
Physiology and Application of Gibberellins in Postharvest Horticultural Crops


Abstract
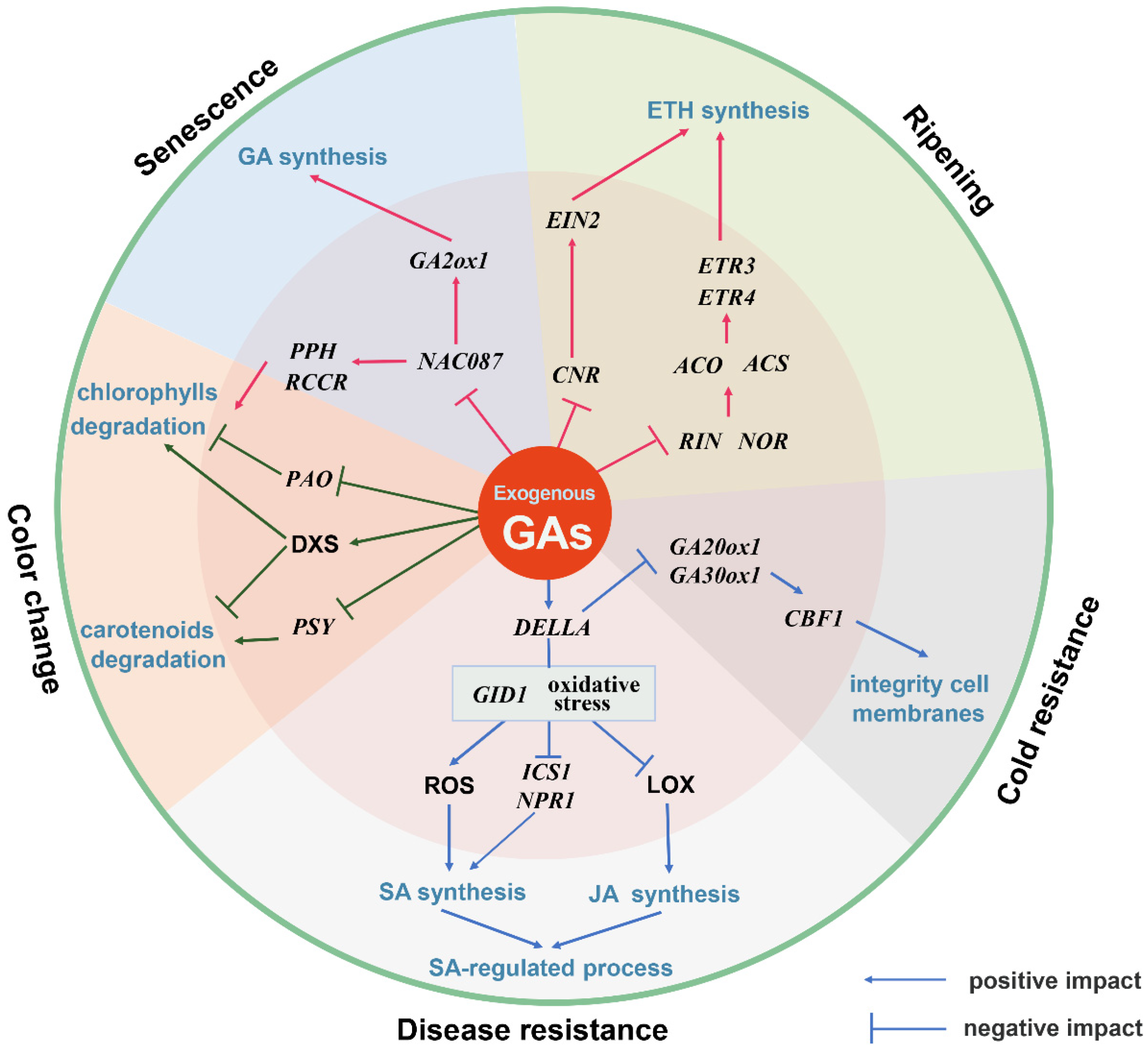
:1. Introduction
2. Gibberellins for Regulating the Ripening and Senescence Process
3. Gibberellins for Exterior Quality Control of Horticultural Crops
4. Gibberellins for Improving the Internal Quality of Horticultural Crops

{kind=link}
| Horticultural Varieties | Main Points | Concentration of GAs | References |
|---|---|---|---|
| Apple (Malus pumila Mill.) | GA3 spraying retards the decrease in soluble solids content in fruits. | 100 mg L−1 GA3 | Liu et al., 2022 [18] |
| Banana (Musa nana Lour.) | GA3 and CPPU treatments reduce water loss in fruits. | 50 mg L−1 GA3 + 10 mg L−1 CPPU | Huang et al., 2012 [36] |
| ‘rabbit eye’ blueberry (Vaccinium virgatum Ait.) | GA3 spraying retards the decrease in soluble solids content of fruits. | 500 mg L−1 GA3 | Zang et al., 2016 [33] |
| ‘rabbit eye’ blueberry (Vaccinium virgatum Ait.) | GA3 spraying retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 500 mg L−1 GA3 | Zang et al., 2016 [33] |
| Broccoli (Brassica capitata var. italica) | GA3 and CPPU treatments reduce water loss. | 50 mg L−1 GA3 + 10 mg L−1 CPPU | Huang et al., 2012 [36] |
| Sweet cherry (Cerasus pseudocerasus (Lindl.) G. Don) | GA3 spraying retards fruit weight loss. | 30 mg L−1 GA3 | Burhan et al., 2022 [31] |
| Sweet cherry (Cerasus pseudocerasus (Lindl.) G. Don) | GA3 spraying raises fruit acidity levels. | 60 mg L−1 GA3 | Ozkan et al., 2016 [34] |
| ‘Barhee’ date (Ziziphus jujuba Mill.) | GA3 spraying retards the decline of Vc content in fruits. | 50 ppm GA3 | Awad et al., 2012 [37] |
| Garlic (Allium sativum L.) | GA3 injection increases soluble protein content of fruits. | 1 mM GA3 | Liu et al., 2019 [7] |
| ‘Cabernet Sauvignon’ grape (Vitis vinifera L.) | GA3 spraying increases phenol content of fruits. | 15 mg L−1 GA3 | Song et al., 2023 [41] |
| Kiwifruit (Actinidia chinensis Planch.) | GA3 spraying retards the decline of fruit hardness and soluble solids content. | 0.3 g L−1 GA3 | Yang et al., 2023 [28] |
| Welsh onion (Allium cepa L.) | GA3 spraying increases soluble protein content of fruits. | 40 ppm GA3 | Yamazaki et al., 2015 [40] |
| ‘Nanguo’ pear (Pyrus spp) | GA3 soaking retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 100 mg L−1 GA3 | Martínez-Romero et al., 2000 [39] |
| Plum (Prunus salicina Lindl.) | GA3 spraying retards the decrease in soluble solids content of fruits and reduces titratable acidity at harvest. | 200 mg L−1 GA3 | Barac et al., 2022 [32] |
| Plum (Prunus salicina Lindl.) | GA3 spraying retards the reduction of soluble solids and titratable acid content in fruits. | 50 mg L−1 GA3 | Erogul et al., 2015 [35] |
| Plum (Prunus salicina Lindl.) | GA3 soaking retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 200 μL L−1 GA3 | Li et al., 2006 [38] |
| Plum (Prunus salicina Lindl.) | GA3 spraying retards the decline of ascorbic acid concentration and the increase of total phenolic content of fruits. | 50 mg L−1 GA3 | Erogul et al., 2015 [35] |
| ‘Angelino’ plum (Prunus salicina Lindl.) | GA3 spraying retards fruit weight loss and decay. | 50 mg L−1 GA3 | Erogul et al., 2016 [29] |
| ‘BlackStar’ plum (Prunus salicina Lindl.) | GA3 spraying retards fruit weight loss and decay. | 50 ppm GA3 | Harman et al., 2016 [30] |
| ‘Obilnaja’ plum (Prunus salicina Lindl.) | GA3 spraying retards fruit weight loss and decay. | 50 ppm GA3 | Harman et al., 2016 [30] |
5. Gibberellins for Improving Cold Resistance in Horticultural Crops
6. Gibberellins Regulate Disease Resistance in Horticultural Crops
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin Localization and Transport in Plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, J.; Suter, P.J. The occurrence of gibberellin A1 in higher plants: Isolation from the seed of runner bean (Phaseolus multiflorus). Naturwissenschaften 1958, 45, 46. [Google Scholar] [CrossRef]
- MacMillan, J. Occurrence of Gibberellins in Vascular Plants, Fungi, and Bacteria. J. Plant Growth Regul. 2001, 20, 387–442. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Zhu, Z.; Liang, H.; Chen, G.; Li, F.; Wang, Y.; Liao, C.; Hu, Z. The bHLH transcription factor SlPRE2 regulates tomato fruit development and modulates plant response to gibberellin. Plant Cell Rep. 2019, 38, 1053–1064. [Google Scholar] [CrossRef]
- Li, H.; Wu, H.; Qi, Q.; Li, H.; Li, Z.; Chen, S.; Ding, Q.; Wang, Q.; Yan, Z.; Gai, Y.; et al. Gibberellins Play a Role in Regulating Tomato Fruit Ripening. Plant Cell Physiol. 2019, 60, 1619–1629. [Google Scholar] [CrossRef]
- Liu, H.; Deng, R.; Huang, C.; Cheng, Z.; Meng, H. Exogenous gibberellins alter morphology and nutritional traits of garlic (Allium sativum L.) bulb. Sci. Hortic. 2019, 246, 298–306. [Google Scholar] [CrossRef]
- Xie, Y.; Onik, J.C.; Hu, X.; Duan, Y.; Lin, Q. Effects of (S)-Carvone and Gibberellin on Sugar Accumulation in Potatoes during Low Temperature Storage. Molecules 2018, 23, 3118. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.J.; Tan, G.F.; Zhou, W.Q.; Wang, G.L. Gibberellin and the plant growth retardant Paclobutrazol altered fruit shape and ripening in tomato. Protoplasma 2020, 257, 853–861. [Google Scholar] [CrossRef]
- Fan, Z.Q.; Wei, W.; Tan, X.L.; Shan, W.; Kuang, J.F.; Lu, W.J.; Su, X.G.; Lakshmanan, P.; Lin, H.T.; Chen, J.Y. A NAC transcription factor BrNAC087 is involved in gibberellin-delayed leaf senescence in Chinese flowering cabbage. Postharvest Biol. Technol. 2021, 181, 111673. [Google Scholar] [CrossRef]
- Zhao, H.; Lv, W.J.; Fan, Y.L.; Li, H.Q. Gibberellic acid enhances postharvest toon sprout tolerance to chilling stress by increasing the antioxidant capacity during the short-term cold storage. Sci. Hortic. 2018, 237, 184–191. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, C.; Cheng, Y.; Ruan, M.; Ye, Q.; Wang, R.; Yao, Z.; Wan, H. Molecular Evolution and Functional Divergence of Stress-Responsive Cu/Zn Superoxide Dismutases in Plants. Int. J. Mol. Sci. 2022, 23, 7082. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.K.; Mahmud, S.; Dutta, A.K.; Sarkar, S.; Laboni, A.A.; Hossain, M.S.; Nagata, A.; Karmaker, P.; Razu, M.H.; Kazi, T.; et al. Volatile compounds of Bacillus pseudomycoides induce growth and drought tolerance in wheat (Triticum aestivum L.). Sci. Rep. 2022, 12, 19137. [Google Scholar] [CrossRef]
- Silva, F.V.M.; Sulaiman, A. Control of Enzymatic Browning in Strawberry, Apple, and Pear by Physical Food Preservation Methods: Comparing Ultrasound and High-Pressure Inactivation of Polyphenoloxidase. Foods 2022, 11, 1942. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.K.; Wang, C.Y.; Gross, K.C.; Smith, D.L. Reduction of chilling injury and transcript accumulation of heat shock proteins in tomato fruit by methyl jasmonate and methyl salicylate. Plant Sci. 2001, 161, 1153–1159. [Google Scholar] [CrossRef]
- Li, J.; Cao, J.; Wang, C.; Hao, N.; Zhang, X.; Liu, M.; Wu, T. Research Progress on the Leaf Morphology, Fruit Development and Plant Architecture of the Cucumber. Plants 2022, 11, 2128. [Google Scholar] [CrossRef]
- Cruz-Castillo, J.G.; Woolley, D.J. Pedicel weight and length do not affect kiwifruit size. Eur. J. Hortic. Sci. 2006, 71, 272–276. [Google Scholar]
- Liu, C.; Xiao, P.; Jiang, F.; Wang, S.; Liu, Z.; Song, G.; Li, W.; Lv, T.; Li, J.; Wang, D.; et al. Exogenous gibberellin treatment improves fruit quality in self-pollinated apple. Plant Physiol. Biochem. 2022, 174, 11–21. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, R. Gibberellic acid influences the production of malformed and button berries, and fruit yield and quality in strawberry (Fragaria×ananassa Duch.). Sci. Hortic. 2009, 119, 430–433. [Google Scholar] [CrossRef]
- Liu, B.; Liu, X.; Yang, S.; Chen, C.; Xue, S.; Cai, Y.; Wang, D.; Yin, S.; Gai, X.; Ren, H. Silencing of the gibberellin receptor homolog, CsGID1a, affects locule formation in cucumber (Cucumis sativus) fruit. New Phytol. 2016, 210, 551–563. [Google Scholar] [CrossRef]
- Wang, H.T.; Ou, L.Y.; Chen, T.A.; Kuan, Y.C. Refrigeration, forchlorfenuron, and gibberellic acid treatments differentially regulate chlorophyll catabolic pathway to delay yellowing of broccoli. Postharvest Biol. Technol. 2023, 197, 112221. [Google Scholar] [CrossRef]
- Barry, G.H.; Le Roux, S. Preharvest Foliar Sprays of Prohexadione-calcium, a Gibberellin-biosynthesis Inhibitor, Induce Chlorophyll Degradation and Carotenoid Synthesis in Citrus Rinds. Hortscience 2010, 45, 242–247. [Google Scholar] [CrossRef]
- Rehman, M.; Singh, Z.; Khurshid, T. Pre-harvest spray application of prohexadione-calcium and paclobutrazol improves rind colour and regulates fruit quality in M7 Navel oranges. Sci. Hortic. 2018, 234, 87–94. [Google Scholar] [CrossRef]
- Alos, E.; Cercos, M.; Rodrigo, M.J.; Zacarias, L.; Talon, M. Regulation of color break in citrus fruits. Changes in pigment profiling and gene expression induced by gibberellins and nitrate, two ripening retardants. J. Agric. Food Chem. 2006, 54, 4888–4895. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, K.; Maoz, I.; Lapidot, O.; Kochanek, B.; Butnaro, Y.; Shlisel, M.; Lerno, L.; Ebeler, S.E.; Lichter, A. Effects of gibberellin and cytokinin on phenolic and volatile composition of Sangiovese grapes. Sci. Hortic. 2022, 295, 111672. [Google Scholar] [CrossRef]
- Qu, S.S.; Li, M.M.; Wang, G.; Zhu, S.J. Application of ABA and GA3 alleviated browning of litchi (Litchi chinensis Sonn.) via different strategies. Postharvest Biol. Technol. 2021, 181, 111672. [Google Scholar] [CrossRef]
- Keawmanee, N.; Ma, G.; Zhang, L.C.; Yahata, M.; Murakami, K.; Yamamoto, M.; Kojima, N.; Kato, M. Exogenous gibberellin induced regreening through the regulation of chlorophyll and carotenoid metabolism in Valencia oranges. Plant Physiol. Biochem. 2022, 173, 14–24. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Li, X.; Wu, R.; Zhang, X.; Fan, X.; Li, G.; Gong, H.; Yin, X.; Zhang, A. The mechanism of gibberellins treatment suppressing kiwifruit postharvest ripening processes by transcriptome analysis. Postharvest Biol. Technol. 2023, 198, 112223. [Google Scholar] [CrossRef]
- Erogul, D.; Sen, F. The effect of preharvest gibberellic acid applications on fruit quality of ʽAngelinoʼ plums during storage. Sci. Hortic. 2016, 202, 111–116. [Google Scholar] [CrossRef]
- Harman, Y.; Sen, F. The effect of different concentrations of pre-harvest gibberellic acid on the quality and durability of ‘Obilnaja’ and ‘Black Star’ plum varieties. Food Sci. Technol. 2016, 36, 362–368. [Google Scholar] [CrossRef]
- Burhan, O.; Erdal, A.; Saraçoğlu, O.; Karakaya, O.; Gun, S. Effects of GA 3, CACl 2 and Modified Atmosphere Packaging (MAP) Applications on Fruit Quality of Sweet Cherry at Cold Storage. Int. J. Fruit Sci. 2022, 22, 696–710. [Google Scholar] [CrossRef]
- Barac, G.; Mastilovic, J.; Kevresan, Z.; Milic, B.; Kovac, R.; Milovic, M.; Kalajdzic, J.; Bajic, A.; Magazin, N.; Keserovic, Z. Effects of Plant Growth Regulators on Plum (Prunus domestica L.) Grown on Two Rootstocks at Harvest and at the Postharvest Period. Horticultuae 2022, 8, 621. [Google Scholar] [CrossRef]
- Zang, Y.X.; Chun, I.J.; Zhang, L.L.; Hong, S.B.; Zheng, W.W.; Xu, K. Effect of gibberellic acid application on plant growth attributes, return bloom, and fruit quality of rabbiteye blueberry. Sci. Hortic. 2016, 200, 13–18. [Google Scholar] [CrossRef]
- Ozkan, Y.; Ucar, M.; Yildiz, K.; Ozturk, B. Pre-harvest gibberellic acid (GA3) treatments play an important role on bioactive compounds and fruit quality of sweet cherry cultivars. Sci. Hortic. 2016, 211, 358–362. [Google Scholar] [CrossRef]
- Erogul, D.; Sen, F. Effects of gibberellic acid treatments on fruit thinning and fruit quality in Japanese plum (Prunus salicina Lindl.). Sci. Hortic. 2015, 186, 137–142. [Google Scholar] [CrossRef]
- Huang, H.; Jiang, Y. Effect of plant growth regulators on banana fruit and broccoli during storage. Sci. Hortic. 2012, 145, 62–67. [Google Scholar] [CrossRef]
- Awad, M.A.; Al-Qurashi, A.D. Gibberellic acid spray and bunch bagging increase bunch weight and improve fruit quality of ‘Barhee’ date palm cultivar under hot arid conditions. Sci. Hortic. 2012, 138, 96–100. [Google Scholar] [CrossRef]
- Li, H.; Jiang, Y.; Li, J. Use of GA3 to inhibit flesh browning development of plum fruit during storage at low temperature. Eur. J. Hortic. Sci. 2006, 71, 231–235. [Google Scholar]
- Martínez-Romero, D.; Valero, D.; Serrano, M.; Burló, F.; Carbonell-Barrachina, A.; Burgos, L.; Riquelme, F. Exogenous Polyamines and Gibberellic Acid Effects on Peach (Prunus persica L.) Storability Improvement. J. Food Sci. 2000, 65, 288–294. [Google Scholar] [CrossRef]
- Yamazaki, H.; Shiraiwa, N.; Itai, A.; Honda, I. Involvement of Gibberellins in the Regulation of Tillering in Welsh Onion (Allium fistulosum L.). Hortic. J. 2015, 84, 334–341. [Google Scholar] [CrossRef]
- Song, J.Q.; Zhang, A.; Gao, F.; Liang, H.Z.; Li, M.Q.; Zhang, J.; Wang, G.J.; Qu, H.G.; Cheng, S.W.; Ruan, S.L.; et al. Modification of wine phenolic profiles by gibberellic acid application in ‘Cabernet Gernischt’ grapevines before anthesis. J. Sci. Food Agric. 2023, 103, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Ding, Y.; Zhao, J.; Nie, Y.; Zhang, Y.; Sheng, J.; Tang, X. Effects of Postharvest Gibberellic Acid Treatment on Chilling Tolerance in Cold-Stored Tomato (Solanum lycopersicum L.) Fruit. Food Bioprocess Technol. 2016, 9, 1202–1209. [Google Scholar] [CrossRef]
- Ding, Y.; Zhao, J.; Nie, Y.; Fan, B.; Wu, S.; Zhang, Y.; Sheng, J.; Shen, L.; Zhao, R.; Tang, X. Salicylic-Acid-Induced Chilling- and Oxidative-Stress Tolerance in Relation to Gibberellin Homeostasis, C-Repeat/Dehydration-Responsive Element Binding Factor Pathway, and Antioxidant Enzyme Systems in Cold-Stored Tomato Fruit. J. Agric. Food Chem. 2016, 64, 8200–8206. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sheng, J.; Li, S.; Nie, Y.; Zhao, J.; Zhu, Z.; Wang, Z.; Tang, X. The role of gibberellins in the mitigation of chilling injury in cherry tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2015, 101, 88–95. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Li, S.; Li, Y.; Wang, Y. Trichothecium roseum infection promotes ripening of harvested muskmelon fruits and induces the release of specific volatile compound. J. Plant Pathol. 2019, 101, 529–538. [Google Scholar] [CrossRef]
- Xue, H.; Bi, Y.; Sun, Y.; Hussain, R.; Wang, H.; Zhang, S.; Zhang, R.; Long, H.; Nan, M.; Cheng, X.; et al. Acetylsalicylic acid treatment reduce Fusarium rot development and neosolaniol accumulation in muskmelon fruit. Food Chem. 2019, 289, 278–284. [Google Scholar] [CrossRef]
- Ma, L.Y.; Zhao, L.; Bi, Y.; An, L. Efficacy of erythromycin and daidzein with pyrimethanil for postharvest disease control in melon. Henan Agric. Sci. 2005, 78–81. [Google Scholar] [CrossRef]
- Jia, Z.S.; Wang, Z.H.; Wang, W.H.; Jiang, Y.B. Relationship between the occurrence of black skin disease and related physiological indicators during storage of Dangshan pear fruit. North China J. Agric. 2021, 36, 203–209. [Google Scholar]
- Ma, L.; Li, H.X.; Wang, Y.; Li, Y.C.; Bi, Y. The effect of pre-harvest gibberellins on ‘black peel’ pears. Effect of pre-harvest gibberellin spray on the prevention and control of black skin disease in ‘apple pear’ during storage at room temperature. China Fruit Tree 2018, 193, 1–4+10. [Google Scholar] [CrossRef]
- Eshel, D.; Ben-Arie, R.; Dinoor, A.; Prusky, D. Resistance of Gibberellin-Treated Persimmon Fruit to Alternaria alternata Arises from the Reduced Ability of the Fungus to Produce Endo-1,4-beta-Glucanase. Phytopathology 2000, 90, 1256–1262. [Google Scholar] [CrossRef]
- Biton, E.; Kobiler, I.; Feygenberg, O.; Yaari, M.; Friedman, H.; Prusky, D. Control of alternaria black spot in persimmon fruit by a mixture of gibberellin and benzyl adenine, and its mode of action. Postharvest Biol. Technol. 2014, 94, 82–88. [Google Scholar] [CrossRef]
- Wu, S.J. Mechanism of Action of Gibberellin and Brassinolide in the Regulation of Early Blight Resistance in Postharvest Tomato Fruits. Master’s thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2018. [Google Scholar]
- De Bruyne, L.; Hofte, M.; De Vleesschauwer, D. Connecting growth and defense: The emerging roles of brassinosteroids and gibberellins in plant innate immunity. Mol. Plant 2014, 7, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D. Salicylic acid signaling in disease resistance. Plant Sci. 2014, 228, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, P.; Oh, Y.; Panthee, D.R. Current Status of Early Blight Resistance in Tomato: An Update. Int. J. Mol. Sci. 2017, 18, 2019. [Google Scholar] [CrossRef]

| Horticultural Varieties | Main Points | Concentration | References |
|---|---|---|---|
| Cabbage (Brassica rapa var. glabra Regel) | GA3 spraying retarded fruit senescence. | 100 μM GA3 | Fan et al., 2021 [10] |
| Garlic (Allium sativum L.) | GA3 injection improves the nutritional traits of fruits. | 1 mM GA3 | Liu et al., 2019 [7] |
| Potato (Solanum tuberosum L.) | GA spraying accumulates tuber sugar nutrients. | 0.015 g L−1 GA | Xie et al., 2018 [8] |
| Tomato (Solanum lycopersicum L.) | GA3 spraying affects the size of fruits. | 50 μM GA3 | Zhu et al.,2019 [5] |
| Tomato (Solanum lycopersicum L.) | GA3 injection delays color change in fruit at ripening stage. | 0.1 mM GA3 | Li et al., 2019 [7] |
| Toona (Toona sinensis (A. Juss.) Roem.) | GA3 soaking retards fruit senescence. | 100 mg L–1 GA3 | Zhao et al., 2018 [11] |
| Horticultural Varieties | Main Points | Concentration of GAs | References |
|---|---|---|---|
| Apple (Malus pumila Mill.) | GA3 spraying increases fruit stem length and improves fruit shape index. | 100 mg L−1 GA3 | Liu et al., 2022 [18] |
| Apple (Malus pumila Mill.) | GA3 spraying increases fruit weight and improves fruit shape index. | 100 mg L−1 GA3 | Liu et al., 2022 [18] |
| Broccoli (Brassica capitata var. italica) | GA3 soaking retards the degradation of chlorophyll in fruit peel and slows down the color change of fruit skin. | 10 mg L−1 GA3 | Wang et al., 2023 [21] |
| Citrus (Citrus reticulata Blanco) | ProCa spraying reduces chlorophyll and increases carotenoid concentrations in fruits. | 400 mg·L−1 ProCa | Barry et al., 2010 [22] |
| Citrus (Citrus reticulata Blanco) | PBZ spraying affects fruit color. | 1500 mg L−1 PBZ | Rehman et al., 2018 [23] |
| Citrus (Citrus reticulata Blanco) | GA3 spraying affects fruit color by influencing enzyme activity. | 60 mg L−1 GA3 | Alos et al., 2006 [24] |
| Cucumber (Cucumis sativus Linn.) | GA3 spraying regulates fruit shape. | 50 μM GA3 | Liu et al., 2016 [20] |
| Grape (Vitis vinifera L.) | GA3 spraying reduces flavanol and anthocyanin content and slows down the color change of fruit skin. | 20 ppm GA3 | Tyagi et al., 2022 [25] |
| ‘Hayward’ kiwifruit (Actinidia chinensis Planch.) | GA3 soaking increases fruit stem length and improves fruit shape index. | 100 mg L–1 GA3 | Cruz-Castillo et al., 2006 [17] |
| Lychee (Litchi chinensis Sonn.) | GA3 soaking retards the degradation of anthocyanins in fruit peel and slows down the color change of fruit skin. | 0.05 g L−1 GA3 | Qu et al., 2021 [26] |
| ‘Valencia’ orange (Citrus sinensis (Linn.) Osbeck) | GA3 spraying retards the degradation of chlorophyll in fruit peel and reduces carotenoid content. | 500 μM GA3 | Keawmanee et al., 2022 [27] |
| ‘Chandler’ strawberry (Fragaria chiloensis (L.) Duchesne) | GA3 spraying increases fruit stem length and improves fruit shape index. | 75 ppm GA3 | Sharma et al., 2009 [19] |
| Tomato (Solanum lycopersicum L.) | GA3 spraying improves fruit shape index. | 0.1 mM GA3 | Chen et al., 2020 [9] |
| Horticultural Varieties | Main Points | Concentration of GAs | References |
|---|---|---|---|
| Tomato (Solanum lycopersicum L.) | GA3 spraying maintains the integrity of fruit cell membranes during low-temperature storage and activates cold resistance regulation. | 0.5 mM GA3 | Zhu et al., 2016 [42] |
| Cherry tomato (Solanum lycopersicum L.) | GA3 spraying maintains the integrity of fruit cell membranes during low-temperature storage and enhances antioxidant capacity. | 0.2 mmol L−1 GA3 | Ding et al., 2015 [44] |
| gib-3 tomato (Solanum lycopersicum L.) | GA3 deficiency leads to accelerated destruction of fruit cell membranes during low-temperature storage and reduces cold resistance. | GA mutant | Ding et al., 2016 [43] |
| Horticultural Varieties | Main Points | Concentration | References |
|---|---|---|---|
| Melon (Citrus limon (L.) Osbeck) | GA3 and AZX spraying inhibits the growth of Trichothecium roseum and Fusarium. | 100 mg L−1 GA3 + 100 mg L−1 AZX | Ma et al., 2005 [47] |
| ‘apple pear’ pear (Pyrus spp.) | GA3 spraying increases the thickness of protective tissue on the fruit surface and controls black skin disease. | 50 mg L−1 GA3 | Ma et al., 2018 [49] |
| Persimmon (Diospyros kaki Thunb.) | GA3 spraying reduces fruit cuticle cracking and controls black spot. | 200 μg mL–1 GA3 | Biton et al., 2014 [51] |
| gib-3 tomato (Solanum lycopersicum L.) | GA regulates early blight resistance by regulating hormone content. | GA mutant | Wu et al., 2018 [52] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Cao, Y.; Tang, J.; He, X.; Li, M.; Li, C.; Ren, X.; Ding, Y. Physiology and Application of Gibberellins in Postharvest Horticultural Crops. Horticulturae 2023, 9, 625. https://doi.org/10.3390/horticulturae9060625
Zhang J, Cao Y, Tang J, He X, Li M, Li C, Ren X, Ding Y. Physiology and Application of Gibberellins in Postharvest Horticultural Crops. Horticulturae. 2023; 9(6):625. https://doi.org/10.3390/horticulturae9060625
Chicago/Turabian StyleZhang, Jingyu, Yuhao Cao, Jia Tang, Xujie He, Ming Li, Chen Li, Xiaolin Ren, and Yuduan Ding. 2023. "Physiology and Application of Gibberellins in Postharvest Horticultural Crops" Horticulturae 9, no. 6: 625. https://doi.org/10.3390/horticulturae9060625
APA StyleZhang, J., Cao, Y., Tang, J., He, X., Li, M., Li, C., Ren, X., & Ding, Y. (2023). Physiology and Application of Gibberellins in Postharvest Horticultural Crops. Horticulturae, 9(6), 625. https://doi.org/10.3390/horticulturae9060625