
Unraveling the Pomegranate Genome: Comprehensive Analysis of R2R3-MYB Transcription Factors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Identification of R2R3-MYB Gene Family Members and Physicochemical Properties Analysis in Pomegranate
2.3. Phylogenetic Analysis
2.4. Chromosomal Position and Collinearily Analysis
2.5. Gene Structure and Protein Motif Analysis
2.6. Analysis of Promoter Cis-Acting Elements
2.7. RNA Extraction Library Construction, and Sequencing
2.8. Expression of R2R3-MYB Genes in Pomegranate
2.9. Real-Time Quantitative PCR (qRT-PCR)
3. Results
3.1. Identification of R2R3-MYB Gene Family Members Physicochemical Properties of Pomegranate R2R3-MYB Proteins
3.2. Phylogenetic Analysis of the R2R3-MYB Gene Family
3.3. Chromosomal Position and Collinearity Analysis
3.4. R2R3-MYB Gene Structures and Protein Domains Analysis in Pomegranate
3.5. Promoter Analysis of R2R3-MYB Genes in Pomegranate
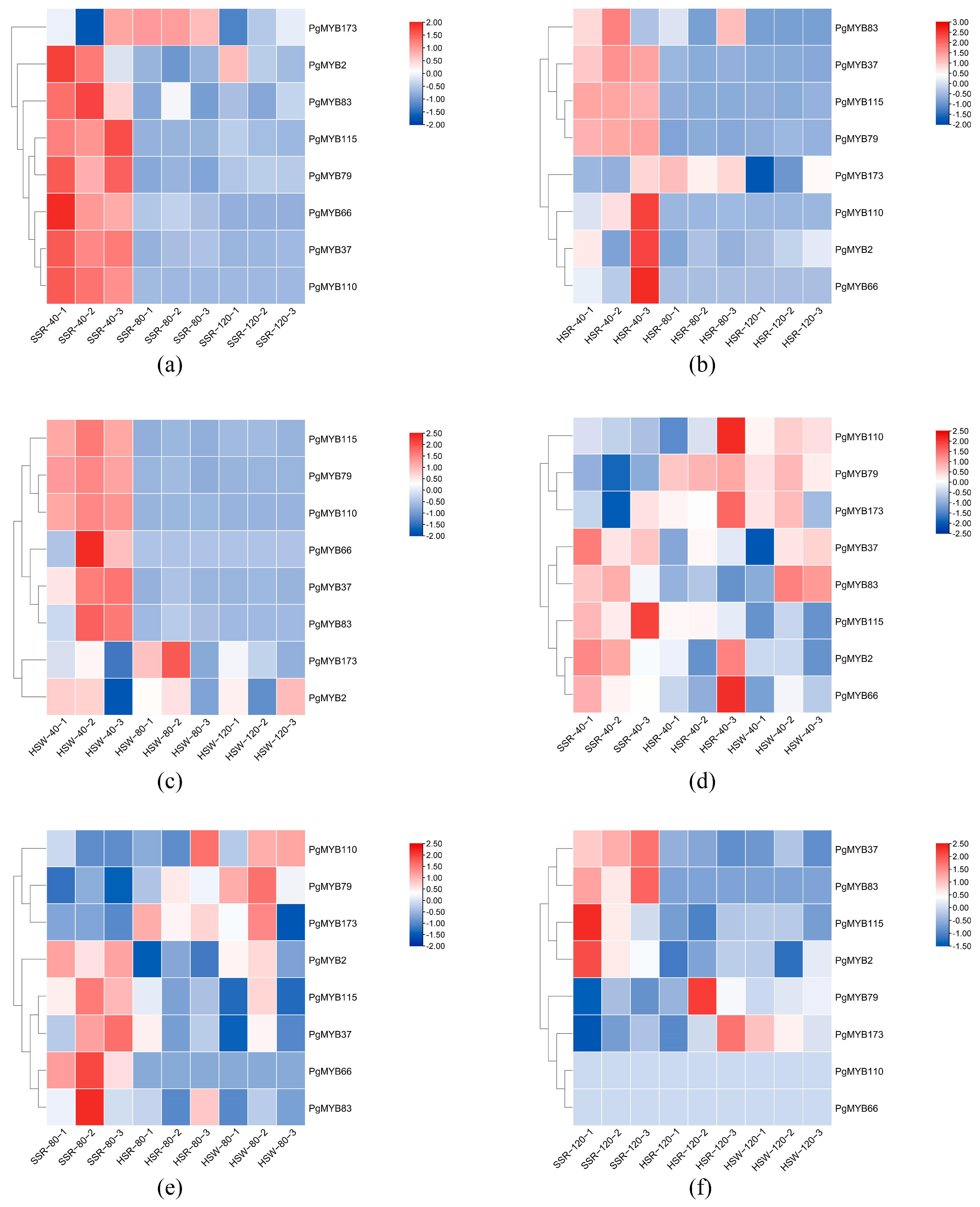
3.6. Expression of R2R3-MYB Genes
3.7. The Validation of PgMYB Expression with qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yuan, Z.H.; Fang, Y.M.; Zhang, T.K.; Fei, Z.J.; Han, F.M.; Liu, C.Y.; Liu, M.; Xiao, W.; Zhang, W.J.; Wu, S.; et al. The pomegranate (Punica granatum L.) genome provides insights into fruit quality and ovule developmental biology. Plant Biotechnol. J. 2018, 16, 1363–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, J.A.T.; Rana, T.S.; Narzary, D.; Verma, N.; Meshram, D.T.; Ranade, S.A. Pomegranate biology and biotechnology: A review. Sci. Hortic. 2013, 160, 85–107. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, X.; Ni, H.H.; Yuan, F.Y.; Zhang, S.M. Identification and Functional Analysis of CAD Gene Family in Pomegranate (Punica granatum). Genes 2023, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.H.; Ge, D.P.; Ren, Y.; Dong, J.M.; Zhao, X.Q.; Liu, X.Q.; Yuan, Z.H. The comparative analysis and identification of secondary metabolites between Tibet wild and cultivated pomegranates (Punica granatum L.) in China. J. Integr. Agric. 2022, 21, 736–750. [Google Scholar] [CrossRef]
- Murthy, K.N.; Reddy, V.K.; Veigas, J.M.; Murthy, U.D. Study on wound healing activity of Punica granatum peel. J. Med. Food 2004, 7, 256–259. [Google Scholar] [CrossRef]
- Alighourchi, H.; Barzegar, M.; Abbasi, S. Anthocyanins characterization of 15 Iranian pomegranate (Punica granatum L.) varieties and their variation after cold storage and pasteurization. Eur. Food Res. Technol. 2008, 227, 881–887. [Google Scholar] [CrossRef]
- Zhang, X.H.; Zhao, Y.J.; Ren, Y.; Wang, Y.Y.; Yuan, Z.H. Fruit Breeding in Regard to Color and Seed Hardness: A Genomic View from Pomegranate. Agronomy 2020, 10, 991. [Google Scholar] [CrossRef]
- Xue, H.; Cao, S.Y.; Li, H.X.; Zhang, J.; Niu, J.; Chen, L.N.; Zhang, F.H.; Zhao, D.G. De novo transcriptome assembly and quantification reveal differentially expressed genes between soft-seed and hard-seed pomegranate (Punica granatum L.). PLoS ONE 2017, 12, e0178809. [Google Scholar] [CrossRef] [Green Version]
- Dalimov, D.N.; Dalimova, G.N.; Bhatt, M. Chemical composition and lignins of tomato and pomegranate seeds. Chem. Nat. Compd. 2003, 39, 37–40. [Google Scholar] [CrossRef]
- Niu, J.; Cao, D.; Li, H.; Xue, H.; Chen, L.; Liu, B.; Cao, S. Quantitative proteomics of pomegranate varieties with contrasting seed hardness during seed development stages. Tree Genet. Genomes 2018, 14, 14. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Niu, J.; Cao, D.; Li, H.; Xue, H.; Chen, L.; Zhang, F.L.; Zhao, D. Comparative proteomics analysis of pomegranate seeds on fruit maturation period (Punica granatum L.). J. Integr. Agric. 2015, 14, 2558–2564. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Xu, C.; Ming, R.; Tang, H.; Guyot, R.; Kramer, E.M.; Hu, Y.; Yi, X.; Qi, Y.; Xu, X.; et al. The pomegranate (Punica granatum L.) genome and the genomics of punicalagin biosynthesis. Plant J. 2017, 91, 1108–1128. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Yang, S.S.; Liang, Z.; Feng, B.R.; Liu, L.; Huang, Y.B.; Tang, Y.X. Genome-wide analysis of the MYB transcription factor superfamily in soybean. BMC Plant Biol. 2012, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Millard, P.S.; Kragelund, B.B.; Burow, M. R2R3 MYB transcription factors-functions outside the DNA-binding domain. Trends Plant Sci. 2009, 24, 934–946. [Google Scholar] [CrossRef]
- Jiang, C.H.; Gu, J.Y.; Chopra, S.; Gu, X.; Peterson, T. Ordered origin of the typical two- and three-repeat Myb genes. Gene 2004, 326, 13–22. [Google Scholar] [CrossRef]
- Xu, B.J.; Chen, B.; Qi, X.L.; Liu, S.L.; Zhao, Y.B.; Tang, C.; Meng, X.L. Genome-wide Identification and Expression Analysis of RcMYB Genes in Rhodiola crenulate. Front. Genet. 2022, 13, 831611. [Google Scholar] [CrossRef]
- Jacopo, M.; Pierdomenico, P.; Silvia, G. Novel R2R3 MYB transcription factors regulate anthocyanin synthesis in Aubergine tomato plants. BMC Plant Biol. 2023, 23, 148. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Matus, J.T.; Aquea, F.; Arce-Johnson, P. Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes. BMC Plant Biol. 2008, 8, 83. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ding, Z.; Ruan, M.; Yu, X.; Peng, M.; Liu, Y. Kiwifruit R2R3-MYB transcription factors and contribution of the novel AcMYB75 to red kiwifruit anthocyanin biosynthesis. Sci. Rep. 2017, 7, 16861. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; He, J.; Dong, J.; Hou, X.; Zhang, X. Genomic survey and expression profiling of the MYB gene family in watermelon. Hortic. Plant J. 2018, 4, 1–15. [Google Scholar] [CrossRef]
- Xie, F.F.; Hua, Q.Z.; Chen, C.B.; Zhang, Z.K.; Zhang, R.; Zhao, J.T.; Hu, G.B.; Chen, J.Y.; Qin, Y.H. Genome-Wide Characterization of R2R3-MYB Transcription Factors in Pitaya Reveals a R2R3-MYB Repressor HuMYB1 Involved in Fruit Ripening through Regulation of Betalain Biosynthesis by Repressing Betalain Biosynthesis-Related Genes. Cells 2021, 10, 1949. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.H.; Zhang, S.Z.; Wang, R.K.; Zhang, R.F.; Hao, Y.J. Genome wide analysis of the apple MYB transcription factor family allows the identification of MdoMYB121 gene confering abiotic stress tolerance in plants. PLoS ONE 2013, 8, e69955. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.H.; Hu, Q.; Shen, J.L.; Liu, S.M.; Yang, Z.G.; Chen, K.; Klosterman, S.J.; Javornik, B.; Zhang, X.L.; Zhu, L.F. GhMYB4 downregulates lignin biosynthesis and enhances cotton resistance to Verticillium dahliae. Plant Cell Rep. 2021, 40, 735–751. [Google Scholar] [CrossRef]
- Zhou, J.L.; Lee, C.H.; Zhong, R.Q.; Ye, Z.H. MYB58 and MYB63 Are Transcriptional Activators of the Lignin Biosynthetic Pathway during Secondary Cell Wall Formation in Arabidopsis. Plant Cell 2009, 21, 248–266. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, R.L.; Zhong, R.; Ye, Z.H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009, 50, 1950–1964. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.H.; Jeon, H.W.; Kim, W.C.; Kim, J.Y.; Han, K.H. The MYB46/MYB83-mediated transcriptional regulatory programme is a gatekeeper of secondary wall biosynthesis. Ann. Bot. 2014, 114, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Cheng, S.F.; Liu, X.Y.; Du, H.; Dai, M.Q.; Yang, W.J.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef]
- Li, Z.W.; Zhou, Y.J.; Liang, H.Z.; Li, Q.; Jiang, Y.M.; Duan, X.W.; Jiang, G.X. MaMYB13 is involved in response to chilling stress via activating expression of VLCFAs and phenylpropanoids biosynthesis-related genes in postharvest banana fruit. Food Chem. 2022, 405, 134957. [Google Scholar] [CrossRef]
- Gomez-Casati, F.D.; Busi, V.M.; Barchiesi, J.; Peralta, A.D.; Hedin, N.; Bhadauria, V. Application of bioinformatics to plant breeding. Curr. Issues Mol. Biol. 2018, 27, 89–103. [Google Scholar] [CrossRef]
- Gong, L.Y. Cloning and Expression Analysis of Genes Related with Lignin Synthesis in Pomegranate Seed. Ph.D. Thesis, Anhui Agricultural University, Hefei, China, 2014. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Zhu, L.; Guan, Y.X.; Zhang, Z.H.; Song, A.P.; Chen, S.M.; Jiang, J.F.; Chen, F.D. CmMYB8 encodes an R2R3 MYB transcription factor which represses lignin and flavonoid synthesis in chrysanthemum. Plant Physiol. Biochem. 2020, 149, 217–224. [Google Scholar] [CrossRef]
- Soler, M.; Plasencia, A.; Larbat, R.; Pouzet, C.; Jauneau, A.; Rivas, S.; Pesquet, E.; Lapierre, C.; Truchet, I.; Grima-Pettenati, J. The Eucalyptus linker histone variant EgH1.3 cooperates with the transcription factor EgMYB1 to control lignin biosynthesis during wood formation. New Phytol. 2017, 213, 287–299. [Google Scholar] [CrossRef]
- Tombuloglu, H. Genome-wide identification and expression analysis of R2R3, 3R-and 4R-MYB transcription factors during lignin biosynthesis in flax (Linum usitatissimum). Genomics 2020, 112, 782–795. [Google Scholar] [CrossRef]
- Tian, Q.Y. Functional Characterization of PtoMYB216 and PtoMYB125 Involved in Transcriptional Regulation of Lignin Biosysnthesis in Populus. Ph.D. Thesis, Southwest University, Chongqing, China, 2013. [Google Scholar]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Zoreky, N.S. Antimicrobial activity of pomegranate (Punica granatum L.) fruit peels. Int. J. Food Microbiol. 2009, 134, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Pantiora, P.D.; Balaouras, A.I.; Mina, I.K.; Freris, C.I.; Pappas, A.C.; Danezis, G.P.; Zoidis, E.; Georgiou, C.A. The Therapeutic Alliance between Pomegranate and Health Emphasizing on Anticancer Properties. Antioxidants 2023, 12, 187. [Google Scholar] [CrossRef] [PubMed]
- Caruso, A.; Barbarossa, A.; Tassone, A.; Ceramella, J.; Carocci, A.; Catalano, A.; Basile, G.; Fasio, A.; Iacopetta, D.; Franchini, C.; et al. Pomegranate: Nutraceutical with promising benefits on human health. Appl. Sci. 2020, 10, 6915. [Google Scholar] [CrossRef]
- Escarcega, G.H.; Sánchez-Chávez, E.; Álvarez, S.P.; Caballero, S.M.; Parra, J.M.S.; Flores-Córdova, M.A.; Salazar, N.A.S.; Barrios, D.L.O. Determination of antioxidant phenolic, nutritional quality and volatiles in pomegranates (Punica granatum L.) cultivated in Mexico. Int. J. Food Prop. 2020, 23, 979–991. [Google Scholar] [CrossRef]
- Karlen, S.D.; Zhang, C.C.; Peck, M.L.; Smith, R.A.; Padmakshan, D.; Helmich, K.E.; Free, H.C.A.; Lee, S.; Smith, B.G.; Lu, F.C. Monolignol ferulate conjugates are naturally incorporated into plant lignins. Sci. Adv. 2016, 2, e1600393. [Google Scholar] [CrossRef] [Green Version]
- Xiao, R.X.; Zhang, C.; Guo, X.R.; Li, H.; Lu, H. MYB Transcription Factors and Its Regulation in Secondary Cell Wall Formation and Lignin Biosynthesis during Xylem Development. Int. J. Mol. Sci. 2021, 22, 3560. [Google Scholar] [CrossRef]
- Goicoechea, M.; Lacombe, E.; Legay, S.; Mihaljevic, S.; Rech, P.; Jauneau, A.; Lapierre, C.; Pollet, B.; Verhaegen, D.; Chaubet-Gigot, N.; et al. EgMYB2, a new transcriptional activator from Eucalyptus xylem, regulates secondary cell wall formation and lignin biosynthesis. Plant J. 2005, 43, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, R.; Dalton, S.; Roberts, L.A.; Moron-Garcia, O.M.; Iacono, R.; Kosik, O.; Gallagher, J.A.; Bosch, M. Modified expression of ZmMYB167 in Brachypodium distachyon and Zea mays leads to increased cell wall lignin and phenolic content. Sci. Rep. 2019, 9, 8800. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.Y.; Wang, X.Q.; Li, C.F.; Lu, W.X.; Yang, L.; Jiang, Y.Z.; Luo, K.M. Functional Characterization of the Poplar R2R3-MYB Transcription Factor PtoMYB216 Involved in the Regulation of Lignin Biosynthesis during Wood Formation. PLoS ONE 2013, 8, e76369. [Google Scholar] [CrossRef]
- Hou, X.J.; Li, S.B.; Liu, S.R.; Hu, C.G.; Zhang, J.Z. Genome-Wide Classification and Evolutionary and Expression Analyses of Citrus MYB Transcription Factor Families in Sweet Orange. PLoS ONE 2014, 9, e112375. [Google Scholar] [CrossRef]
- Ma, D.W.; Constabel, C.P. MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants. Trends Plant Sci. 2019, 24, 275–289. [Google Scholar] [CrossRef]
- Liu, J.Y.; Osbourn, A.; Ma, P.D. MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Zhang, B.H.; Gu, G.; Yuan, J.Z.; Shen, S.J.; Jin, L.; Lin, Z.Q.; Lin, J.F.; Xie, X.F. Genome-wide identification and expression analysis of the R2R3-MYB gene family in tobacco (Nicotiana tabacum L.). BMC Genom. 2022, 23, 432. [Google Scholar] [CrossRef]
- Sowjanya, P.R.; Shilpa, P.; Patil, G.P.; Babu, D.K.; Sharma, J.; Sangnure, V.R.; Mundewadikar, D.M.; Natarajan, P.; Marathe, A.R.; Reddy, U.K. Reference quality genome sequence of Indian pomegranate cv. ‘Bhagawa’ (Punica granatum L.). Front. Plant Sci. 2022, 13, 947164. [Google Scholar] [CrossRef]
- Bai, J.S.; Song, M.J.; Gao, J.; Li, G.T. Whole genome duplication and dispersed duplication characterize the evolution of the plant PINOID gene family across plant species. Gene 2022, 829, 146494. [Google Scholar] [CrossRef]
- Kroon, M.; Lameijer, E.W.; Lakenberg, N.; Hehir-Kwa, J.Y.; Thung, D.T.; Slagboom, P.E.; Kok, J.N.; Ye, K. Detecting dispersed duplications in high-throughput sequencing data using a database-free approach. Bioinformatics 2016, 32, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Xue, C.; Li, J.M.; Qiao, X.; Li, L.T.; Yu, L.A.; Huang, Y.H.; Wu, J. Genome-Wide Identification, Evolution and Functional Divergence of MYB Transcription Factors in Chinese White Pear (Pyrus bretschneideri). Plant Cell Physiol. 2016, 57, 824–847. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suo, H.; Zhang, X.; Hu, L.; Ni, H.; Langjia, R.; Yuan, F.; Zhang, M.; Zhang, S. Unraveling the Pomegranate Genome: Comprehensive Analysis of R2R3-MYB Transcription Factors. Horticulturae 2023, 9, 779. https://doi.org/10.3390/horticulturae9070779
Suo H, Zhang X, Hu L, Ni H, Langjia R, Yuan F, Zhang M, Zhang S. Unraveling the Pomegranate Genome: Comprehensive Analysis of R2R3-MYB Transcription Factors. Horticulturae. 2023; 9(7):779. https://doi.org/10.3390/horticulturae9070779
Chicago/Turabian StyleSuo, Heming, Xuan Zhang, Lei Hu, Huihui Ni, Renzeng Langjia, Fangyu Yuan, Maowen Zhang, and Shuiming Zhang. 2023. "Unraveling the Pomegranate Genome: Comprehensive Analysis of R2R3-MYB Transcription Factors" Horticulturae 9, no. 7: 779. https://doi.org/10.3390/horticulturae9070779
APA StyleSuo, H., Zhang, X., Hu, L., Ni, H., Langjia, R., Yuan, F., Zhang, M., & Zhang, S. (2023). Unraveling the Pomegranate Genome: Comprehensive Analysis of R2R3-MYB Transcription Factors. Horticulturae, 9(7), 779. https://doi.org/10.3390/horticulturae9070779