
Stable Soil Moisture Alleviates Water Stress and Improves Morphogenesis of Tomato Seedlings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Conditions
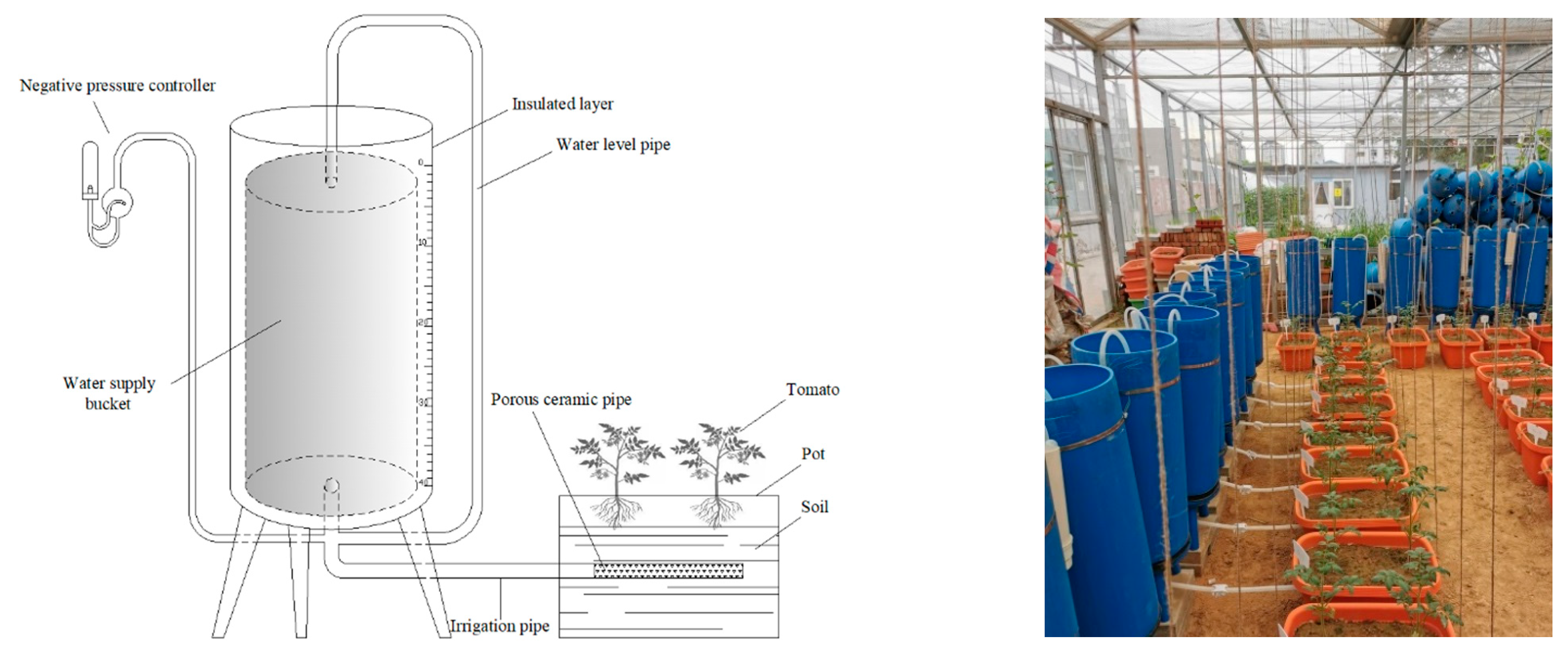
2.2. Device Used to Maintain Stable Soil Moisture
2.3. Experimental Design
2.4. Sampling and Measurements
2.4.1. Fluctuation Coefficient of the SWC
2.4.2. Sampling and Measurements of Tomato Shoots
2.4.3. Sampling and Measurements of Tomato Roots
2.4.4. Morphological Indicators
2.4.5. Leaf Photosynthetic Pigments and Gas Exchange
2.4.6. Physiological Indicators
2.5. Statistical Analysis
3. Results
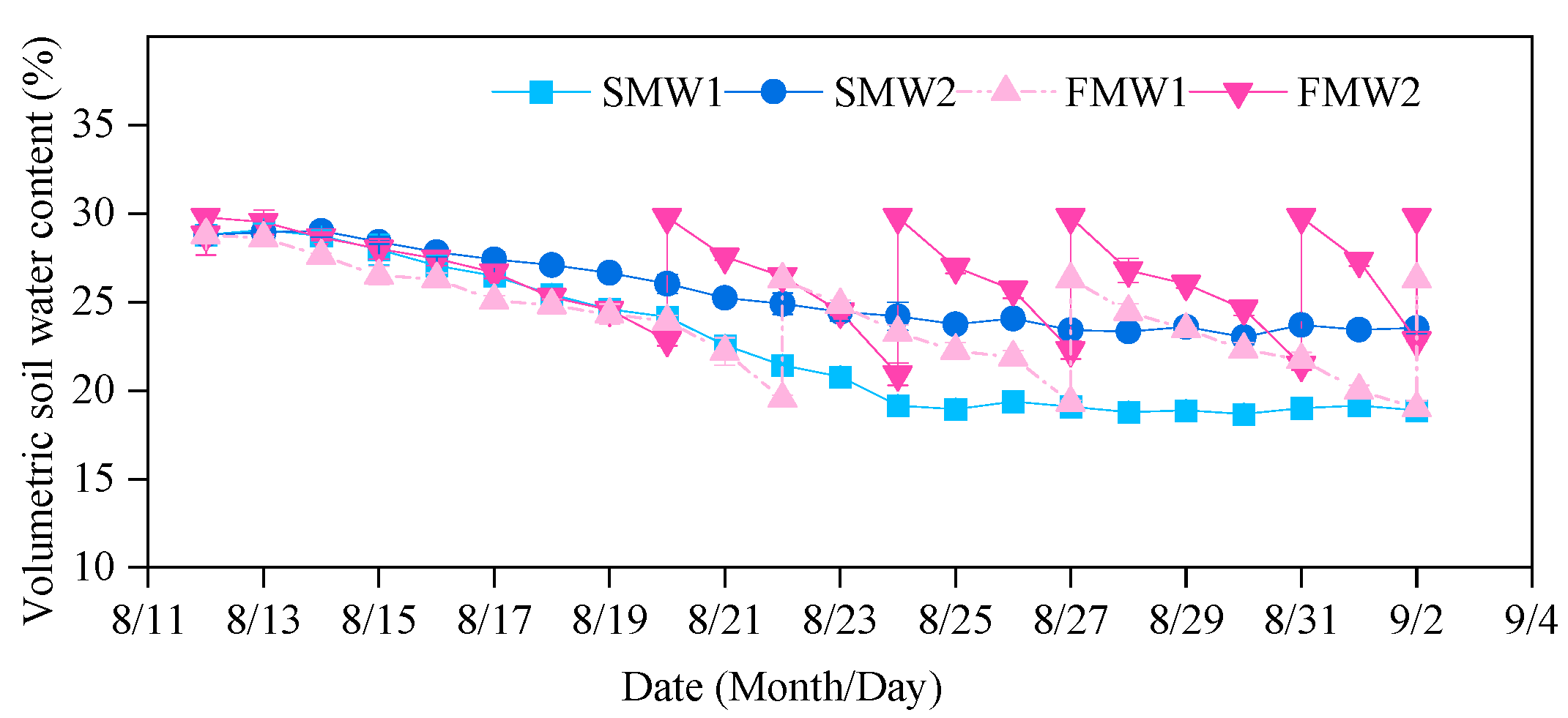
3.1. Soil Moisture Parameters
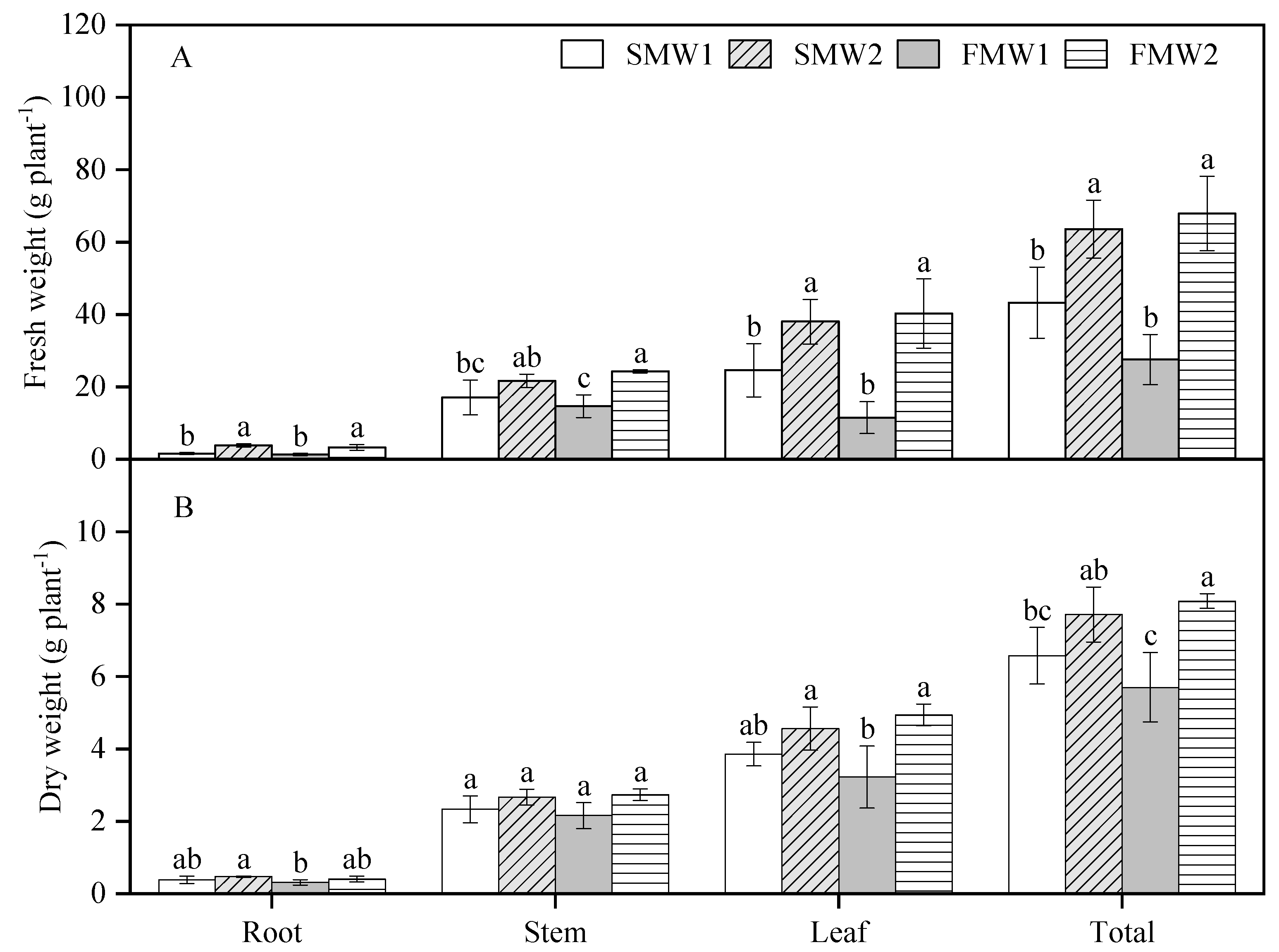
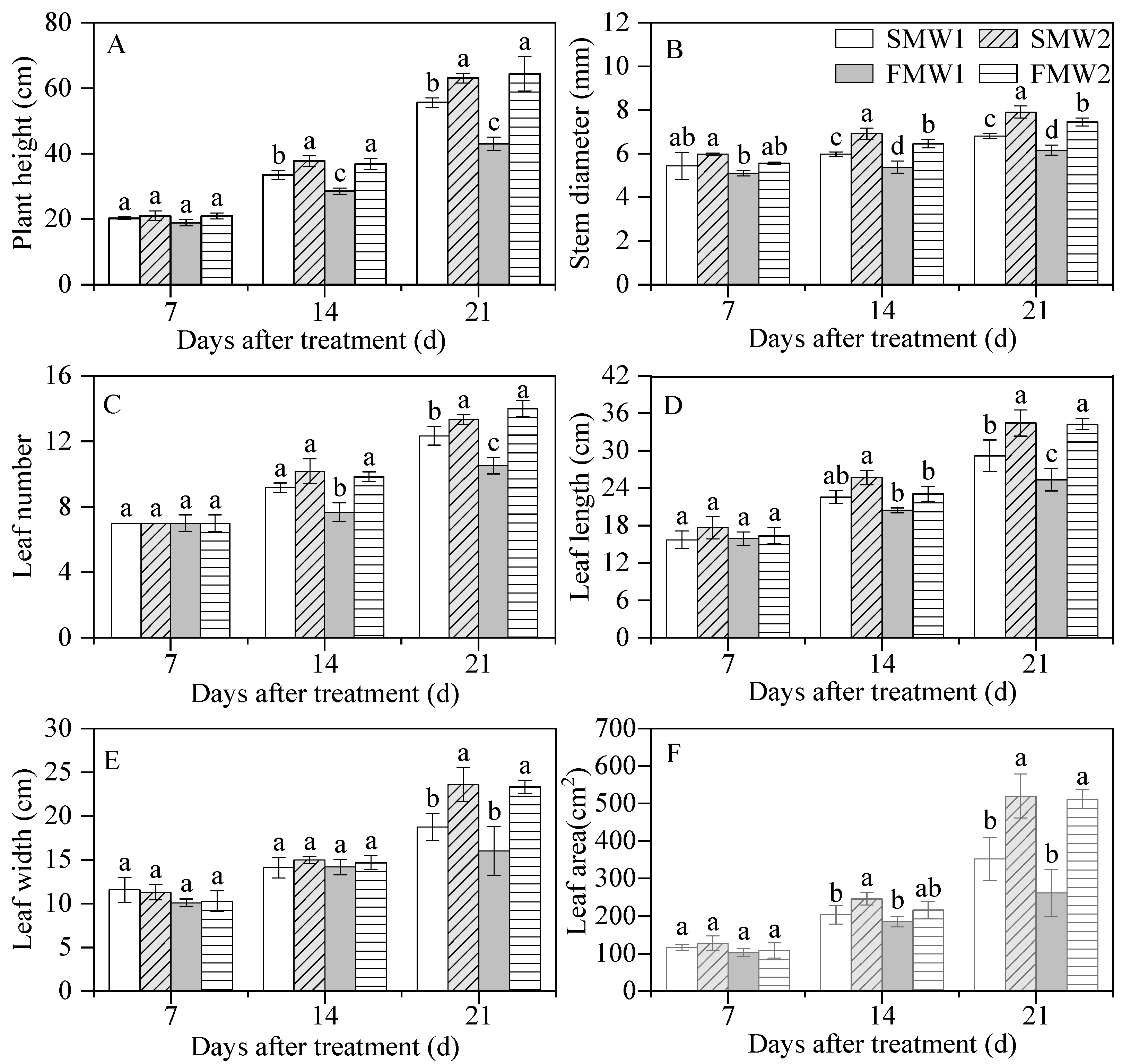
3.2. Biomass and Morphological Traits
3.3. LRWC, LREC, and Photosynthetic Pigments
3.4. Root Morphology, Root Activity, and Root/Shoot Ratio
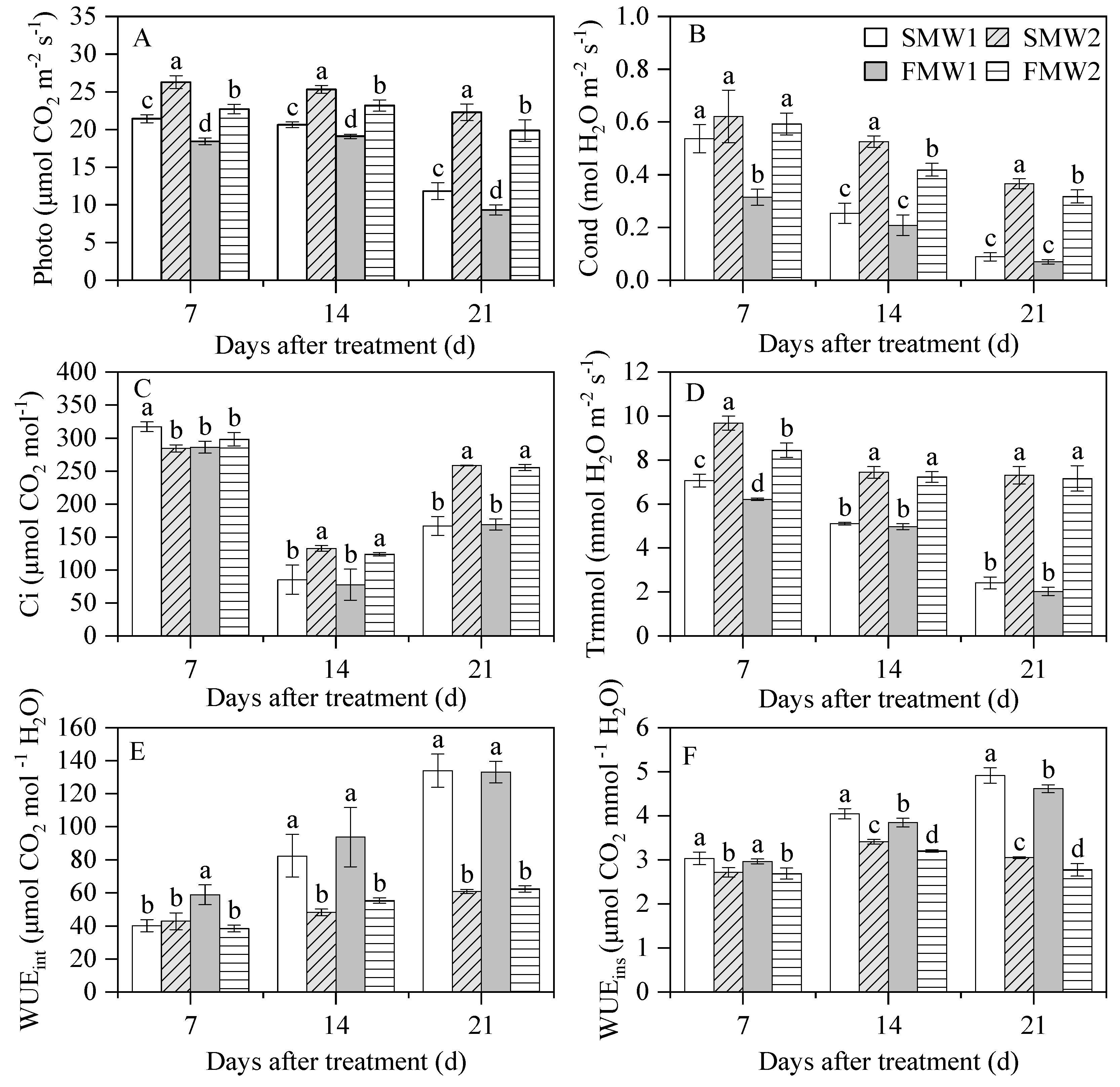
3.5. Leaf Gas Exchange and Leaf WUE
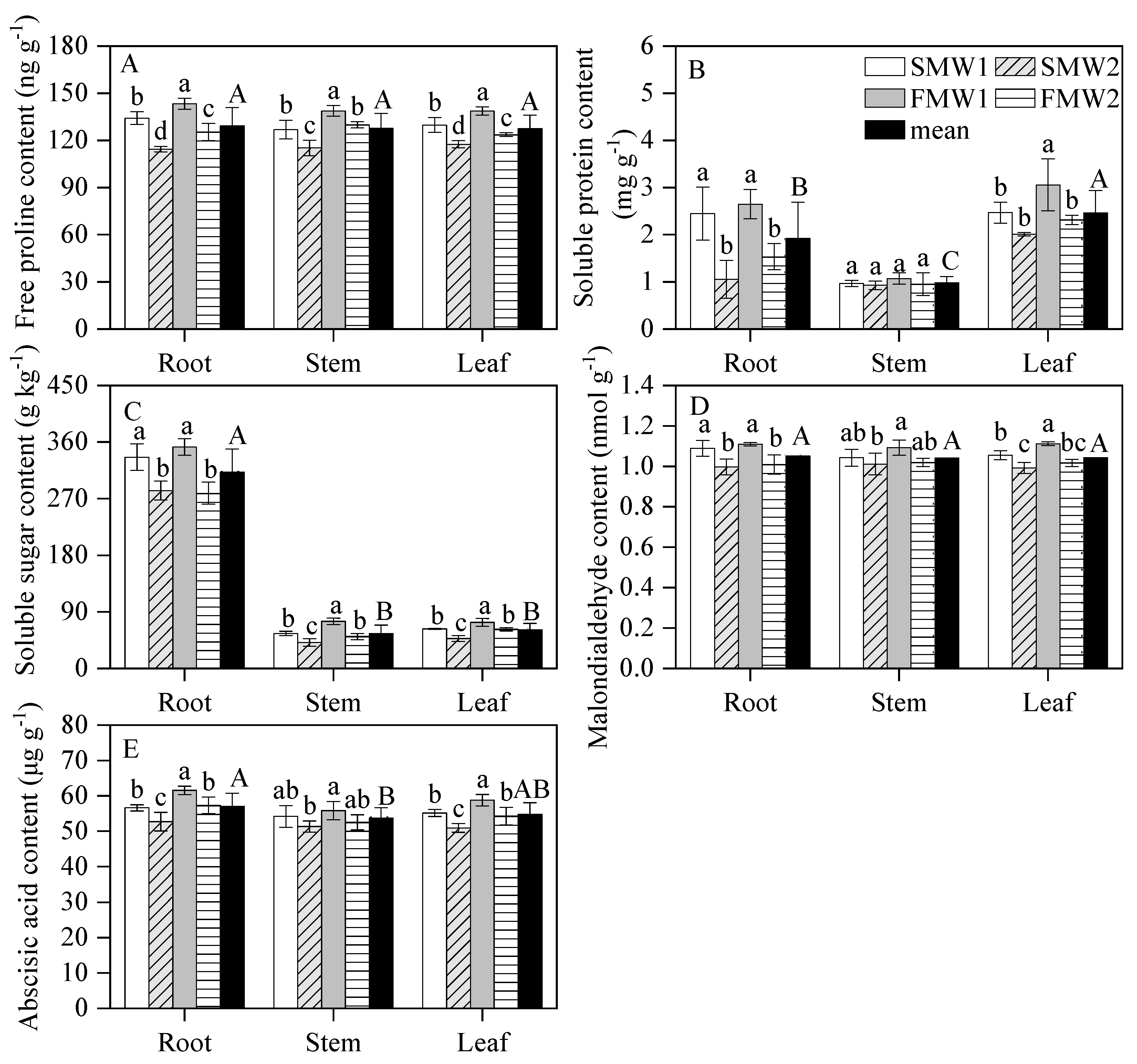
3.6. Osmotic Substances, Malondialdehyde and ABA
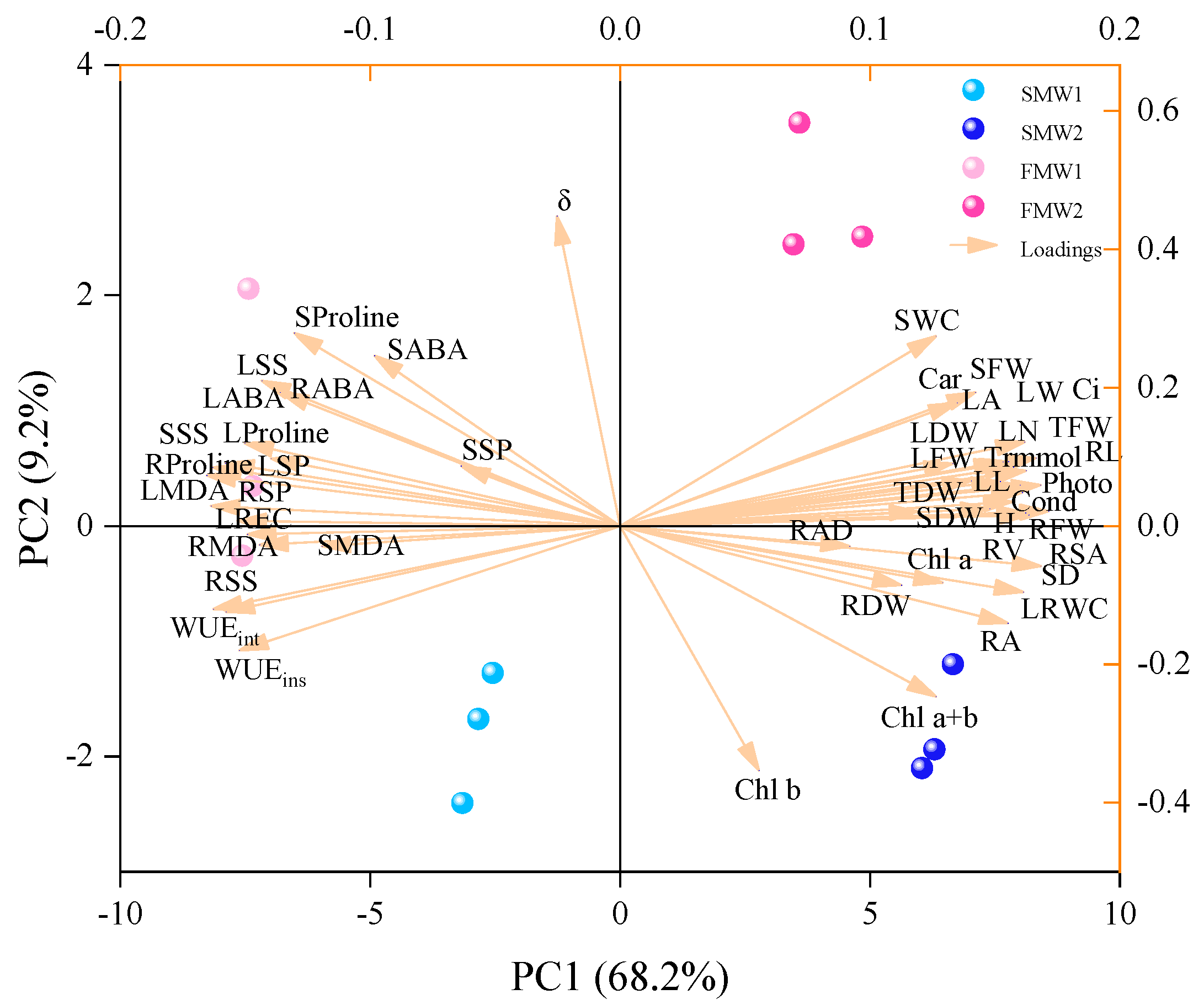
3.7. The Relations between Soil Water and Plant Parameters
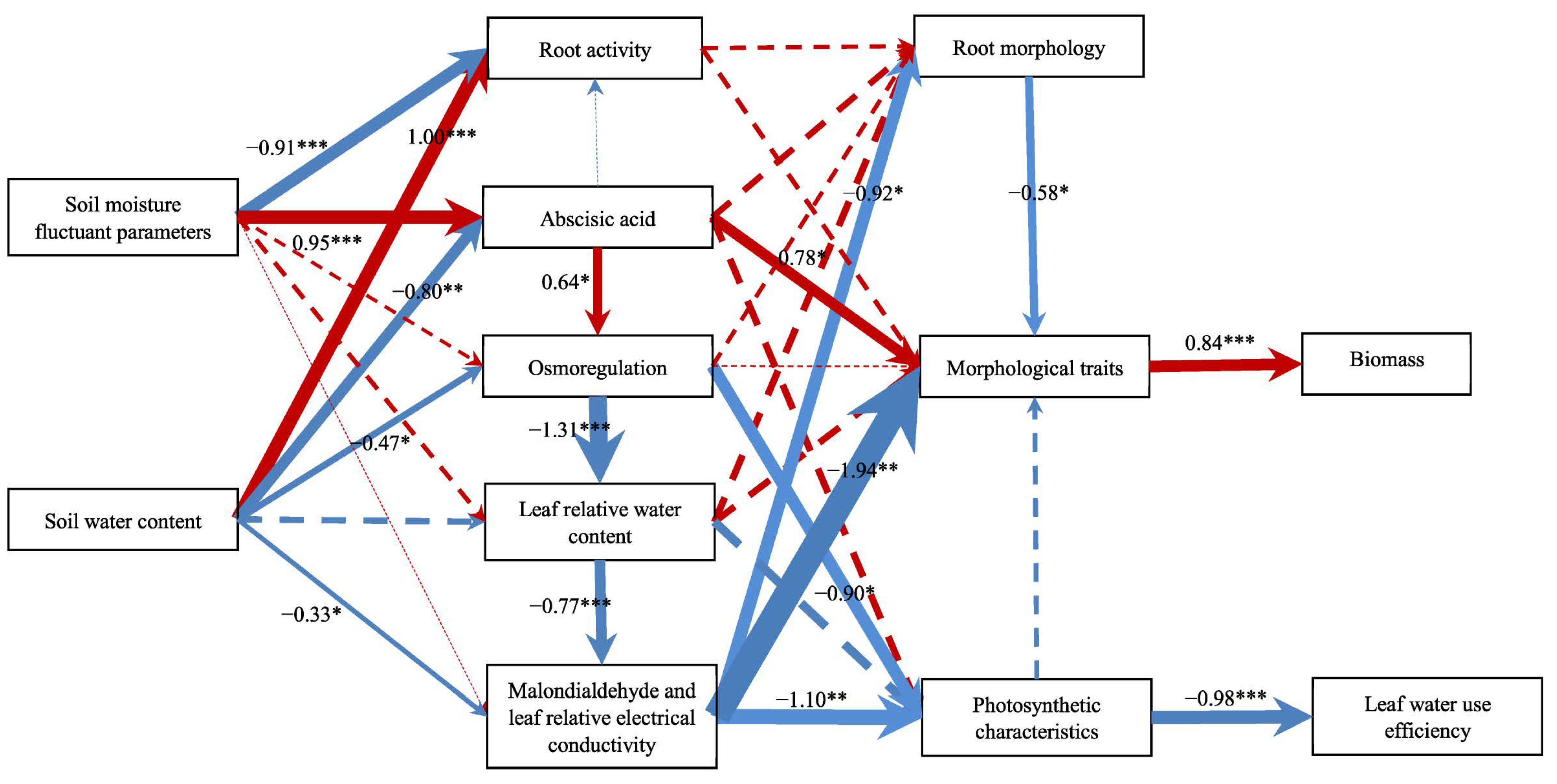
3.8. The Influence Path of Changing Soil Moisture on Water Use Efficiency
4. Discussion
4.1. Stable Soil Moisture Improves Plant Morphogenesis of Tomato Seedlings
4.2. Stable Soil Moisture Alleviates Soil Water Stress
4.3. Stable Soil Moisture Improves WUE of Tomato
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Nomenclature
| Abbreviation | Full Name |
| ABA | Abscisic acid |
| Ci | Intercellular CO2 concentration |
| Cond | Stomatal conductance |
| DW | Dry weight |
| FC | Field capacity |
| FM | Fluctuating soil moisture |
| FW | Fresh weight |
| LREC | Leaf relative electrical conductivity |
| LRWC | Leaf relative water content |
| NPI | Negative pressure irrigation |
| PCA | Principal component analysis |
| Photo | Net photosynthetic rate |
| PLS-PM | Partial least squares path |
| SM | Stable soil moisture |
| SWC | Soil water content |
| Trmmol | Transpiration rate |
| WUE | Water use efficiency |
| WUEins | Instantaneous water use efficiency |
| WUEint | Intrinsic water use efficiency |
| δ | Fluctuation coefficient |
References
- Farooq, M.; Hussain, M.; Ul-Allah, S.; Siddique, K.H.M. Physiological and Agronomic Approaches for Improving Water-Use Efficiency in Crop Plants. Agric. Water Manag. 2019, 219, 95–108. [Google Scholar] [CrossRef]
- Mega, R.; Abe, F.; Kim, J.-S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning Water-Use Efficiency and Drought Tolerance in Wheat Using Abscisic Acid Receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef]
- Islam, M.R.; Hu, Y.; Mao, S.; Mao, J.; Eneji, A.E.; Xue, X. Effectiveness of a Water-Saving Super-Absorbent Polymer in Soil Water Conservation for Corn (Zea mays L.) Based on Eco-Physiological Parameters. J. Sci. Food Agric. 2011, 91, 1998–2005. [Google Scholar] [CrossRef]
- Khapte, P.S.; Kumar, P.; Burman, U.; Kumar, P. Deficit Irrigation in Tomato: Agronomical and Physio-Biochemical Implications. Sci. Hortic. 2019, 248, 256–264. [Google Scholar] [CrossRef]
- Nangare, D.D.; Singh, Y.; Kumar, P.S.; Minhas, P.S. Growth, Fruit Yield and Quality of Tomato (Lycopersicon esculentum Mill.) as Affected by Deficit Irrigation Regulated on Phenological Basis. Agric. Water Manag. 2016, 171, 73–79. [Google Scholar] [CrossRef]
- Yang, P.; Bian, Y.; Long, H.; Drohan, P.J. Comparison of Emitters of Ceramic Tube and Polyvinyl Formal under Negative Pressure Irrigation on Soil Water Use Efficiency and Nutrient Uptake of Crown Daisy. Agric. Water Manag. 2020, 228, 105830. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, S.; Jia, S.; Du, F.; Qi, H.; Li, J.; Song, X.; Zhao, N.; Nie, L.; Fan, F. Precise Soil Water Control Using a Negative Pressure Irrigation System to Improve the Water Productivity of Greenhouse Watermelon. Agric. Water Manag. 2021, 258, 107144. [Google Scholar] [CrossRef]
- Zhang, J.; Ji, J.; Wang, P.; Long, H.; Wu, X. Molecular Mechanism of Negative Pressure Irrigation Inhibiting Root Growth and Improving Water Use Efficiency in Maize. Plant Soil 2022, 472, 127–143. [Google Scholar] [CrossRef]
- Li, S.; Tan, D.; Wu, X.; Degré, A.; Long, H.; Zhang, S.; Lu, J.; Gao, L.; Zheng, F.; Liu, X.; et al. Negative Pressure Irrigation Increases Vegetable Water Productivity and Nitrogen Use Efficiency by Improving Soil Water and NO3–-N Distributions. Agric. Water Manag. 2021, 251, 106853. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, S.; Song, Y.; Huaiyu, L. Negative Pressure Irrigation System Reduces Soil Nitrogen Loss for Lettuce during Greenhouse Production. Agronomy 2021, 11, 2380. [Google Scholar] [CrossRef]
- Yang, P.; Drohan, P.J.; Long, H.; Bian, Y.; Bryant, R.B. Negative Pressure Irrigation on Water Use Efficiency, Yield and Quality of Brassica chinensis L. J. Sci. Food Agric. 2021, 102, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, P.; Ji, J.; Long, H.; Wu, X. Transcriptome Analysis Reveals the Molecular Mechanism of Yield Increases in Maize under Stable Soil Water Supply. PLoS ONE 2021, 16, e0257756. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Wang, Z.; Zhu, G.; Yu, K.; Li, G.; Long, H. Stable Soil Moisture Improves the Water Use Efficiency of Maize by Alleviating Short-Term Soil Water Stress. Front. Plant Sci. 2022, 13, 833041. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Bai, J.; Yang, M.; Ma, E.; Yan, M.; Long, H.; Liu, J.; Li, L. Negative Pressure Irrigation for Greenhouse Crops in China: A Review. Agric. Water Manag. 2022, 264, 107497. [Google Scholar] [CrossRef]
- Li, F.; Li, Y.; Li, S.; Wu, G.; Niu, X.; Shen, A. Green Light Promotes Healing and Root Regeneration in Double-Root-Cutting Grafted Tomato Seedlings. Sci. Hortic. 2021, 289, 110503. [Google Scholar] [CrossRef]
- Zhou, T.; Wu, Z.; Wang, Y.; Su, X.; Qin, C.; Huo, H.; Jiang, F. Modelling Seedling Development Using Thermal Effectiveness and Photosynthetically Active Radiation. J. Integr. Agric. 2019, 18, 2521–2533. [Google Scholar] [CrossRef]
- Qu, Z.; Qi, X.; Liu, Y.; Liu, K.; Li, C. Interactive Effect of Irrigation and Polymer-Coated Potassium Chloride on Tomato Production in a Greenhouse. Agric. Water Manag. 2020, 235, 106149. [Google Scholar] [CrossRef]
- Pazzagli, P.T.; Weiner, J.; Liu, F. Effects of CO2 Elevation and Irrigation Regimes on Leaf Gas Exchange, Plant Water Relations, and Water Use Efficiency of Two Tomato Cultivars. Agric. Water Manag. 2016, 169, 26–33. [Google Scholar] [CrossRef]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef]
- Abbasi, S.; Sadeghi, A.; Safaie, N. Streptomyces Alleviate Drought Stress in Tomato Plants and Modulate the Expression of Transcription Factors ERF1 and WRKY70 Genes. Sci. Hortic. 2020, 265, 109206. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Ditta, A.; Suleman, M.; Ullah, M. Zinc-Induced Anti-Oxidative Defense and Osmotic Adjustments to Enhance Drought Stress Tolerance in Sunflower (Helianthus annuus L.). Environ. Exp. Bot. 2022, 193, 104682. [Google Scholar] [CrossRef]
- Yang, X.; Bornø, M.L.; Wei, Z.; Liu, F. Combined Effect of Partial Root Drying and Elevated Atmospheric CO2 on the Physiology and Fruit Quality of Two Genotypes of Tomato Plants with Contrasting Endogenous ABA Levels. Agric. Water Manag. 2021, 254, 106987. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Shahanzari, A.; Andersen, M.N.; Jacobsen, S.-E. ABA Regulated Stomatal Control and Photosynthetic Water Use Efficiency of Potato (Solanum tuberosum L.) during Progressive Soil Drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- Pérez-Pérez, J.G.; Navarro, J.M.; Robles, J.M.; Dodd, I.C. Prolonged Drying Cycles Stimulate ABA Accumulation in Citrus Macrophylla Seedlings Exposed to Partial Rootzone Drying. Agric. Water Manag. 2018, 210, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, F.; Wang, G.; Gao, Y.; Liang, Y.; Chen, J.; Si, Z.; Ramatshaba, T.S.; Zain, M.; Shafeeq-ur-rahman; Duan, A. Nitrous Oxide Emission from Winter Wheat Field as Responded to Irrigation Scheduling and Irrigation Methods in the North China Plain. Agric. Water Manag. 2019, 222, 367–374. [Google Scholar] [CrossRef]
- Siakou, M.; Bruggeman, A.; Eliades, M.; Zoumides, C.; Djuma, H.; Kyriacou, M.C.; Emmanouilidou, M.G.; Spyros, A.; Manolopoulou, E.; Moriana, A. Effects of Deficit Irrigation on ‘Koroneiki’ Olive Tree Growth, Physiology and Olive Oil Quality at Different Harvest Dates. Agric. Water Manag. 2021, 258, 107200. [Google Scholar] [CrossRef]
- Mazarei, R.; Soltani Mohammadi, A.; Ebrahimian, H.; Naseri, A.A. Temporal Variability of Infiltration and Roughness Coefficients and Furrow Irrigation Performance under Different Inflow Rates. Agric. Water Manag. 2021, 245, 106465. [Google Scholar] [CrossRef]
- North, G.B.; Nobel, P.S. Changes in Hydraulic Conductivity and Anatomy Caused by Drying and Rewetting Roots of Agave Deserti (Agavaceae). Am. J. Bot. 1991, 78, 906–915. [Google Scholar] [CrossRef]
- Salah, A.; Nwafor, C.C.; Han, Y.; Liu, L.; Rashid, M.; Batool, M.; El-Badri, A.M.; Cao, C.; Zhan, M. Spermidine and Brassinosteroid Regulate Root Anatomical Structure, Photosynthetic Traits and Antioxidant Defense Systems to Alleviate Waterlogging Stress in Maize Seedlings. S. Afr. J. Bot. 2022, 144, 389–402. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, Y.; Chen, G.; Yang, H.; Du, T. Water Use Efficiency Was Improved at Leaf and Yield Levels of Tomato Plants by Continuous Irrigation Using Semipermeable Membrane. Agric. Water Manag. 2018, 203, 430–437. [Google Scholar] [CrossRef]
- Wang, Z. Effects of Soil Moisture Temporal Variation on Growth, Physiology and Water Use Efficiency of Maize and Lettuces; Chinese Academy of Agricultural Sciences: Beijing, China, 2020. [Google Scholar]
- Li, G.; Bai, Y.; Wang, L.; Lu, Y.; Zhang, J.; Zhang, Y. Effects of Fertilizer under Different Dripline Spacings on Summer Maize in Northern China. Sci. Rep. 2021, 11, 18922. [Google Scholar] [CrossRef]
- Challabathula, D.; Analin, B.; Mohanan, A.; Bakka, K. Differential Modulation of Photosynthesis, ROS and Antioxidant Enzyme Activities in Stress-Sensitive and -Tolerant Rice Cultivars during Salinity and Drought upon Restriction of COX and AOX Pathways of Mitochondrial Oxidative Electron Transport. J. Plant Physiol. 2022, 268, 153583. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Altaf, M.M.; Khan, L.U.; Shahid, S.; Jahan, M.S. Melatonin Alleviates Salt Damage in Tomato Seedling: A Root Architecture System, Photosynthetic Capacity, Ion Homeostasis, and Antioxidant Enzymes Analysis. Sci. Hortic. 2021, 285, 110145. [Google Scholar] [CrossRef]
- Jabborova, D.; Ma, H.; Bellingrath-Kimura, S.D.; Wirth, S. Impacts of Biochar on Basil (Ocimum Basilicum) Growth, Root Morphological Traits, Plant Biochemical and Physiological Properties and Soil Enzymatic Activities. Sci. Hortic. 2021, 290, 110518. [Google Scholar] [CrossRef]
- Wang, B.; Li, P.-P.; Huang, C.-H.; Liu, G.-B.; Yang, Y.-F. Effects of Root Morphological Traits on Soil Detachment for Ten Herbaceous Species in the Loess Plateau. Sci. Total Environ. 2021, 754, 142304. [Google Scholar] [CrossRef]
- Shu, L.-Z.; Liu, R.; Min, W.; Wang, Y.; Hong-mei, Y.; Zhu, P.; Zhu, J. Regulation of Soil Water Threshold on Tomato Plant Growth and Fruit Quality under Alternate Partial Root-Zone Drip Irrigation. Agric. Water Manag. 2020, 238, 106200. [Google Scholar] [CrossRef]
- Gong, X.; Qiu, R.; Sun, J.; Ge, J.; Li, Y.; Wang, S. Evapotranspiration and Crop Coefficient of Tomato Grown in a Solar Greenhouse under Full and Deficit Irrigation. Agric. Water Manag. 2020, 235, 106154. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Liang, Y.; Kang, C.; Kaiser, E.; Kuang, Y.; Yang, Q.; Li, T. Red/Blue Light Ratios Induce Morphology and Physiology Alterations Differently in Cucumber and Tomato. Sci. Hortic. 2021, 281, 109995. [Google Scholar] [CrossRef]
- Guo, L.; Bornø, M.L.; Niu, W.; Liu, F. Biochar Amendment Improves Shoot Biomass of Tomato Seedlings and Sustains Water Relations and Leaf Gas Exchange Rates under Different Irrigation and Nitrogen Regimes. Agric. Water Manag. 2021, 245, 106580. [Google Scholar] [CrossRef]
- Sanchez, G.; Trinchera, L.; Russolollo, G. Plspm: Tools for Partial Least Squares Path Modeling (PLS-PM). R Packag. Version 0.4.9. 2015. Available online: https://rdrr.io/cran/plspm/ (accessed on 12 October 2022).
- Chen, J.; Song, D.; Luan, H.; Liu, D.; Wang, X.; Sun, J.; Gonzalo, M. Living and Dead Microorganisms in Mediating Soil Carbon Stocks under Long-Term Fertilization in a Rice-Wheat Rotation Study Site and Soil Sampling. Front. Microbiol. 2022, 13, 854216. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Gao, S.; Lu, Y.; Liao, Y.; Nie, J.; Cao, W. Co-Incorporation of Green Manure and Rice Straw Improves Rice Production, Soil Chemical, Biochemical and Microbiological Properties in a Typical Paddy Field in Southern China. Soil Tillage Res. 2020, 197, 104499. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, J.; Gao, Y.; Sun, Z.; Chen, S.; Yao, N.; Ma, H.; Feng, H.; Yu, Q.; He, J. Simulation of Plant Height of Winter Wheat under Soil Water Stress Using Modified Growth Functions. Agric. Water Manag. 2020, 232, 106066. [Google Scholar] [CrossRef]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J.; Wang, Y.; Yang, Q. Melatonin Improves Drought Resistance in Maize Seedlings by Enhancing the Antioxidant System and Regulating Abscisic Acid Metabolism to Maintain Stomatal Opening under PEG-Induced Drought. J. Plant Biol. 2021, 64, 299–312. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Nie, Z.; Sun, X. Effects of Molybdenum on Water Utilization, Antioxidative Defense System and Osmotic-Adjustment Ability in Winter Wheat (Triticum Aestivum) under Drought Stress. Plant Physiol. Biochem. 2014, 83, 365–374. [Google Scholar] [CrossRef]
- Hessini, K.; Martínez, J.P.; Gandour, M.; Albouchi, A.; Soltani, A.; Abdelly, C. Effect of Water Stress on Growth, Osmotic Adjustment, Cell Wall Elasticity and Water-Use Efficiency in Spartina Alterniflora. Environ. Exp. Bot. 2009, 67, 312–319. [Google Scholar] [CrossRef]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.; Fuggi, A.; Carillo, P. Hordeum Vulgare and Hordeum Maritimum Respond to Extended Salinity Stress Displaying Different Temporal Accumulation Pattern of Metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef]
- Yooyongwech, S.; Samphumphuang, T.; Tisarum, R.; Theerawitaya, C.; Cha-um, S. Arbuscular Mycorrhizal Fungi (AMF) Improved Water Deficit Tolerance in Two Different Sweet Potato Genotypes Involves Osmotic Adjustments via Soluble Sugar and Free Proline. Sci. Hortic. 2016, 198, 107–117. [Google Scholar] [CrossRef]
- Sánchez, F.J.; Manzanares, M.; de Andres, E.F.; Tenorio, J.L.; Ayerbe, L. Turgor Maintenance, Osmotic Adjustment and Soluble Sugar and Proline Accumulation in 49 Pea Cultivars in Response to Water Stress. Field Crop. Res. 1998, 59, 225–235. [Google Scholar] [CrossRef]
- Ghaffari, H.; Tadayon, M.R.; Bahador, M.; Razmjoo, J. Investigation of the Proline Role in Controlling Traits Related to Sugar and Root Yield of Sugar Beet under Water Deficit Conditions. Agric. Water Manag. 2021, 243, 106448. [Google Scholar] [CrossRef]
- Li, S.; Wang, X.; Liu, X.; Thompson, A.J.; Liu, F. Elevated CO2 and High Endogenous ABA Level Alleviate PEG-Induced Short-Term Osmotic Stress in Tomato Plants. Environ. Exp. Bot. 2022, 194, 104763. [Google Scholar] [CrossRef]
- da Silva, F.B.; Macedo, F.D.C.O.; Daneluzzi, G.S.; Capelin, D.; Silva, A.R.; Müller, C.; de Oliveira, R.F. Action Potential Propagation Effect on Gas Exchange of ABA-Mutant Microtomato after Re-Irrigation Stimulus. Environ. Exp. Bot. 2020, 178, 104149. [Google Scholar] [CrossRef]
- Tezara, W.; Domínguez, T.S.T.; Loyaga, D.W.; Ortiz, R.N.; Chila, V.H.R.; Ortega, M.J.B. Photosynthetic Activity of Oil Palm (Elaeis Guineensis) and Interspecific Hybrid Genotypes (Elaeis Oleifera × Elaeis Guineensis), and Response of Hybrids to Water Deficit. Sci. Hortic. 2021, 287, 110263. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Zeng, Y.; Guo, D.-J.; Singh, M.; Rajput, V.D.; Malviya, M.K.; Wei, K.-J.; Sharma, A.; Li, D.-P.; et al. Foliar Application of Silicon Boosts Growth, Photosynthetic Leaf Gas Exchange, Antioxidative Response and Resistance to Limited Water Irrigation in Sugarcane (Saccharum officinarum L.). Plant Physiol. Biochem. 2021, 166, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Farias, S.; Villalobos-Soublett, E.; Riveros-Burgos, C.; Zúñiga, M.; Ahumada-Orellana, L.E. Effect of Irrigation Cut-off Strategies on Yield, Water Productivity and Gas Exchange in a Drip-Irrigated Hazelnut (Corylus avellana L. Cv. Tonda Di Giffoni) Orchard under Semiarid Conditions. Agric. Water Manag. 2020, 238, 106173. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Test Period | Fluctuation Period | ||||
|---|---|---|---|---|---|---|
| The Range of SWC (%) | Mean SWC (%) | δ | The Range of SWC (%) | Mean SWC (%) | δ | |
| SMW1 | 18.7~29.1 | 22.6 (65% FC) | 0.03 | 18.7~24.2 | 19.9 (57% FC) | 0.02 |
| SMW2 | 23.0~29.0 | 25.5 (73% FC) | 0.02 | 23.0~26.0 | 24.1 (69% FC) | 0.01 |
| FMW1 | 19.0~28.8 | 24.0 (68% FC) | 0.08 | 19.0~26.3 | 22.8 (65% FC) | 0.07 |
| FMW2 | 20.9~29.8 | 26.6 (76% FC) | 0.11 | 20.9~29.8 | 26.1 (74% FC) | 0.10 |
| Treatment | LRWC (%) | LREC (%) | Chlorophyll a (μg·cm−2) | Chlorophyll b (μg·cm−2) | Carotenoids (μg·cm−2) | Chlorophyll a + b (μg·cm−2) |
|---|---|---|---|---|---|---|
| SMW1 | 69.42 ± 1.15 b | 17.06 ± 0.53 ab | 17.70 ± 1.33 b | 6.10 ± 1.28 a | 1.30 ± 0.15 b | 23.80 ± 0.31 b |
| SMW2 | 74.27 ± 0.94 a | 15.25 ± 1.22 b | 20.77 ± 1.35 a | 6.97 ± 1.20 a | 2.39 ± 0.47 a | 27.73 ± 0.67 a |
| FMW1 | 62.58 ± 2.06 c | 18.50 ± 0.64 a | 17.36 ± 1.73 b | 5.05 ± 1.06 a | 1.37 ± 0.45 b | 22.41 ± 2.68 b |
| FMW2 | 71.21 ± 0.21 b | 15.19 ± 1.39 b | 19.18 ± 0.54 ab | 4.90 ± 0.30 a | 2.62 ± 0.11 a | 24.08 ± 0.52 b |
| Treatment | Root Length (cm) | Root Surface Area (cm2) | Root Volume (cm3) | Average Root Diameter (mm) | Specific Root Length (m·g−1) | Root Activity (μg·h−1·g−1) | Root/Shoot Ratio (g·g−1) |
|---|---|---|---|---|---|---|---|
| SMW1 | 442.59 ± 119.45 b | 39.99 ± 8.50 b | 0.56 ± 0.23 bc | 0.67 ± 0.12 a | 12.71 ± 7.35 ab | 81.63 ± 8.07 c | 0.06 ± 0.01 a |
| SMW2 | 729.79 ± 101.54 a | 60.93 ± 6.19 a | 0.96± 0.23 a | 0.86 ± 0.18 a | 15.37 ± 1.50 ab | 140.53 ± 3.35 a | 0.07 ± 0.00 a |
| FMW1 | 254.00 ± 59.31 b | 26.00 ± 4.93 c | 0.37± 0.05 c | 0.65 ± 0.13 a | 8.18 ± 2.21 b | 71.05 ± 1.18 d | 0.06 ± 0.02 a |
| FMW2 | 690.71 ± 125.55 a | 55.55 ± 6.28 a | 0.92± 0.19 ab | 0.81 ± 0.22 a | 17.07 ± 1.93 a | 99.27 ± 6.76 b | 0.05 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Long, H.; Zhang, R.; Drohan, P.J.; Xu, A.; Niu, L. Stable Soil Moisture Alleviates Water Stress and Improves Morphogenesis of Tomato Seedlings. Horticulturae 2023, 9, 391. https://doi.org/10.3390/horticulturae9030391
Li G, Long H, Zhang R, Drohan PJ, Xu A, Niu L. Stable Soil Moisture Alleviates Water Stress and Improves Morphogenesis of Tomato Seedlings. Horticulturae. 2023; 9(3):391. https://doi.org/10.3390/horticulturae9030391
Chicago/Turabian StyleLi, Ge, Huaiyu Long, Renlian Zhang, Patrick J. Drohan, Aiguo Xu, and Li Niu. 2023. "Stable Soil Moisture Alleviates Water Stress and Improves Morphogenesis of Tomato Seedlings" Horticulturae 9, no. 3: 391. https://doi.org/10.3390/horticulturae9030391
APA StyleLi, G., Long, H., Zhang, R., Drohan, P. J., Xu, A., & Niu, L. (2023). Stable Soil Moisture Alleviates Water Stress and Improves Morphogenesis of Tomato Seedlings. Horticulturae, 9(3), 391. https://doi.org/10.3390/horticulturae9030391