
Correlations between the Phylogenetic Relationship of 14 Tulasnella Strains and Their Promotion Effect on Dendrobium crepidatum Protocorm

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolation and Identification
2.2. Protocorm Formation
2.3. Symbiotic Culture of Protocorms
2.4. Assessment of the Fungal Capacity to Promote Protocorm and Seedlings Growth
2.5. Statistical Analysis
3. Results
3.1. Tulasnella Identification and Phylogenetic Analyses
3.2. Compatibility between Tulasnella Stains and D. crepidatum Protocorms
3.3. Effects of different Tulasnella Strains on the Biomass of the Symbionts
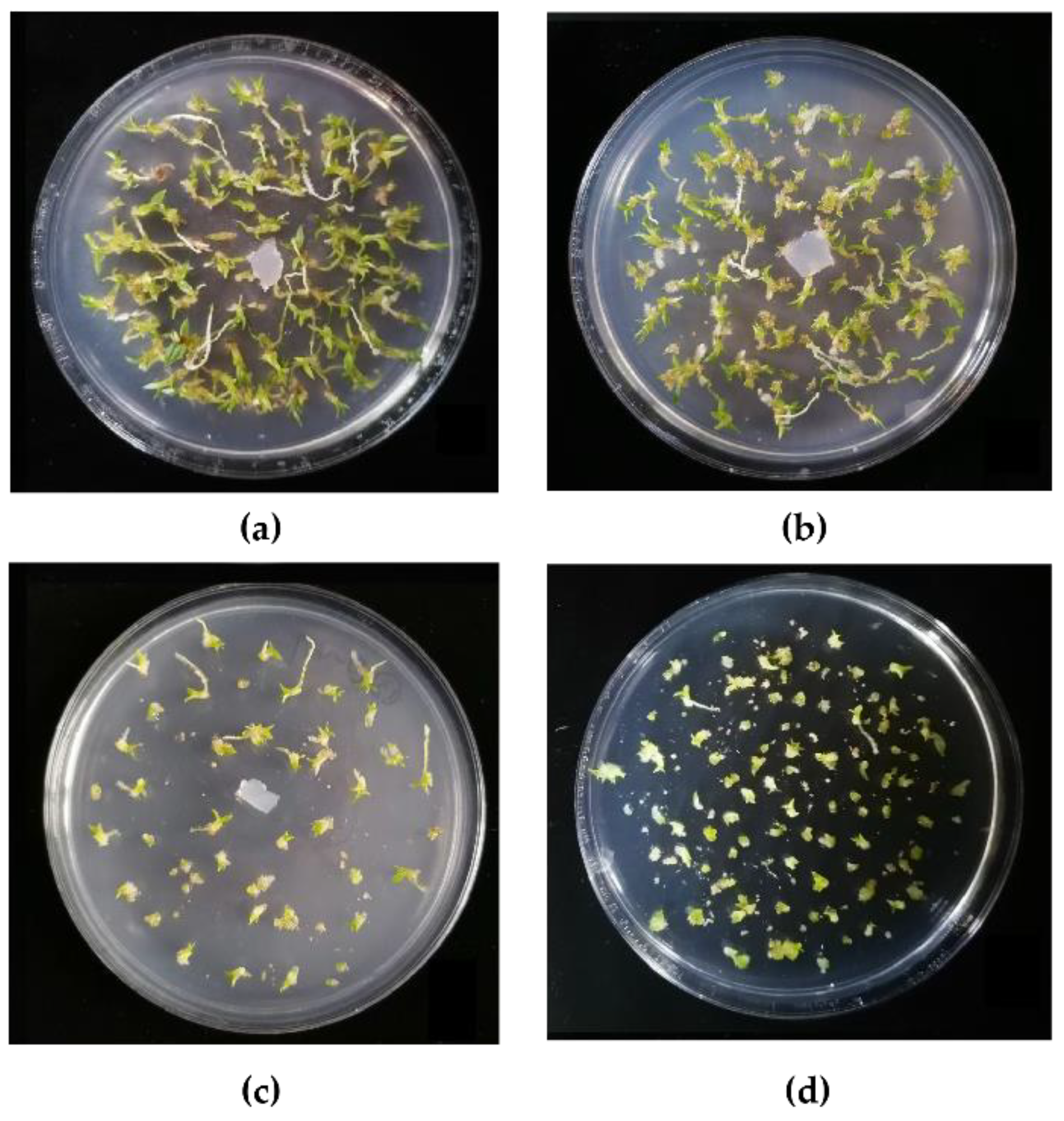
3.4. Effects of Different Tulasnella Strains on the Morphology of the Symbionts
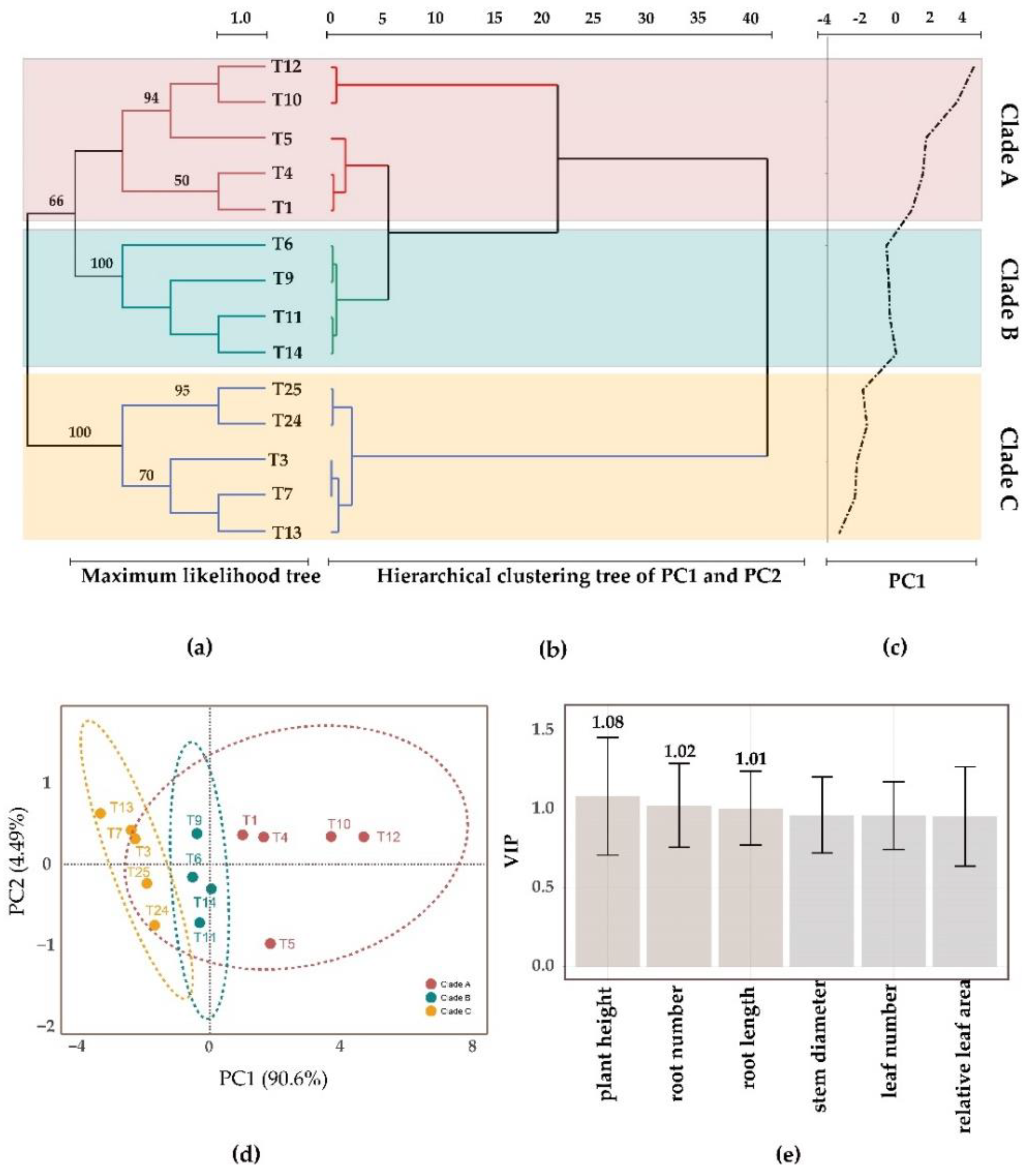
3.5. Association between Relationship and Symbiotic Effects among Different Tulasnella Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Favre-Godal, Q.; Gourguillon, L.; Lordel-Madeleine, S.; Gindro, K.; Choisy, P. Orchids and their mycorrhizal fungi: An insufficiently explored relationship. Mycorrhiza 2020, 30, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.C.; Luo, Y.; Jacquemyn, H. Co-Cultures of Mycorrhizal Fungi Do Not Increase Germination and Seedling Development in the Epiphytic Orchid Dendrobium nobile. Front. Plant Sci. 2020, 11, 571426. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.H.; Chen, X.G.; Selosse, M.A.; Gao, J.Y. Compatible and Incompatible Mycorrhizal Fungi With Seeds of Dendrobium Species: The Colonization Process and Effects of Coculture on Germination and Seedling Development. Front. Plant Sci. 2022, 13, 823794. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-G.; Wu, Y.-H.; Li, N.-Q.; Gao, J.-Y. What role does the seed coat play during symbiotic seed germination in orchids: An experimental approach with Dendrobium officinale. BMC Plant Biol. 2022, 22, 375. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-M.; Chung, K.; Liang, C.-K.; Tsai, W.-C. New Insights into the Symbiotic Relationship between Orchids and Fungi. Appl. Sci. 2019, 9, 585. [Google Scholar] [CrossRef] [Green Version]
- Yeung, E.C. A perspective on orchid seed and protocorm development. Bot. Stud. 2017, 58, 33. [Google Scholar] [CrossRef] [Green Version]
- Dearnaley, J.; Perotto, S.; Selosse, M.-A. Structure and development of orchid mycorrhizas. Mol. Mycorrhizal Symbiosis 2016, 2016, 63–86. [Google Scholar] [CrossRef]
- Peterson, R.L.; Farquhar, M.L. Mycorrhizas-integrated development between roots and fungi. Mycologia 1994, 86, 311–326. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef]
- Adamo, M.; Chialva, M.; Calevo, J.; Rose, S.; Girlanda, M.; Perotto, S.; Balestrini, R. The Dark Side of Orchid Symbiosis: Can Tulasnella calospora Decompose Host Tissues? Int. J. Mol. Sci. 2020, 21, 3139. [Google Scholar] [CrossRef] [PubMed]
- Dearnaley, J.; Martos, F.; Selosse, M. Fungal Associations. In The Mycota; Springer: Berlin/Heidelberg, Germany, 2012; p. 207. [Google Scholar] [CrossRef]
- Herrera, H.; Valadares, R.; Contreras, D.; Bashan, Y.; Arriagada, C. Mycorrhizal compatibility and symbiotic seed germination of orchids from the Coastal Range and Andes in south central Chile. Mycorrhiza 2017, 27, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Freitas, E.F.S.; da Silva, M.; Cruz, E.D.S.; Mangaravite, E.; Bocayuva, M.F.; Veloso, T.G.R.; Selosse, M.A.; Kasuya, M.C.M. Diversity of mycorrhizal Tulasnella associated with epiphytic and rupicolous orchids from the Brazilian Atlantic Forest, including four new species. Sci. Rep. 2020, 10, 7069. [Google Scholar] [CrossRef] [PubMed]
- Solis, K.; Jose Barriuso, J.; Garces-Claver, A.; Gonzalez, V. Tulasnella tubericola (Tulasnellaceae, Cantharellales, Basidiomycota): A new Rhizoctonia-like fungus associated with mycorrhizal evergreen oak plants artificially inoculated with black truffle (Tuber melanosporum) in Spain. Phytotaxa 2017, 317, 175–187. [Google Scholar] [CrossRef]
- Linde, C.C.; May, T.W.; Phillips, R.D.; Ruibal, M.; Smith, L.M.; Peakall, R. New species of Tulasnella associated with terrestrial orchids in Australia. IMA Fungus 2017, 8, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Kompe, Y.O.; Mutlu, V.A.; Ozkoc, I.; Demiray, S.; Bozkurt, S. Fungal diversity and ex vitro symbiotic germination of Serapias vomeracea (Orchidaceae). Acta Bot. Croat. 2022, 81, 108–116. [Google Scholar] [CrossRef]
- Jasinge, N.U.; Huynh, T.; Lawrie, A.C. Consequences of season of prescribed burning on two spring-flowering terrestrial orchids and their endophytic fungi. Aust. J. Bot. 2018, 66, 298–312. [Google Scholar] [CrossRef]
- Thakur, J.; Dwivedi, M.D.; Uniyal, P.L. Ultrastructural studies and molecular characterization of root-associated fungi of Crepidium acuminatum (D. Don) Szlach.: A threatened and medicinally important taxon. J. Genet. 2018, 97, 1139–1146. [Google Scholar] [CrossRef]
- Lee, S.G.; Lee, J.S.; Choi, S. Identification of Mycorrhizal Symbiont Detected in Roots of Two Cymbidium Species Native to Korea. Flower Res. J. 2019, 27, 17–23. [Google Scholar] [CrossRef]
- Linde, C.C.; Phillips, R.D.; Crisp, M.D.; Peakall, R. Congruent species delineation of Tulasnella using multiple loci and methods. New Phytol. 2014, 201, 6–12. [Google Scholar] [CrossRef]
- Rachanarin, C.; Suwannarach, N.; Kumla, J.; Srimuang, K.-O.; McKenzie, E.H.C.; Lumyong, S. A new endophytic fungus, Tulasnella phuhinrongklaensis (Cantharellales, Basidiomycota) isolated from roots of the terrestrial orchid, Phalaenopsis pulcherrima. Phytotaxa 2018, 374, 99–109. [Google Scholar] [CrossRef]
- Suarez, J.P.; Weiss, M.; Abele, A.; Garnica, S.; Oberwinkler, F.; Kottke, I. Diverse tulasnelloid fungi form mycorrhizas with epiphytic orchids in an Andean cloud forest. Mycol. Res. 2006, 110, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Arifin, A.R.; Reiter, N.H.; May, T.W.; Linde, C.C. New species of Tulasnella associated with Australian terrestrial orchids in the subtribes Megastylidinae and Thelymitrinae. Mycologia 2022, 114, 388–412. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.; Assabgui, R.A.; Seifert, K. Taxonomy and phylogeny of the basidiomycetous hyphomycete genus Hormomyces. Fungal Syst. Evol. 2021, 7, 177–196. [Google Scholar] [CrossRef] [PubMed]
- Reiter, N.; Lawrie, A.C.; Linde, C.C. Matching symbiotic associations of an endangered orchid to habitat to improve conservation outcomes. Ann. Bot. 2018, 122, 947–959. [Google Scholar] [CrossRef]
- Arifin, A.R.; May, T.W.; Linde, C.C. New species of Tulasnella associated with Australian terrestrial orchids in the Cryptostylidinae and Drakaeinae. Mycologia 2021, 113, 212–230. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenet. Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- He, M.-Q.; Zhao, R.-L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.-L.; Li, G.-J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.-L.; Wu, G.; Dai, Y.-C.; He, S.-H.; Cui, B.-K.; Zhou, J.-L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 43–74. [Google Scholar] [CrossRef]
- Huang, H.; Zi, X.M.; Lin, H.; Gao, J.Y. Host-specificity of symbiotic mycorrhizal fungi for enhancing seed germination, protocorm formation and seedling development of over-collected medicinal orchid, Dendrobium devonianum. J. Microbiol. 2018, 56, 42–48. [Google Scholar] [CrossRef]
- Wang, X.-J.; Wu, Y.-H.; Ming, X.-J.; Wang, G.; Gao, J.-Y. Isolating ecological-specific fungi and creating fungus-seed bags for epiphytic orchid conservation. GECCO 2021, 28, e01714. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.Y.; Chen, X.M.; Guo, S.X.; Lee, Y.I. Effect of different mycobionts on symbiotic germination and seedling growth of Dendrobium officinale, an important medicinal orchid. Bot. Stud. 2020, 61, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujari, I.; Sankar Babu, V. Precocious in vitro flowering in threatened ornamental orchid, Dendrobium ovatum—Decoding the causal factors. Curr. Plant Biol. 2022, 31, 100257. [Google Scholar] [CrossRef]
- Liu, J.; Yu, L.; Wang, C.; Zhang, Y.; Xi, H.; Si, J.; Zhang, L.; Yan, J. Preparation, Structural Features and in vitro Immunostimulatory Activity of a Glucomannan From Fresh Dendrobium catenatum Stems. Front. Nutr. 2021, 8, 823803. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.; Jang, H.-J.; Han, A.-R.; Kim, Y.-R.; Jin, C.-H.; Jung, C.-H.; Kang, K.-B.; Kim, S.-H.; Hong, M.-J.; Kim, J.-B.; et al. Chemical and Biological Profiles of Dendrobium in Two Different Species, Their Hybrid, and Gamma-Irradiated Mutant Lines of the Hybrid Based on LC-QToF MS and Cytotoxicity Analysis. Plants 2021, 10, 1376. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.-Y.; Shao, S.-C.; Liu, S.-J.; Gao, J.-Y. Do the fungi associated with roots of adult plants support seed germination? A case study on Dendrobium exile (Orchidaceae). GECCO 2019, 17, e00582. [Google Scholar] [CrossRef]
- Meng, Y.-Y.; Fan, X.-L.; Zhou, L.-R.; Shao, S.-C.; Liu, Q.; Selosse, M.-A.; Gao, J.-Y. Symbiotic fungi undergo a taxonomic and functional bottleneck during orchid seeds germination: A case study on Dendrobium moniliforme. Symbiosis 2019, 79, 205–212. [Google Scholar] [CrossRef]
- Shao, S.C.; Wang, Q.X.; Beng, K.C.; Zhao, D.K.; Jacquemyn, H. Fungi isolated from host protocorms accelerate symbiotic seed germination in an endangered orchid species (Dendrobium chrysotoxum) from southern China. Mycorrhiza 2020, 30, 529–539. [Google Scholar] [CrossRef]
- Chen, D.-Y.; Wang, X.-J.; Li, T.-Q.; Li, N.-Q.; Gao, J.-Y. In situ seedling baiting to isolate plant growth-promoting fungi from Dendrobium officinale, an over-collected medicinal orchid in China. GECCO 2021, 28, e01659. [Google Scholar] [CrossRef]
- Ding, X.-Q.; Zou, Y.-Q.; Liu, J.; Wang, X.-C.; Hu, Y.; Liu, X.; Zhang, C.-F. Dendrocrepidamine, a novel octahydroindolizine alkaloid from the roots of Dendrobium crepidatum. J. Asian Nat. Prod. Res. 2021, 23, 1085–1092. [Google Scholar] [CrossRef]
- Xu, X.L.; Li, Z.S.; Yang, R.M.; Zhou, H.G.; Bai, Y.B.; Yu, M.; Ding, G.; Li, B. Crepidatumines C and D, Two New Indolizidine Alkaloids from Dendrobium crepidatum Lindl. ex Paxt. Molecules 2019, 24, 3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Ren, J.; Wang, L.; Zhao, X.; Zhang, M.; Shimizu, K.; Zhang, C.F. Protective effects of total alkaloids from Dendrobium crepidatum against LPS-induced acute lung injury in mice and its chemical components. Phytochemistry 2018, 149, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Liang, Z.; Zhang, L.; Hu, L.; Fu, J.; Wei, G.; Huang, Y. Identification of three types of O-glycosylated flavonoids in Dendrobium loddigesii, Dendrobium primulinum, Dendrobium crepidatum, Dendrobium porphyrochilum and Dendrobium hancockii by MS. Rapid Commun. Mass Spectrom. 2022, 37, e9421. [Google Scholar] [CrossRef] [PubMed]
- Paudel, M.R.; Chand, M.B.; Pant, B.; Pant, B. Assessment of Antioxidant and Cytotoxic Activities of Extracts of Dendrobium crepidatum. Biomolecules 2019, 9, 478. [Google Scholar] [CrossRef] [Green Version]
- Swangmaneecharern, P.; Serivichyaswat, P.T.; Nontachaiyapoom, S. Promoting effect of orchid mycorrhizal fungi Epulorhiza isolates on seed germination of Dendrobium orchids. Sci. Hortic. 2012, 148, 55–58. [Google Scholar] [CrossRef]
- Nguyen, D.Q.; Li, H.; Tran, T.T.; Sivasithamparam, K.; Jones, M.G.K.; Wylie, S.J. Four Tulasnella taxa associated with populations of the Australian evergreen terrestrial orchid Cryptostylis ovata. Fungal Biol. 2020, 124, 24–33. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.X.; Gross, J.; Yoon, H.S.; Bhattacharya, D. Plastid origin and evolution: New models provide insights into old problems. Plant Physiol. 2011, 155, 1552–1560. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Xu, L.; Xia, W.; Liang, L.; Bai, X.; Li, L.; Xu, L.; Liu, L. Mycorrhizal Compatibility and Germination-Promoting Activity of Tulasnella Species in Two Species of Orchid (Cymbidium mannii and Epidendrum radicans). Horticulturae 2021, 7, 472. [Google Scholar] [CrossRef]
- Alam, M.K.; Rashid, M.H.; Hossain, M.S.; Salam, M.A.; Rouf, M.A. In Vitro Seed Propagation of Dendrobium (Dendrobium transparens) Orchid as Influenced by Different Media. Biotechnology 2002, 1, 111–115. [Google Scholar] [CrossRef]
- Meng, Y.Y.; Zhang, W.L.; Selosse, M.A.; Gao, J.Y. Are fungi from adult orchid roots the best symbionts at germination? A case study. Mycorrhiza 2019, 29, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.; Stanley, M.; Day, J.; Semiao, A. A comparison of methods for the non-destructive fresh weight determination of filamentous algae for growth rate analysis and dry weight estimation. J. Appl. Phycol. 2017, 29, 1725–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Gao, J. In vitro tetraploid induction from multigenotype protocorms and tetraploid regeneration in Dendrobium officinale. PCTOC 2020, 141, 289–298. [Google Scholar] [CrossRef]
- Yu, X.; Shi, P.; Schrader, J.; Niklas, K. Nondestructive estimation of leaf area for 15 species of vines with different leaf shapes. Am. J. Bot. 2020, 107, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- Smith, J. Mycorrhizal Symbiosis, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2009; Volume 73. [Google Scholar] [CrossRef]
- Tsavkelova, E.; Kolomeitseva, G. Fusarium–orchid interactions under greenhouse conditions. S. Afr. J. Bot. 2022, 146, 889–896. [Google Scholar] [CrossRef]
- Warcup, J.; Talbot, P. Perfect states of Rhizoctonia associated with orchids III. New Phytol. 2006, 70, 35–40. [Google Scholar] [CrossRef]
- Cruz, D.; Suarez, J.P.; Piepenbring, M. Morphological revision of Tulasnellaceae, with two new species of Tulasnella and new records of Tulasnella spp. for Ecuador. Nova Hedwig. 2016, 102, 279–338. [Google Scholar] [CrossRef]
- Currah, R.; Sigler, L.; Hambleton, S. New records and new taxa of fungi from the mycorrhizae of terrestrial orchids of Alberta. Can. J. Bot. 2011, 65, 2473–2482. [Google Scholar] [CrossRef]
- Milligan, M.; Williams, P. The mycorrhizal relationship of multinucleate rhizoctonias from non-orchids with Microtis (Orchidaceae). New Phytol. 2006, 108, 205–209. [Google Scholar] [CrossRef]
- Andersen, T. A comparative taxonomic study of Rhizoctonia sensu lato employing morphological, ultrastructural and molecular methods. Mycol. Res. 1996, 100, 1117–1128. [Google Scholar] [CrossRef]
- Cruz, D.; Suárez, J.P.; Kottke, I.; Piepenbring, M. Cryptic species revealed by molecular phylogenetic analysis of sequences obtained from basidiomata of Tulasnella. Mycologia 2014, 106, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Jacquemyn, H.; Brys, R.; Merckx, V.; Waud, M.; Lievens, B.; Wiegand, T. Coexisting orchid species have distinct mycorrhizal communities and display strong spatial segregation. New Phytol. 2014, 202, 616–627. [Google Scholar] [CrossRef] [PubMed]
- González, D.; Rodriguez-Carres, M.; Boekhout, T.; Stalpers, J.; Kuramae, E.; Nakatani, A.; Vilgalys, R.; Cubeta, M. Phylogenetic relationships of Rhizoctonia fungi within the Cantharellales. Fungal Biol. 2016, 120, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Rannala, B. Bayesian species delimitation using multilocus sequence data. Proc. Natl. Acad. Sci. USA 2010, 107, 9264–9269. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Leache, A.; Burbrink, F.; McGuire, J.; Moritz, C. Coalescent-based species delimitation in integrative taxonomy. Trends Ecol. Evol. 2012, 27, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Jeewon, R.; Hyde, K. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Ruibal, M.P.; Peakall, R.; Smith, L.M.; Linde, C.C. Phylogenetic and microsatellite markers for Tulasnella (Tulasnellaceae) mycorrhizal fungi associated with Australian orchids. Appl. Plant Sci. 2013, 1, 1200394. [Google Scholar] [CrossRef]
- Tang, Y.-J.; Zhou, D.-Y.; Dai, J.; Li, Y.; Xing, Y.-M.; Guo, S.-X.; Chen, J. Potential Specificity between Mycorrhizal Fungi Isolated from Widespread Dendrobium spp. and Rare, D. huoshanense Seeds. Curr. Microbiol. 2022, 79, 264. [Google Scholar] [CrossRef]
- Tĕšitelová, T.; Tĕšitel, J.; Jersáková, J.; RÍhová, G.; Selosse, M.A. Symbiotic germination capability of four Epipactis species (Orchidaceae) is broader than expected from adult ecology. Am. J. Bot. 2012, 99, 1020–1032. [Google Scholar] [CrossRef]
- Tan, X.M.; Wang, C.L.; Chen, X.M.; Zhou, Y.Q.; Wang, Y.Q.; Luo, A.X.; Liu, Z.H.; Guo, S.X. In vitro seed germination and seedling growth of an endangered epiphytic orchid, Dendrobium officinale, endemic to China using mycorrhizal fungi (Tulasnella sp.). Sci. Hortic. 2014, 165, 62–68. [Google Scholar] [CrossRef]
- Liu, S.; Liu, M.; Liao, Q.G.; Lu, F.B.; Zhao, X.L. Effects of inoculated mycorrhizal fungi and non-mycorrhizal beneficial micro-organisms on plant traits, nutrient uptake and root-associated fungal community composition of the Cymbidium hybridum in greenhouse. J. Appl. Microbiol. 2021, 131, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huo, W.; Hou, J.; Liu, L.; Yu, X.; Xu, L. Effects and Benefits of Orchid Mycorrhizal Symbionts on Dendrobium officinale. Horticulturae 2022, 8, 861. [Google Scholar] [CrossRef]
- Fuji, M.; Miura, C.; Yamamoto, T.; Komiyama, S.; Suetsugu, K.; Yagame, T.; Yamato, M.; Kaminaka, H. Relative effectiveness of Tulasnella fungal strains in orchid mycorrhizal symbioses between germination and subsequent seedling growth. Symbiosis 2020, 81, 53–63. [Google Scholar] [CrossRef]
- Chen, J.; Yan, B.; Tang, Y.; Xing, Y.; Li, Y.; Zhou, D.; Guo, S. Symbiotic and Asymbiotic Germination of Dendrobium officinale (Orchidaceae) Respond Differently to Exogenous Gibberellins. Int. J. Mol. Sci. 2020, 21, 6104. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Species | Original Plants | NCBI Accession Number | |||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | TEF | |||
| T1 | T. calospora | C. mannii | OM672252 | OM728181 | OP537825 | OP819646 |
| T3 | T. asymmetrica | E. radicans | OM672253 | OM728182 | OP537826 | OP819647 |
| T4 | T. calospora | C. mannii | OM672254 | OM728183 | OP537827 | OP819648 |
| T5 | T. calospora | C. faberi | OM672255 | OM728184 | OP537828 | OP819649 |
| T6 | T. calospora | C. faberi | OM672256 | OM728185 | OP537829 | OP819650 |
| T7 | T. asymmetrica | C. mannii | OM672257 | OM728186 | OP537830 | OP819651 |
| T9 | T. calospora | C. mannii | OM672258 | OM728187 | OP537831 | OP819652 |
| T10 | T. calospora | C. goeringii | OM672259 | OM728188 | OP537832 | OP819653 |
| T11 | T. calospora | C. goeringii | OM672260 | OM728189 | OP537833 | OP819654 |
| T12 | T. calospora | C. goeringii | OM672261 | OM728190 | OP537834 | OP819655 |
| T13 | T. asymmetrica | C. goeringii | OM672262 | OM728191 | OP537835 | OP819656 |
| T14 | T. calospora | E. radicans | OM672263 | OM728192 | OP537836 | OP819657 |
| T24 | T. asymmetrica | E.radicans | OM672264 | OM728193 | OP537837 | OP819658 |
| T25 | T. asymmetrica | C. mannii | OM672265 | OM728194 | OP537838 | OP819659 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Li, Z.; Wang, S.; Yang, F.; Li, L.; Liu, L. Correlations between the Phylogenetic Relationship of 14 Tulasnella Strains and Their Promotion Effect on Dendrobium crepidatum Protocorm. Horticulturae 2022, 8, 1213. https://doi.org/10.3390/horticulturae8121213
Zhao J, Li Z, Wang S, Yang F, Li L, Liu L. Correlations between the Phylogenetic Relationship of 14 Tulasnella Strains and Their Promotion Effect on Dendrobium crepidatum Protocorm. Horticulturae. 2022; 8(12):1213. https://doi.org/10.3390/horticulturae8121213
Chicago/Turabian StyleZhao, Jiayi, Zhenjian Li, Siyu Wang, Fu Yang, Lubin Li, and Lei Liu. 2022. "Correlations between the Phylogenetic Relationship of 14 Tulasnella Strains and Their Promotion Effect on Dendrobium crepidatum Protocorm" Horticulturae 8, no. 12: 1213. https://doi.org/10.3390/horticulturae8121213
APA StyleZhao, J., Li, Z., Wang, S., Yang, F., Li, L., & Liu, L. (2022). Correlations between the Phylogenetic Relationship of 14 Tulasnella Strains and Their Promotion Effect on Dendrobium crepidatum Protocorm. Horticulturae, 8(12), 1213. https://doi.org/10.3390/horticulturae8121213