



Bioefficacy of Lecanoric Acid Produced by Parmotrema austrosinense (Zahlbr.) Hale against Tea Fungal Pathogens


 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Antifungal Activity of Lecanoric Acid
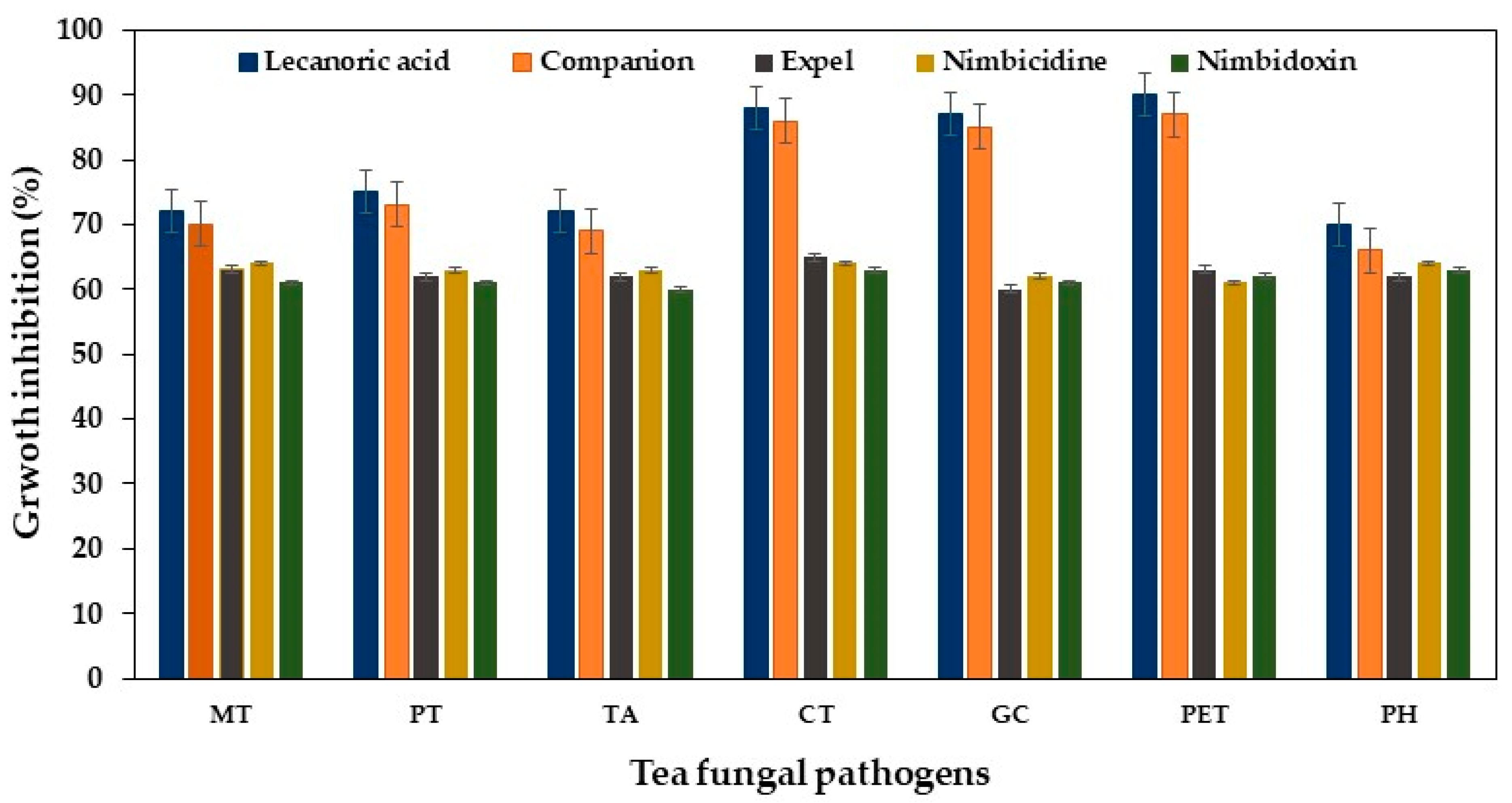
2.2. Suppressive Effect of Systemic and Botanical Fungicide
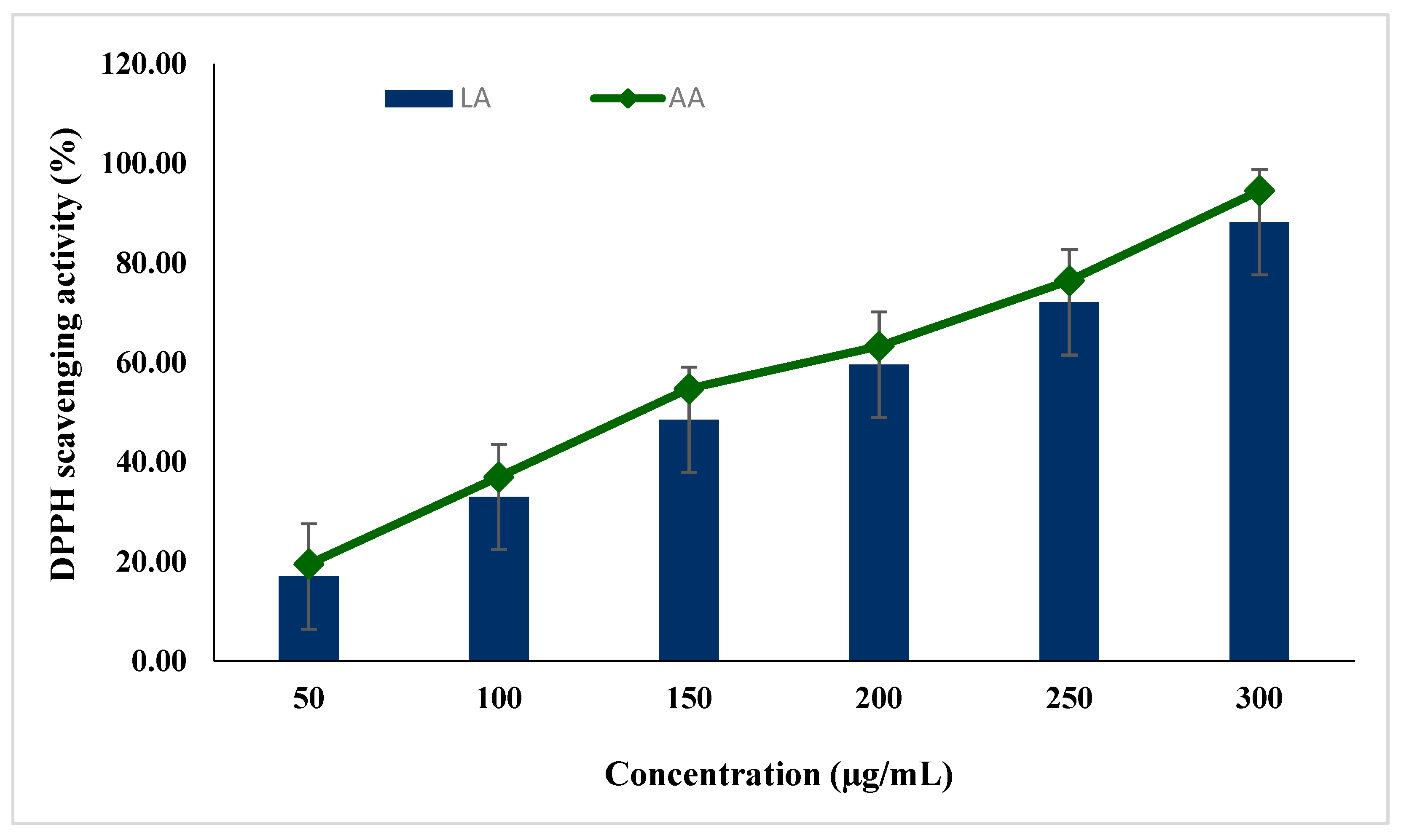
2.3. Free Radical Scavenging Activity of Lecanoric Acid
3. Discussion
4. Materials and Methods
4.1. Lichen Sample Collection and Identification
4.2. Isolation of Mycobionts from P. austrosinense
4.3. Production and Confirmation of Lecanoric Acid from Mycobionts
4.4. Purification of Lecanoric Acid by Column Chromatography
4.5. Antifungal Activity of Lecanoric Acid
4.6. Determination of Free Radical Scavenging Activity by DPPH Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ponmurugan, P.; Ayyappadasan, G.; Verma, R.S.; Nayaka, S. Survey, distribution pattern and elemental composition of lichens in Yercaud hills of Eastern Ghats in southern India. J. Environ. Biol. 2016, 37, 407. [Google Scholar]
- Shukla, V.; Joshi, G.P.; Rawat, M. Lichens as a potential natural source of bioactive compounds: A review. Phytochem. Rev. 2010, 9, 303–314. [Google Scholar] [CrossRef]
- Ganesan, A.; Thangapandian, M.; Ponnusamy, P.; Sundaraj, J. Antibacterial and antioxidant activity of Parmotrema reticulatum obtained from Eastern Ghats, Southern India. Biomed. Res. 2017, 28, 1593–1597. [Google Scholar]
- Vartia, K.O. Antibiotics in lichens. In The Lichens; Elsevier: Amsterdam, The Netherlands, 1973; pp. 547–561. [Google Scholar]
- Tanas, S.; Odabasoglu, F.; Halici, Z.; Cakir, A.; Aygun, H.; Aslan, A.; Suleyman, H. Evaluation of anti-inflammatory and antioxidant activities of Peltigera rufescens lichen species in acute and chronic inflammation models. J. Nat. Med. 2010, 64, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, G.; El-Naggar, A.M.; St. Clair, L.L.; O’Neill, K.L. Anticancer activities of selected species of North American lichen extracts. Phytother. Res. 2015, 29, 100–107. [Google Scholar] [CrossRef]
- Sati, S.; Joshi, S. Antibacterial activity of the Himalayan lichen Parmotrema nilgherrense extracts. Br. Microbiol. Res. J. 2011, 1, 26. [Google Scholar] [CrossRef]
- Roser, L.A.; Erkoc, P.; Ingelfinger, R.; Henke, M.; Ulshöfer, T.; Schneider, A.-K.; Laux, V.; Geisslinger, G.; Schmitt, I.; Fürst, R. Lecanoric acid mediates anti-proliferative effects by an M phase arrest in colon cancer cells. Biomed. Pharmacother. 2022, 148, 112734. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Matsuo, M.; Shibata, S. Biosynthesis of lichen depsides, lecanoric acid and atranorin. Chem. Pharm. Bull. 1965, 13, 1015–1017. [Google Scholar] [CrossRef] [Green Version]
- Paguirigan, J.A.; Liu, R.; Im, S.M.; Hur, J.-S.; Kim, W. Evaluation of Antimicrobial Properties of Lichen Substances against Plant Pathogens. Plant Pathol. J. 2022, 38, 25. [Google Scholar] [CrossRef]
- Nugraha, A.S.; Untari, L.F.; Laub, A.; Porzel, A.; Franke, K.; Wessjohann, L.A. Anthelmintic and antimicrobial activities of three new depsides and ten known depsides and phenols from Indonesian lichen: Parmelia cetrata Ach. Nat. Prod. Res. 2021, 35, 5001–5010. [Google Scholar] [CrossRef]
- Umezawa, H.; Shibamoto, N.; Naganawa, H.; Ayukawa, S.; Matsuzaki, M.; Takeuchi, T.; Kono, K.; Sakamoto, T. Isolation of lecanoric acid, an inhibitor of histidine decarboxylase from a fungus. J. Antibiot. 1974, 27, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponmurugan, P.; Manjukarunambika, K.; Gnanamangai, B.M. Impact of various foliar diseases on the biochemical, volatile and quality constituents of green and black teas. Australas. Plant Pathol. 2016, 45, 175–185. [Google Scholar] [CrossRef]
- Ponmurugan, P.; Gnanamangai, B.M.; Baby, U. Incidence, prevalence and clonal susceptibility of Phomopsis canker disease in southern Indian tea plantations. Indian Phytopathol. 2013, 66, 46–52. [Google Scholar]
- Ramya, M.; Ponmurugan, P.; Saravanan, D. Management of Cephaleuros parasiticaus Karst (Trentepohliales: Trentepohliaceae), an algal pathogen of tea plant, Camellia sinsensis (L.) (O. Kuntze). Crop Prot. 2013, 44, 66–74. [Google Scholar] [CrossRef]
- Baby, U.; Chandramouli, B. Biological antagonism of Trichoderma and Gliocladium spp. against certain primary root pathogens of tea. J. Plant. Crops 1996, 24, 249–255. [Google Scholar]
- Mareeswaran, J.; Nepolean, P.; Jayanthi, R.; Premkumar, S.; Radhakrishnan, B. In vitro studies on branch canker pathogen (Macrophoma sp.) infecting tea. J. Plant Pathol. Microbiol. 2015, 6, 284. [Google Scholar]
- König, G.M.; Wright, A.D. 1H and 13C-NMR and biological activity investigations of four lichen-derived compounds. Phytochem. Anal. 1999, 10, 279–284. [Google Scholar] [CrossRef]
- Kalidoss, R.; Ayyappadasan, G.; Gnanamangai, B.M.; Ponmurugan, P. Antibacterial activity of lichen Parmotrema spp. Inter. J Pharmaceut. Biol. Sci. 2020, 8, 7–11. [Google Scholar]
- Wei, A.H.; Zhou, D.N.; Ruan, J.L.; Cai, Y.L.; Xiong, C.M.; Li, M.X. Characterisation of phenols and antioxidant and hypolipidaemic activities of Lethariella cladonioides. J. Sci. Food Agric. 2012, 92, 373–379. [Google Scholar] [CrossRef]
- Nepolean, P.; Balamurugan, A.; Jayanthi, R.; Mareeswaran, J.; Premkumar, R. Bioefficacy of certain chemical and biofungicides against Hypoxylon spp. causing wood rot disease in tea. J. Plant Crops 2014, 42, 341–347. [Google Scholar]
- Kalim, S.; Luthra, Y.; Gandhi, S. Influence of Bavistin Seed Treatment on Morpho-physiological and Biochemical Parameters of Cowpea Roots Susceptible to Rhizoctonia Species. J. Mycol. Plant Pathol. 2000, 30, 375–379. [Google Scholar]
- Vyas, C.S. Hand Book of Systemic Fungicides; General Aspects; Mcgraw-Hill Education: New Delhi, India, 1993. [Google Scholar]
- Basile, A.; Conte, B.; Rigano, D.; Senatore, F.; Sorbo, S. Antibacterial and antifungal properties of acetonic extract of Feijoa sellowiana fruits and its effect on Helicobacter pylori growth. J. Med. Food 2010, 13, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Rai, H.; Upreti, D.; Trivedi, S.; Shukla, P. Antifungal activity of a common Himalayan foliose lichen Parmotrema tinctorum (Despr. ex Nyl.) Hale. Nat. Sci. 2011, 9, 167–171. [Google Scholar]
- Gulluce, M.; Aslan, A.; Sokmen, M.; Sahin, F.; Adiguzel, A.; Agar, G.; Sokmen, A. Screening the antioxidant and antimicrobial properties of the lichens Parmelia saxatilis, Platismatia glauca, Ramalina pollinaria, Ramalina polymorpha and Umbilicaria nylanderiana. Phytomedicine 2006, 13, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Moriano, C.; Gómez-Serranillos, M.P.; Crespo, A. Antioxidant potential of lichen species and their secondary metabolites. A systematic review. Pharm. Biol. 2016, 54, 1–17. [Google Scholar] [CrossRef]
- Wolseley, P. A Compendium of the Macrolichens from India, Nepal and Sri Lanka. By Dharani Dhar, Bishen Singh Mahendra Pal Singh, Dehra Dun, India. Lichenologist 2008, 40, 267–268. [Google Scholar] [CrossRef]
- Divakar, P.K.; Upreti, D.K. Parmelioid Lichens in India: A Revisionary Study; Bishen Singh Mahendra Pal Sin: Dehra Dun, India, 2005. [Google Scholar]
- Verma, N.; Behera, B.C. In Vitro culture of lichen partners: Need and Implications. Recent Adv. Lichenol. 2015, 2, 147–159. [Google Scholar]
- Orange, A.; James, P.W.; White, F. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2001. [Google Scholar]
- Yoshimura, I.; Kinoshita, Y.; Yamamoto, Y.; Huneck, S.; Yamada, Y. Analysis of secondary metabolites from lichen by high performance liquid chromatography with a photodiode array detector. Phytochem. Anal. 1994, 5, 197–205. [Google Scholar] [CrossRef]
- Adams, P.; Wong, J. The effect of chemical pesticides on the infection of sclerotia of Sclerotinia minor by the biocontrol agent Sporidesmium sclerotivorum. Phytopathology 1991, 81, 1340–1343. [Google Scholar] [CrossRef]
- Cappuccino, J.; Sherman, N. Microbiology, a Laboratory Manual; Benjamin, Cummings Pub. Co.: New York, NY, USA, 1992. [Google Scholar]
- Mareeswaran, J.; Nepolean, P. In vitro screening of chemical and organic fungicides against Branch canker disease in tea. J. Mycopathol. Res. 2016, 54, 299–301. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Tea Fungal Pathogens # | Concentration of Lecanoric Acid (μL)/Growth Inhibition (%) | |||
|---|---|---|---|---|
| 5 | 10 | 15 | 20 | |
| Hypoxylon serpens | 53.5 ± 1.3 | 75.3 ± 1.3 | 77.7 ± 1.3 | 75.0 ± 1.5 |
| Macrophoma theicola | 53.3 ± 1.5 | 73.0 ± 1.7 | 76.5 ± 1.5 | 70.7 ± 1.8 |
| Phomopsistheae | 60.0 ± 1.7 | 76.0 ± 2.0 | 80.5 ± 1.3 | 73.0 ± 2.4 |
| Tunstallia aculeate | 52.5 ± 1.6 | 72.0 ± 1.5 | 75.0 ± 1.4 | 69.5 ± 2.0 |
| Cercosporatheae | 71.3 ± 2.5 | 88.0 ± 1.1 | 91.5 ± 2.0 | 85.5 ± 2.5 |
| Glomerella cingulata | 65.5 ± 2.1 | 84.0 ± 2.3 | 86.5 ± 2.1 | 80.7 ± 2.3 |
| Pestalotiopsis theae | 68.7 ± 2.0 | 90.7 ± 1.5 | 90.0 ± 2.3 | 83.3 ± 1.8 |
| Poria hypolateritia | 50.3 ± 1.5 | 71.5 ± 1.3 | 73.3 ± 1.5 | 68.5 ± 1.6 |
| * CD at p = 0.05 | 5.4 | 6.5 | 5.3 | 4.6 |
| Tea Fungal Pathogens | Solvents (mg mL)/Growth Inhibition | ||||
|---|---|---|---|---|---|
| Ethanol | Methanol | Acetone | Petroleum Ether | Water | |
| Hypoxylon serpens | 72.5 ± 1.2 | 74.0 ± 1.7 | 72.0 ± 1.5 | 73.0 ± 1.6 | 52.5 ± 1.3 |
| Macrophoma theicola | 73.0 ± 1.3 | 74.7 ± 1.0 | 74.3 ± 1.5 | 73.3 ± 1.8 | 55.7 ± 1.3 |
| Phomopsis theae | 74.3 ± 0.9 | 76.0 ± 1.5 | 75.3 ± 1.3 | 75.0 ± 1.5 | 55.3 ± 1.1 |
| Tunstallia aculeate | 71.3 ± 1.1 | 73.0 ± 1.3 | 72.3 ± 1.0 | 72.5 ± 1.2 | 52.0 ± 1.3 |
| Cercospora theae | 84.0 ± 2.0 | 87.5 ± 2.0 | 85.0 ± 1.7 | 85.0 ± 1.9 | 64.0 ± 2.1 |
| Glomerella cingulata | 80.7 ± 1.5 | 85.3 ± 2.0 | 82.3 ± 2.5 | 81.5 ± 1.5 | 53.0 ± 2.1 |
| Pestalotiopsis theae | 81.5 ± 1.3 | 85.7 ± 1.8 | 82.0 ± 2.0 | 83.5 ± 1.5 | 54.5 ± 2.0 |
| Poria hypolateritia | 70.7 ± 1.1 | 73.0 ± 1.6 | 71.5 ± 1.6 | 70.7 ± 1.4 | 53.3 ± 1.7 |
| * CD at p = 0.05 | 4.5 | 5.3 | 4.0 | 3.8 | 3.3 |
| Tea Fungal Pathogens | Concentration of Companion Fungicide (μL)/Growth Inhbition (%) | |||
|---|---|---|---|---|
| 10 | 20 | 30 | 40 | |
| Hypoxylon serpens | 63.0 ± 1.5 | 71.0 ± 1.7 | 70.3 ± 1.1 | 68.3 ± 1.3 |
| Macrophoma theicola | 65.3 ± 1.6 | 70.3 ± 1.5 | 70.7 ± 1.3 | 67.0 ± 1.4 |
| Phomopsis theae | 66.0 ± 1.5 | 71.5 ± 1.7 | 70.3 ± 0.8 | 70.3 ± 1.5 |
| Tunstallia aculeate | 63.0 ± 1.3 | 69.3 ± 1.3 | 69.5 ± 0.9 | 67.5 ± 1.7 |
| Cercospora theae | 72.3 ± 1.6 | 85.7 ± 1.5 | 81.5 ± 0.8 | 80.7 ± 1.1 |
| Glomerella cingulata | 75.5 ± 1.5 | 80.7 ± 1.7 | 80.3 ± 1.6 | 78.0 ± 1.6 |
| Pestalotiopsis theae | 75.3 ± 1.6 | 83.5 ± 1.8 | 83.0 ± 1.9 | 80.0 ± 1.5 |
| Poria hypolateritia | 63.7 ± 0.9 | 68.5 ± 1.5 | 65.7 ± 1.7 | 65.0 ± 1.1 |
| * CD at p = 0.05 | 4.7 | 4.2 | 3.3 | 5.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajendran, K.; Ponmurugan, P.; Gnanamangai, B.M.; Karuppiah, P.; Shaik, M.R.; Khan, M.; Khan, M.; Shaik, B. Bioefficacy of Lecanoric Acid Produced by Parmotrema austrosinense (Zahlbr.) Hale against Tea Fungal Pathogens. Horticulturae 2023, 9, 705. https://doi.org/10.3390/horticulturae9060705
Rajendran K, Ponmurugan P, Gnanamangai BM, Karuppiah P, Shaik MR, Khan M, Khan M, Shaik B. Bioefficacy of Lecanoric Acid Produced by Parmotrema austrosinense (Zahlbr.) Hale against Tea Fungal Pathogens. Horticulturae. 2023; 9(6):705. https://doi.org/10.3390/horticulturae9060705
Chicago/Turabian StyleRajendran, Kalidoss, Ponnusamy Ponmurugan, Balasubramanian Mythili Gnanamangai, Ponmurugan Karuppiah, Mohammed Rafi Shaik, Mujeeb Khan, Merajuddin Khan, and Baji Shaik. 2023. "Bioefficacy of Lecanoric Acid Produced by Parmotrema austrosinense (Zahlbr.) Hale against Tea Fungal Pathogens" Horticulturae 9, no. 6: 705. https://doi.org/10.3390/horticulturae9060705
APA StyleRajendran, K., Ponmurugan, P., Gnanamangai, B. M., Karuppiah, P., Shaik, M. R., Khan, M., Khan, M., & Shaik, B. (2023). Bioefficacy of Lecanoric Acid Produced by Parmotrema austrosinense (Zahlbr.) Hale against Tea Fungal Pathogens. Horticulturae, 9(6), 705. https://doi.org/10.3390/horticulturae9060705