L.; Shu, P.; Cui, X.; Zhang, X. Ethylene Altered Fruit Cuticular Wax, the Expression of Cuticular Wax Synthesis-Related Genes and Fruit Quality during Cold Storage of Apple (Malus Domestica Borkh. c.v. Starkrimson) Fruit. Postharvest Biol. Technol. 2019, 149, 58–65. [Google Scholar] [CrossRef]
- Biswal, A.K.; Hao, Z.; Pattathil, S.; Yang, X.; Winkeler, K.; Collins, C.; Mohanty, S.S.; Richardson, E.A.; Gelineo-Albersheim, I.; Hunt, K.; et al. Downregulation of GAUT12 in Populus Deltoides by RNA Silencing Results in Reduced Recalcitrance, Increased Growth and Reduced Xylan and Pectin in a Woody Biofuel Feedstock. Biotechnol. Biofuels 2015, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Wu, D.; Shi, J.; Balfour, K.; Wang, H.; Zhu, G.; Liu, Y.; Wang, J.; Zhu, Z. Multiple MYB Activators and Repressors Collaboratively Regulate the Juvenile Red Fading in Leaves of Sweetpotato. Front. Plant Sci. 2020, 11, 941. [Google Scholar] [CrossRef]
- Liu, Z.; Running, M.P.; Meyerowitz, E.M. TSO1 Functions in Cell Division during Arabidopsis Flower Development. Development 1997, 124, 665–672. [Google Scholar] [CrossRef]
- Bajaj, R.; Huang, Y.; Gebrechristos, S.; Mikolajczyk, B.; Brown, H.; Prasad, R.; Varma, A.; Bushley, K.E. Transcriptional Responses of Soybean Roots to Colonization with the Root Endophytic Fungus Piriformospora Indica Reveals Altered Phenylpropanoid and Secondary Metabolism. Sci. Rep. 2018, 18, 10227. [Google Scholar] [CrossRef] [Green Version]
- Wagner, T.A.; Kohorn, B.D. Wall-Associated Kinases Are Expressed throughout Plant Development and Are Required for Cell Expansion. Plant Cell 2001, 13, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, R.; Sorin, C.; Chakraborti, D.; Moritz, T.; Schaller, H.; Tellier, F.; Robert, S.; Morin, H.; Bako, L.; Bellini, C. ABCG9, ABCG11 and ABCG14 ABC Transporters Are Required for Vascular Development in Arabidopsis. Plant J. 2013, 76, 811–824. [Google Scholar] [CrossRef]
- Lima, J.E.; Kojima, S.; Takahashi, H.; Von Wirén, N. Ammonium Triggers Lateral Root Branching in Arabidopsis in an AMMONIUM TRANSPORTER1;3-Dependent Manner. Plant Cell 2010, 22, 3621–3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, Y.-H.; Zubo, Y.O.; Tapken, W.; Kim, H.J.; Lavanway, A.M.; Howard, L.; Pilon, M.; Kieber, J.J.; Schaller, G.E. Functional Characterization of the GATA Transcription Factors GNC and CGA1 Reveals Their Key Role in Chloroplast Development, Growth, and Division in Arabidopsis. Plant Physiol. 2012, 160, 332–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Pan, Y.; Tian, W.; Dong, M.; Zhu, H.; Luan, S.; Li, L. Arabidopsis CNGC14 Mediates Calcium Influx Required for Tip Growth in Root Hairs. Mol. Plant 2017, 10, 1004–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Long, R.; Kang, J.; Wang, Z.; Zhang, T.; Sun, H.; Li, X.; Yang, Q. Comparative Proteomic Analysis Reveals That Antioxidant System and Soluble Sugar Metabolism Contribute to Salt Tolerance in Alfalfa (Medicago Sativa L.) Leaves. J. Proteome Res. 2018, 18, 191–203. [Google Scholar] [CrossRef]
- Castro-Rodríguez, R.; Escudero, V.; Reguera, M.; Gil-Díez, P.; Quintana, J.; Prieto, R.I.; Kumar, R.K.; Brear, E.; Grillet, L.; Wen, J.; et al. Medicago Truncatula Yellow Stripe-Like7 Encodes a Peptide Transporter Participating in Symbiotic Nitrogen Fixation. Plant Cell Environ. 2021, 44, 1908–1920. [Google Scholar] [CrossRef]
- Dash, M.; Yordanov, Y.S.; Georgieva, T.; Tschaplinski, T.J.; Yordanova, E.; Busov, V. Poplar Ptab ZIP 1-like Enhances Lateral Root Formation and Biomass Growth under Drought Stress. Plant J. 2017, 89, 692–705. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Liu, C.; Li, Z.; Ran, Q.; Xie, G.; Wang, B.; Fang, S.; Chu, J.; Zhang, J. ZmbZIP4 Contributes to Stress Resistance in Maize by Regulating ABA Synthesis and Root Development. Plant Physiol. 2018, 178, 753–770. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Zhang, M.; Yang, M.; Hu, Y. Arabidopsis JAZ Proteins Interact with and Suppress RHD6 Transcription Factor to Regulate Jasmonate-Stimulated Root Hair Development. Plant Cell 2020, 32, 1049–1062. [Google Scholar] [CrossRef]
- Peer, W.A.; Hosein, F.N.; Bandyopadhyay, A.; Makam, S.N.; Otegui, M.S.; Lee, G.-J.; Blakeslee, J.J.; Cheng, Y.; Titapiwatanakun, B.; Yakubov, B.; et al. Mutation of the Membrane-Associated M1 Protease APM1 Results in Distinct Embryonic and Seedling Developmental Defects in Arabidopsis. Plant Cell 2009, 21, 1693–1721. [Google Scholar] [CrossRef] [Green Version]
- Demirkol, G. PopW Enhances Drought Stress Tolerance of Alfalfa via Activating Antioxidative Enzymes, Endogenous Hormones, Drought Related Genes and Inhibiting Senescence Genes. Plant Physiol. Biochem. 2021, 166, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Satoh, R.; Fujita, Y.; Nakashima, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A Novel Subgroup of BZIP Proteins Functions as Transcriptional Activators in Hypoosmolarity-Responsive Expression of the ProDH Gene in Arabidopsis. Plant Cell Physiol. 2004, 45, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Grabau, E. Bypassing MiRNA-Mediated Gene Regulation under Drought Stress: Alternative Splicing Affects CSD1 Gene Expression. Plant Mol. Biol. 2017, 95, 243–252. [Google Scholar] [CrossRef]
- Vanani, F.R.; Shabani, L.; Sabzalian, M.R.; Dehghanian, F.; Winner, L. Comparative Physiological and Proteomic Analysis Indicates Lower Shock Response to Drought Stress Conditions in a Self-Pollinating Perennial Ryegrass. PLoS ONE 2020, 15, e0234317. [Google Scholar] [CrossRef] [PubMed]
- Sow, M.D.; Le Gac, A.; Fichot, R.; Lanciano, S.; Delaunay, A.; Le Jan, I.; Lesage-Descauses, M.; Citerne, S.; Caius, J.; Brunaud, V.; et al. RNAi Suppression of DNA Methylation Affects the Drought Stress Response and Genome Integrity in Transgenic Poplar. New Phytol. 2021, 232, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, Q.; Fu, T.; Zhou, Y. Overexpression of the Brassica Napus BnLAS Gene in Arabidopsis Affects Plant Development and Increases Drought Tolerance. Plant Cell Rep. 2011, 30, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.K.; Lin, T.; Hannapel, D.J. Untranslated Regions of a Mobile Transcript Mediate RNA Metabolism. Plant Physiol. 2009, 151, 1831–1843. [Google Scholar] [CrossRef] [Green Version]
- Ham, B.-K.; Brandom, J.L.; Xoconostle-Cázares, B.; Ringgold, V.; Lough, T.J.; Lucas, W.J. A Polypyrimidine Tract Binding Protein, Pumpkin RBP50, Forms the Basis of a Phloem-Mobile Ribonucleoprotein Complex. Plant Cell 2009, 21, 197–215. [Google Scholar] [CrossRef] [Green Version]
- Haywood, V.; Yu, T.-S.; Huang, N.-C.; Lucas, W.J. Phloem Long-Distance Trafficking of GIBBERELLIC ACID-INSENSITIVE RNA Regulates Leaf Development: Phloem Delivery of RNA Regulates Leaf Development. Plant J. 2005, 42, 49–68. [Google Scholar] [CrossRef]
- Kim, M.; Canio, W.; Kessler, S.; Sinha, N. Developmental Changes Due to Long-Distance Movement of a Homeobox Fusion Transcript in Tomato. Sci. New Ser. 2001, 293, 287–289. [Google Scholar] [CrossRef]
- Ruiz-Medrano, R.; Xoconostle-Cazares, B.; Lucas, W.J. Phloem Long-Distance Transport of CmNACP MRNA: Implications for Supracellular Regulation in Plants. Development 1999, 126, 4405–4419. [Google Scholar] [CrossRef] [PubMed]
- Reina-Pinto, J.J.; Voisin, D.; Kurdyukov, S.; Faust, A.; Haslam, R.P.; Michaelson, L.V.; Efremova, N.; Franke, B.; Schreiber, L.; Napier, J.A.; et al. Misexpression of FATTY ACID ELONGATION1 in the Arabidopsis Epidermis Induces Cell Death and Suggests a Critical Role for Phospholipase A2 in This Process. Plant Cell 2009, 21, 1252–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Liu, S.; Tang, B.; Chen, J.; Xie, Z.; Nolan, T.M.; Jiang, H.; Guo, H.; Lin, H.-Y.; Li, L.; et al. RD26 Mediates Crosstalk between Drought and Brassinosteroid Signalling Pathways. Nat. Commun. 2017, 8, 14573. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zayed, O.; Yu, Z.; Jiang, W.; Zhu, P.; Hsu, C.-C.; Zhang, L.; Tao, W.A.; Lozano-Durán, R.; Zhu, J.-K. Leucine-Rich Repeat Extensin Proteins Regulate Plant Salt Tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 13123–13128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kang, J.; Cho, D.-I.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-Binding BZIP Factor, Is an Essential Component of Glucose Signaling and Its Overexpression Affects Multiple Stress Tolerance: ABF2 Is a Positive Component of Glucose Signaling. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Dheilly, E.; Gall, S.L.; Guillou, M.-C.; Renou, J.-P.; Bonnin, E.; Orsel, M.; Lahaye, M. Cell Wall Dynamics during Apple Development and Storage Involves Hemicellulose Modifications and Related Expressed Genes. BMC Plant Biol. 2016, 16, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaahtera, L.; Schulz, J.; Hamann, T. Cell Wall Integrity Maintenance during Plant Development and Interaction with the Environment. Nat. Plants 2019, 5, 924–932. [Google Scholar] [CrossRef]
- Qiu, D.; Xu, S.; Wang, Y.; Zhou, M.; Hong, L. Primary Cell Wall Modifying Proteins Regulate Wall Mechanics to Steer Plant Morphogenesis. Front. Plant Sci. 2021, 12, 751372. [Google Scholar] [CrossRef]
- Shimmen, T. The Sliding Theory of Cytoplasmic Streaming: Fifty Years of Progress. J. Plant Res. 2007, 120, 31–43. [Google Scholar] [CrossRef]
- Karim, S.K.A.; Allan, A.C.; Schaffer, R.J.; David, K.M. Cell Division Controls Final Fruit Size in Three Apple (Malus x Domestica) Cultivars. Horticulturae 2022, 8, 657. [Google Scholar] [CrossRef]
- Gillaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A Developmental Perspective. Plant Cell 1993, 5, 1439. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Wang, Y.; Jiang, W.J.; Liu, X.L.; Zhang, X.M.; Yu, H.J.; Huang, S.W.; Liu, G.Q. Characterization and Expression Profiling of Cucumber Kinesin Genes during Early Fruit Development: Revealing the Roles of Kinesins in Exponential Cell Production and Enlargement in Cucumber Fruit. J. Exp. Bot. 2013, 64, 4541–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onelli, E.; Idilli, A.I.; Moscatelli, A. Emerging Roles for Microtubules in Angiosperm Pollen Tube Growth Highlight New Research Cues. Front. Plant Sci. 2015, 6, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaseem, M.F.; Wu, A.-M. Balanced Xylan Acetylation Is the Key Regulator of Plant Growth and Development, and Cell Wall Structure and for Industrial Utilization. IJMS 2020, 21, 7875. [Google Scholar] [CrossRef]
- Batth, R.; Nicolle, C.; Cuciurean, I.S.; Simonsen, H.T. Biosynthesis and Industrial Production of Androsteroids. Plants 2020, 9, 1144. [Google Scholar] [CrossRef]
- Lombardo, V.A.; Osorio, S.; Borsani, J.; Lauxmann, M.A.; Bustamante, C.A.; Budde, C.O.; Andreo, C.S.; Lara, M.V.; Fernie, A.R.; Drincovich, M.F. Metabolic Profiling during Peach Fruit Development and Ripening Reveals the Metabolic Networks That Underpin Each Developmental Stage. Plant Physiol. 2011, 157, 1696–1710. [Google Scholar] [CrossRef] [Green Version]
- Torres, C.A.; Azocar, C.; Ramos, P.; Pérez-Díaz, R.; Sepulveda, G.; Moya-León, M.A. Photooxidative Stress Activates a Complex Multigenic Response Integrating the Phenylpropanoid Pathway and Ethylene, Leading to Lignin Accumulation in Apple (Malus Domestica Borkh.) Fruit. Hortic. Res. 2020, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Stintzi, A.; Weber, H.; Reymond, P.; Browse, J.; Farmer, E.E. Plant Defense in the Absence of Jasmonic Acid: The Role of Cyclopentenones. Proc. Natl. Acad. Sci. USA 2001, 98, 12837–12842. [Google Scholar] [CrossRef] [Green Version]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Treutter, D. Biosynthesis of Phenolic Compounds and Its Regulation in Apple. Plant Growth Regul. 2001, 34, 71–89. [Google Scholar] [CrossRef]
- Ma, L.; He, J.; Liu, H.; Zhou, H. The Phenylpropanoid Pathway Affects Apple Fruit Resistance to Botrytis Cinerea. J. Phytopathol. 2018, 166, 206–215. [Google Scholar] [CrossRef]
- Francini, A.; Sebastiani, L. Phenolic Compounds in Apple (Malus x Domestica Borkh.): Compounds Characterization and Stability during Postharvest and after Processing. Antioxidants 2013, 2, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Cheng, L. The elevated anthocyanin level in the shaded peel of ‘Anjou’ pear enhances its tolerance to high temperature under high light. Plant Sci. 2009, 177, 418–426. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, H.; Zhang, Y.; Zhang, J.; Niu, S.; Wang, X.; Li, W.; Zhang, J.; Yao, Y. The MdMYB16/MdMYB1-miR7125-MdCCR Module Regulates the Homeostasis between Anthocyanin and Lignin Biosynthesis during Light Induction in Apple. New Phytol 2021, 231, 1105–1122. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, C.; Gao, Y.; Xu, Y.; Wang, S.; Li, C.; Xie, Y.; Chen, P.; Yang, P.; Yuan, L.; et al. A Multifaceted Module of BRI1 ETHYLMETHANE SULFONATE SUPRESSOR1 (BES1)-MYB88 in Growth and Stress Tolerance of Apple. Plant Physiol. 2021, 185, 1903–1923. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Chmur, M.; Gruszka, D. Comprehensive Overview of the Brassinosteroid Biosynthesis Pathways: Substrates, Products, Inhibitors, and Connections. Front. Plant Sci. 2020, 11, 1034. [Google Scholar] [CrossRef] [PubMed]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.; Yang, L.; Miñambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis Messenger RNAs Transported to Distant Tissues. Nat. Plants 2015, 1, 15025. [Google Scholar] [CrossRef]
- Zhang, W.-N.; Gong, L.; Ma, C.; Xu, H.-Y.; Hu, J.-F.; Harada, T.; Li, T.-Z. Gibberellic Acid-Insensitive MRNA Transport in Pyrus. Plant Mol. Biol. Rep. 2012, 30, 614–623. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, W.; Li, M.; Harada, T.; Han, Z.; Li, T. Gibberellic Acid Insensitive MRNA Transport in Both Directions between Stock and Scion in Malus. Tree Genet. Genomes 2010, 6, 1013–1019. [Google Scholar] [CrossRef]
- Xu, H.; Iwashiro, R.; Li, T.; Harada, T. Long-Distance Transport of Gibberellic Acid Insensitive MRNA in Nicotiana Benthamiana. BMC Plant Biol. 2013, 13, 165. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.N.; Duan, X.W.; Ma, C.; Harada, T.; Li, T.Z. Transport of MRNA Molecules Coding NAC Domain Protein in Grafted Pear and Transgenic Tobacco. Biol. Plant. 2013, 57, 224–230. [Google Scholar] [CrossRef]
- Xia, C.; Zheng, Y.; Huang, J.; Zhou, X.; Li, R.; Zha, M.; Wang, S.; Huang, Z.; Lan, H.; Turgeon, R.; et al. Elucidation of the Mechanisms of Long-Distance MRNA Movement in a Nicotiana Benthamiana /Tomato Heterograft System. Plant Physiol. 2018, 177, 745–758. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Raw Base (G) | Clean Base (G) | Q20 (%) | Q30 (%) | GC Content (%) | Mapping Ratio (%) |
|---|---|---|---|---|---|---|---|---|
| Control “Fuji/GL-3” Rep1 | 25,143,068 | 23,973,149 | 7.54 | 7.19 | 98.28 | 94.71 | 47.32 | 93.36 |
| Control “Fuji/GL-3” Rep2 | 25,752,761 | 24,541,277 | 7.73 | 7.36 | 98.25 | 94.61 | 47.34 | 93.24 |
| Control “Fuji/GL-3” Rep1 | 22,824,047 | 21,670,561 | 6.85 | 6.5 | 98.31 | 94.75 | 47.45 | 93.47 |
| Control “Fuji/GL-3” Rep2 | 20,255,453 | 19,353,326 | 6.08 | 5.81 | 98.31 | 94.78 | 47.41 | 93.56 |
| Control “Fuji/MdGH3 RNAi” Rep1 | 23,568,295 | 22,345,147 | 7.07 | 6.7 | 98.21 | 94.53 | 47.22 | 93.06 |
| Control “Fuji/MdGH3 RNAi” Rep2 | 23,744,639 | 22,762,112 | 7.12 | 6.83 | 98.31 | 94.73 | 46.8 | 93.77 |
| Control “Fuji/MdGH3 RNAi” Rep1 | 20,469,321 | 19,537,242 | 6.14 | 5.86 | 98.25 | 94.6 | 47.24 | 93.96 |
| Control “Fuji/MdGH3 RNAi” Rep2 | 22,951,961 | 21,696,492 | 6.89 | 6.51 | 98.34 | 94.76 | 47.19 | 94.06 |
| Drought “Fuji/GL-3” Rep1 | 20,040,778 | 18,701,561 | 6.01 | 5.61 | 98.29 | 94.72 | 47.14 | 93.33 |
| Drought “Fuji/GL-3” Rep2 | 21,133,554 | 20,108,894 | 6.34 | 6.03 | 98.37 | 94.86 | 47.25 | 93.38 |
| Drought “Fuji/GL-3” Rep1 | 20,435,878 | 19,390,591 | 6.13 | 5.82 | 98.36 | 94.91 | 47.35 | 93.51 |
| Drought “Fuji/GL-3” Rep2 | 22,430,471 | 21,593,803 | 6.73 | 6.48 | 98.34 | 94.81 | 47.22 | 93.84 |
| Drought “Fuji/MdGH3 RNAi” Rep1 | 21,519,114 | 20,814,869 | 6.46 | 6.24 | 98.34 | 94.78 | 47.12 | 93.53 |
| Drought “Fuji/MdGH3 RNAi” Rep2 | 21,640,685 | 20,732,825 | 6.49 | 6.22 | 98.23 | 94.52 | 47.02 | 93.09 |
| Drought “Fuji/MdGH3 RNAi” Rep1 | 25,823,890 | 24,540,545 | 7.75 | 7.36 | 97.67 | 93.23 | 47.09 | 98.89 |
| Drought “Fuji/MdGH3 RNAi” Rep2 | 21,009,641 | 20,148,636 | 6.3 | 6.04 | 98.28 | 94.69 | 47.05 | 93.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Guo, J.; Jiang, L.; An, W.; Ma, F.; Guan, Q.; Niu, C. Transcriptional Effects of Rootstock on Scion after Drought: A Case Study of Using MdGH3 RNAi as the Rootstock. Horticulturae 2022, 8, 1212. https://doi.org/10.3390/horticulturae8121212
He J, Guo J, Jiang L, An W, Ma F, Guan Q, Niu C. Transcriptional Effects of Rootstock on Scion after Drought: A Case Study of Using MdGH3 RNAi as the Rootstock. Horticulturae. 2022; 8(12):1212. https://doi.org/10.3390/horticulturae8121212
Chicago/Turabian StyleHe, Jieqiang, Junxing Guo, Lijuan Jiang, Wenjing An, Fengwang Ma, Qingmei Guan, and Chundong Niu. 2022. "Transcriptional Effects of Rootstock on Scion after Drought: A Case Study of Using MdGH3 RNAi as the Rootstock" Horticulturae 8, no. 12: 1212. https://doi.org/10.3390/horticulturae8121212
APA StyleHe, J., Guo, J., Jiang, L., An, W., Ma, F., Guan, Q., & Niu, C. (2022). Transcriptional Effects of Rootstock on Scion after Drought: A Case Study of Using MdGH3 RNAi as the Rootstock. Horticulturae, 8(12), 1212. https://doi.org/10.3390/horticulturae8121212