Neuromechanical Model of Rat Hindlimb Walking with Two-Layer CPGs †

 , ,
, ,
Abstract
:1. Introduction
2. Methods
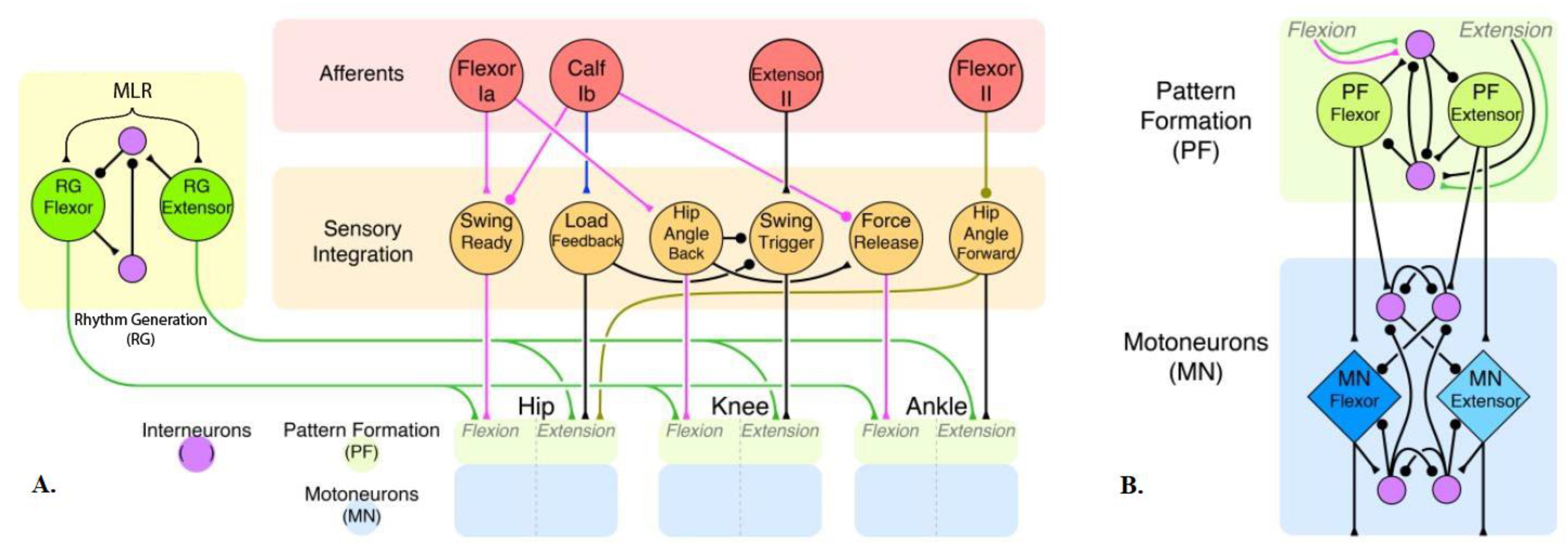
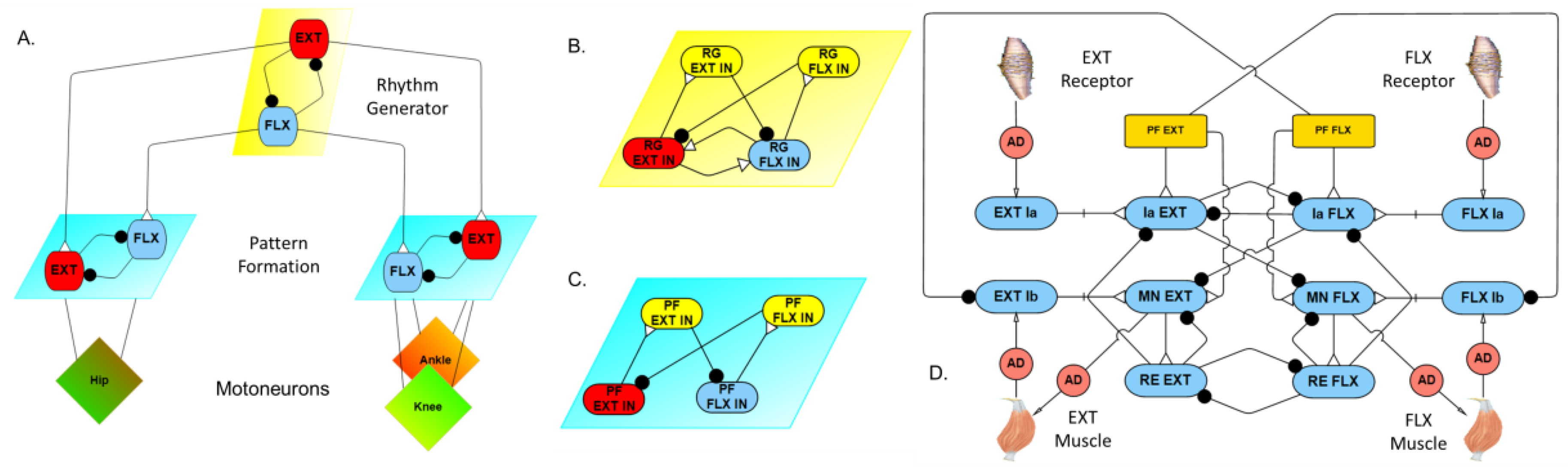
2.1. Model Description
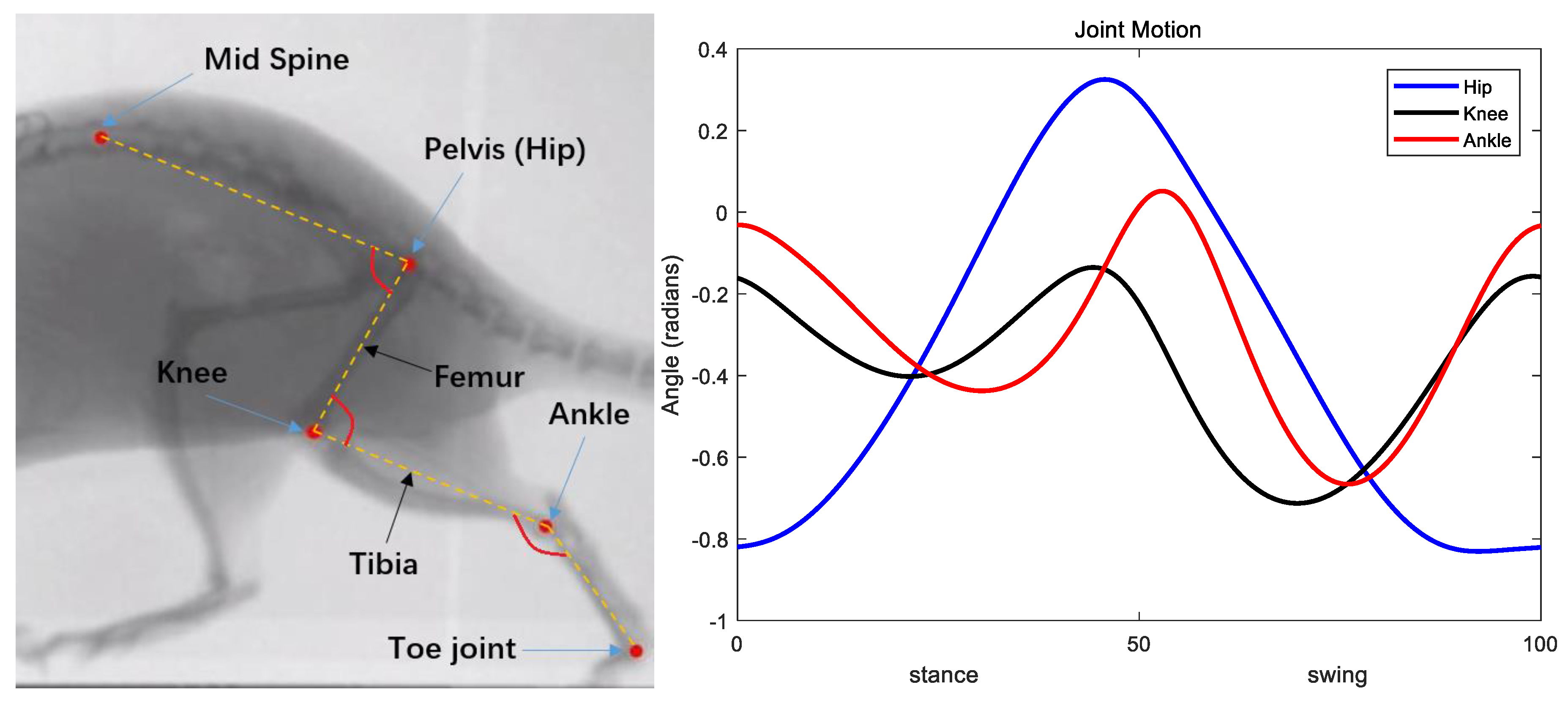
2.2. Model Validation
2.3. Animal Experimental Procedure
3. Results
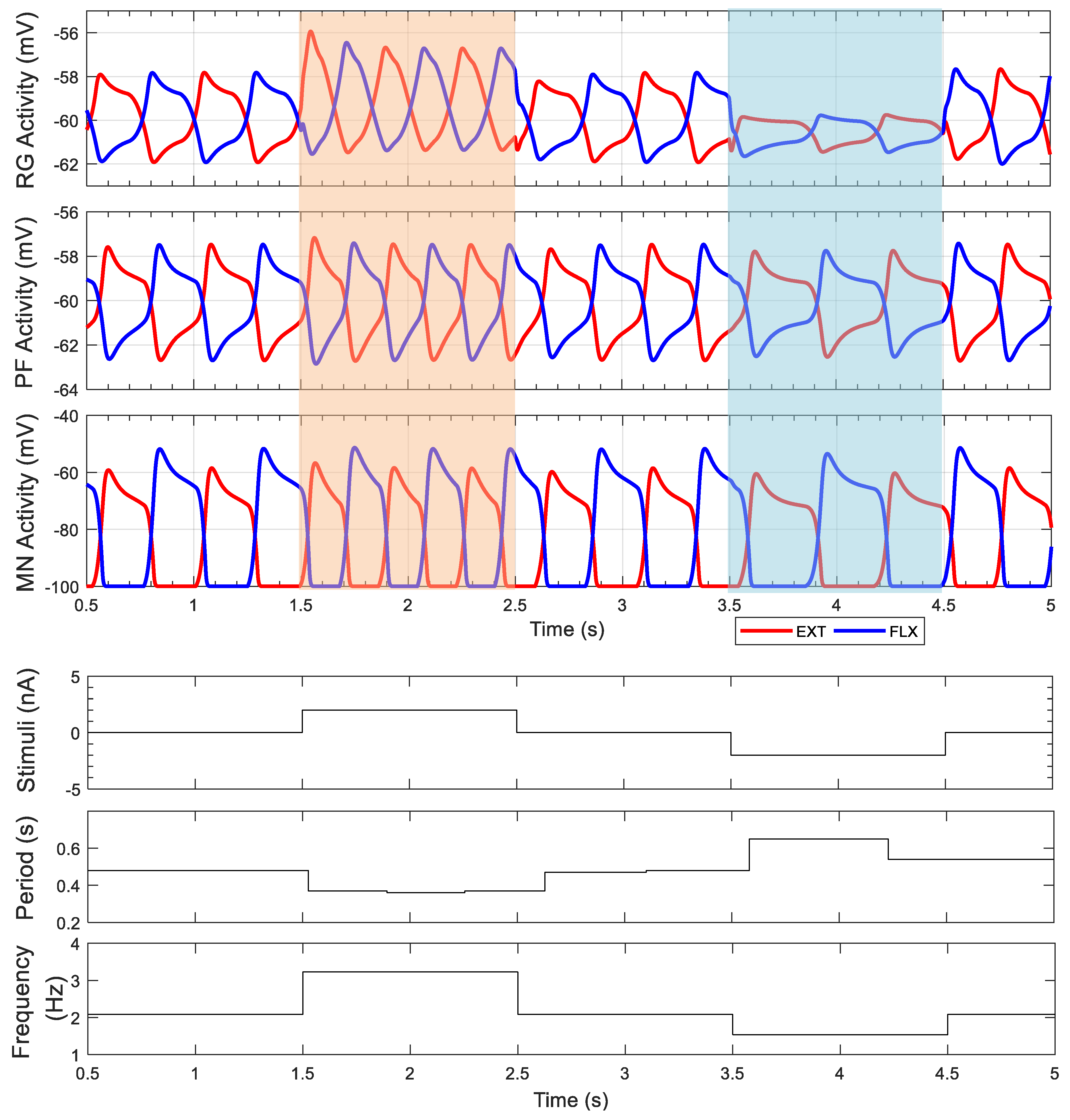
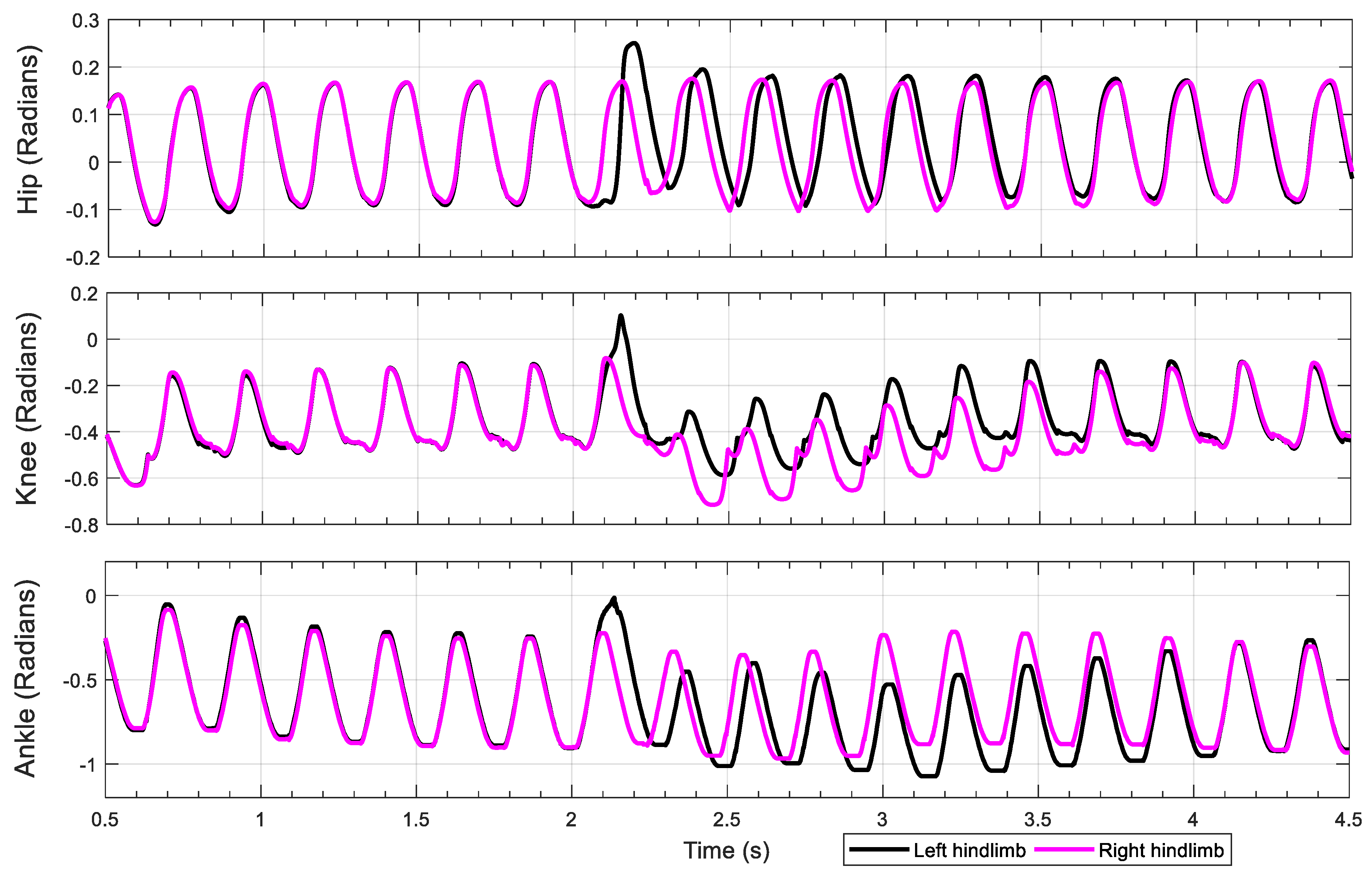
3.1. Intact Model
3.2. Knee–Ankle Synergy
4. Discussion
4.1. Neural Control Adaptations
4.2. Knee–Ankle Synergy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neuron | (mV) | (mV) | (ms) | (mV) | (ms) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RG | 5 | 1 | −60 | 1.5 | 0.5 | −0.6 | −60 | 0.35 | 1 | 0.2 | −40 | 2 |
| PF | 5 | 1 | −60 | 1.5 | 0.5 | −0.6 | −60 | 0.35 | 1 | 0.2 | −40 | 2 |
| MN | 5 | 1 | −100 | 0 | - | - | - | - | - | - | - | - |
| IN | 5 | 1 | -60 | 0 | - | - | - | - | - | - | - | - |
| Synapse | (μS) | (μS) | (μS) | (mV) | (mV) | (mV) |
|---|---|---|---|---|---|---|
| RG to IN | 2.749 | - | - | −40 | −60 | −25 |
| IN to RG | 2.749 | - | - | −70 | −60 | −25 |
| Between RG | 0.1 | - | - | −40 | −65 | −40 |
| PF to IN | 2.749 | - | - | −40 | −60 | −25 |
| IN to PF | 2.749 | - | - | −70 | −60 | −25 |
| RG to PF | 0.1 | - | - | −40 | −60 | −40 |
| PF to MN | - | - | - | −10 | −60 | −50 |
| Hip | - | 2.565 | 3.632 | - | −60 | −40 |
| Knee | - | 4.93 | 1.516 | - | −60 | −40 |
| Ankle | - | 4.054 | 4.522 | - | −60 | −40 |
| PF to Ia | 0.5 | - | - | −40 | −60 | −55 |
| Between Ia | 0.5 | - | - | −70 | −60 | −40 |
| Ia to MN | 2 | - | - | −100 | −60 | −40 |
| MN to RE | 0.5 | - | - | −40 | −100 | −10 |
| Between R | 0.5 | - | - | −70 | −60 | −40 |
| R to MN | 0.5 | - | - | −100 | −60 | −40 |
| R to Ia | 0.5 | - | - | −70 | −60 | −40 |
Appendix B

References
- Brown, T.G. On the nature of the fundamental activity of the nervous centres; together with an analysis of the conditioning of rhythmic activity in progression, and a theory of the evolution of function in the nervous system. J. Physiol. 1914, 48, 18–46. [Google Scholar] [CrossRef] [PubMed]
- Hägglund, M.; Dougherty, K.J.; Borgius, L.; Itohara, S.; Iwasato, T.; Kiehn, O. Optogenetic dissection reveals multiple rhythmogenic modules underlying locomotion. Proc. Natl. Acad. Sci. USA 2013, 110, 11589–11594. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S.; McClellan, A.; Sigvardt, K.; Wallén, P.; Wilén, M. Activation of NMDA-receptors elicits “fictive locomotion” in lamprey spinal cord in vitro. Acta Physiol. Scand. 1981, 113, 549–551. [Google Scholar] [CrossRef] [PubMed]
- Tryba, A.K.; Peña, F.; Ramirez, J.-M. Gasping activity in vitro: A rhythm dependent on 5-HT2A receptors. J. Neurosci. 2006, 26, 2623–2634. [Google Scholar] [CrossRef] [PubMed]
- Wenger, N.; Moraud, E.M.; Gandar, J.; Musienko, P.; Capogrosso, M.; Baud, L.; Le Goff, C.G.; Barraud, Q.; Pavlova, N.; Dominici, N.; et al. Spatiotemporal neuromodulation therapies engaging muscle synergies improve motor control after spinal cord injury. Nat. Med. 2016, 22, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, T.; Ewald, A.; Von Twickel, A.; Büschges, A. Controlling legs for locomotion—Insights from robotics and neurobiology. Bioinspir. Biomim. 2015, 10, 41001. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K.; Ekeberg, Ö.; Büschges, A. Assessing sensory function in locomotor systems using neuro-mechanical simulations. Trends Neurosci. 2006, 29, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.J.; Szczecinski, N.S.; Andrada, E.; Fischer, M.; Quinn, R.D. Using animal data and neural dynamics to reverse engineer a neuromechanical rat model. Lect. Notes Comput. Sci. 2015, 9222, 211–222. [Google Scholar] [CrossRef]
- Hunt, A.; Szczecinski, N.; Quinn, R. Development and training of a neural controller for hind leg walking in a dog robot. Front. Neurorobot. 2017, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- McCrea, D.A.; Rybak, I.A. Organization of mammalian locomotor rhythm and pattern generation. Brain Res. Rev. 2008, 57, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S.; Zangger, P. On the central generation of locomotion in the low spinal cat. Exp. Brain Res. 1979, 34, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Lafreniere-Roula, M.; McCrea, D.A. Deletions of rhythmic motoneuron activity during fictive locomotion and scratch provide clues to the organization of the mammalian central pattern generator. J. Neurophysiol. 2005, 94, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Rybak, I.A.; Stecina, K.; Shevtsova, N.A.; McCrea, D.A. Modelling spinal circuitry involved in locomotor pattern generation: Insights from deletions during fictive locomotion. J. Physiol. 2006, 577, 617–639. [Google Scholar] [CrossRef] [PubMed]
- Rybak, I.A.; Stecina, K.; Shevtsova, N.A.; McCrea, D.A. Modelling spinal circuitry involved in locomotor pattern generation: Insights from the effects of afferent stimulation. J. Physiol. 2006, 577, 641–658. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.A.; Stein, P.S. Synaptic control of hindlimb motoneurones during three forms of the fictive scratch reflex in the turtle. J. Physiol. 1988, 404, 101–128. [Google Scholar] [CrossRef] [PubMed]
- Harris-Warrick, R. Locomotor pattern generation in the rodent spinal cord. In Encyclopedia of Computational Neuroscience; Springer: New York, NY, USA, 2013; pp. 1–15. ISBN 978-1-4614-7320-6. [Google Scholar]
- Zhong, G.; Shevtsova, N.A.; Rybak, I.A.; Harris-Warrick, R.M. Neuronal activity in the isolated mouse spinal cord during spontaneous deletions in fictive locomotion: Insights into locomotor central pattern generator organization. J. Physiol. 2012, 590, 4735–4759. [Google Scholar] [CrossRef] [PubMed]
- Shevtsova, N.A.; Hamade, K.; Chakrabarty, S.; Markin, S.N.; Prilutsky, B.I.; Rybak, I.A. Modeling the organization of spinal cord neural circuits controlling two-joint muscles. In Neuromechanical Modeling of Posture and Locomotion; Springer: New York, NY, USA, 2016; pp. 121–162. ISBN 978-1-4939-3267-2. [Google Scholar]
- Shevtsova, N.A.; Rybak, I.A. Organization of flexor–extensor interactions in the mammalian spinal cord: Insights from computational modelling. J. Physiol. 2016, 594, 6117–6131. [Google Scholar] [CrossRef] [PubMed]
- Markin, S.N.; Klishko, A.N.; Shevtsova, N.A.; Lemay, M.A.; Prilutsky, B.I.; Rybak, I.A. A neuromechanical model of spinal control of locomotion. In Neuromechanical Modeling of Posture and Locomotion; Springer: New York, NY, USA, 2016; pp. 21–65. ISBN 978-1-4939-3267-2. [Google Scholar]
- D’Avella, A.; Saltiel, P.; Bizzi, E. Combinations of muscle synergies in the construction of a natural motor behavior. Nat. Neurosci. 2003, 6, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Markin, S.N.; Lemay, M.A.; Prilutsky, B.I.; Rybak, I.A. Motoneuronal and muscle synergies involved in cat hindlimb control during fictive and real locomotion: A comparison study. J. Neurophysiol. 2012, 107, 2057–2071. [Google Scholar] [CrossRef] [PubMed]
- Ting, L.H.; McKay, J.L. Neuromechanics of muscle synergies for posture and movement. Curr. Opin. Neurobiol. 2007, 17, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Tresch, M.C.; Jarc, A. The case for and against muscle synergies. Curr. Opin. Neurobiol. 2009, 19, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Hart, C.B.; Giszter, S.F. Distinguishing synchronous and time-varying synergies using point process interval statistics: Motor primitives in frog and rat. Front. Comput. Neurosci. 2013, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.; Schmidt, M.; Fischer, M.; Quinn, R. A biologically based neural system coordinates the joints and legs of a tetrapod. Bioinspir. Biomim. 2015, 10, 055004. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Hunt, A.J.; Quinn, R.D. Design process and tools for dynamic neuromechanical models and robot controllers. Biol. Cybern. 2017, 111, 105–127. [Google Scholar] [CrossRef] [PubMed]
- Cofer, D.; Cymbalyuk, G.; Reid, J.; Zhu, Y.; Heitler, W.J.; Edwards, D.H. AnimatLab: A 3D graphics environment for neuromechanical simulations. J. Neurosci. Methods 2010, 187, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lanuza, G.M.; Britz, O.; Wang, Z.; Siembab, V.C.; Zhang, Y.; Velasquez, T.; Alvarez, F.J.; Frank, E.; Goulding, M. V1 and V2b interneurons secure the alternating flexor-extensor motor activity mice require for limbed locomotion. Neuron 2014, 82, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Mussa-Ivaldi, F.A.; Bizzi, E. Motor learning through the combination of primitives. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 1755–1769. [Google Scholar] [CrossRef] [PubMed]
- Szczecinski, N.S.; Quinn, R.D.; Hunt, A.J. Parallel relationships between a nonspiking neuron model and a generalized linear integrate and fire model. submitted.
- Prilutsky, B.I.; Klishko, A.N.; Weber, D.J.; Lemay, M.A. Computing motion dependent afferent activity during cat locomotion using a forward dynamics musculoskeletal model. In Neuromechanical Modeling of Posture and Locomotion; Springer: New York, NY, USA, 2016; pp. 273–307. ISBN 978-1-4939-3267-2. [Google Scholar]
- Young, F.; Rode, C.; Hunt, A.; Quinn, R. Analyzing moment arm profiles in a full-muscle rat hindlimb model. Biomimetics 2019, 4, 10. [Google Scholar] [CrossRef]
- Aoi, S.; Kondo, T.; Hayashi, N.; Yanagihara, D.; Aoki, S.; Yamaura, H.; Ogihara, N.; Funato, T.; Tomita, N.; Senda, K.; et al. Contributions of phase resetting and interlimb coordination to the adaptive control of hindlimb obstacle avoidance during locomotion in rats: A simulation study. Biol. Cybern. 2013, 107, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Lynn, R.; Talbot, J.A.; Morgan, D.L. Differences in rat skeletal muscles after incline and decline running differences in rat skeletal muscles after incline and decline running. J. Appl. Physiol. 1998, 85, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Hiebert, G.W.; Pearson, K.G. Contribution of sensory feedback to the generation of extensor activity during walking in the decerebrate cat. J. Neurophysiol. 1999, 81, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Ivashko, D.G.; Prilutsky, B.I.; Markin, S.N.; Chapin, J.K.; Rybak, I.A. Modeling the spinal cord neural circuitry controlling cat hindlimb movement during locomotion. Neurocomputing 2003, 52–54, 621–629. [Google Scholar] [CrossRef]













© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, K.; Szczecinski, N.S.; Arnold, D.; Andrada, E.; Fischer, M.S.; Quinn, R.D.; Hunt, A.J. Neuromechanical Model of Rat Hindlimb Walking with Two-Layer CPGs. Biomimetics 2019, 4, 21. https://doi.org/10.3390/biomimetics4010021
Deng K, Szczecinski NS, Arnold D, Andrada E, Fischer MS, Quinn RD, Hunt AJ. Neuromechanical Model of Rat Hindlimb Walking with Two-Layer CPGs. Biomimetics. 2019; 4(1):21. https://doi.org/10.3390/biomimetics4010021
Chicago/Turabian StyleDeng, Kaiyu, Nicholas S. Szczecinski, Dirk Arnold, Emanuel Andrada, Martin S. Fischer, Roger D. Quinn, and Alexander J. Hunt. 2019. "Neuromechanical Model of Rat Hindlimb Walking with Two-Layer CPGs" Biomimetics 4, no. 1: 21. https://doi.org/10.3390/biomimetics4010021
APA StyleDeng, K., Szczecinski, N. S., Arnold, D., Andrada, E., Fischer, M. S., Quinn, R. D., & Hunt, A. J. (2019). Neuromechanical Model of Rat Hindlimb Walking with Two-Layer CPGs. Biomimetics, 4(1), 21. https://doi.org/10.3390/biomimetics4010021