
The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review


Abstract
:1. Introduction
2. Involvement of Extracellular Matrix in Normal Wound Healing
3. Clotting and Coagulation (Hemostasis)
4. Inflammation
5. Proliferation
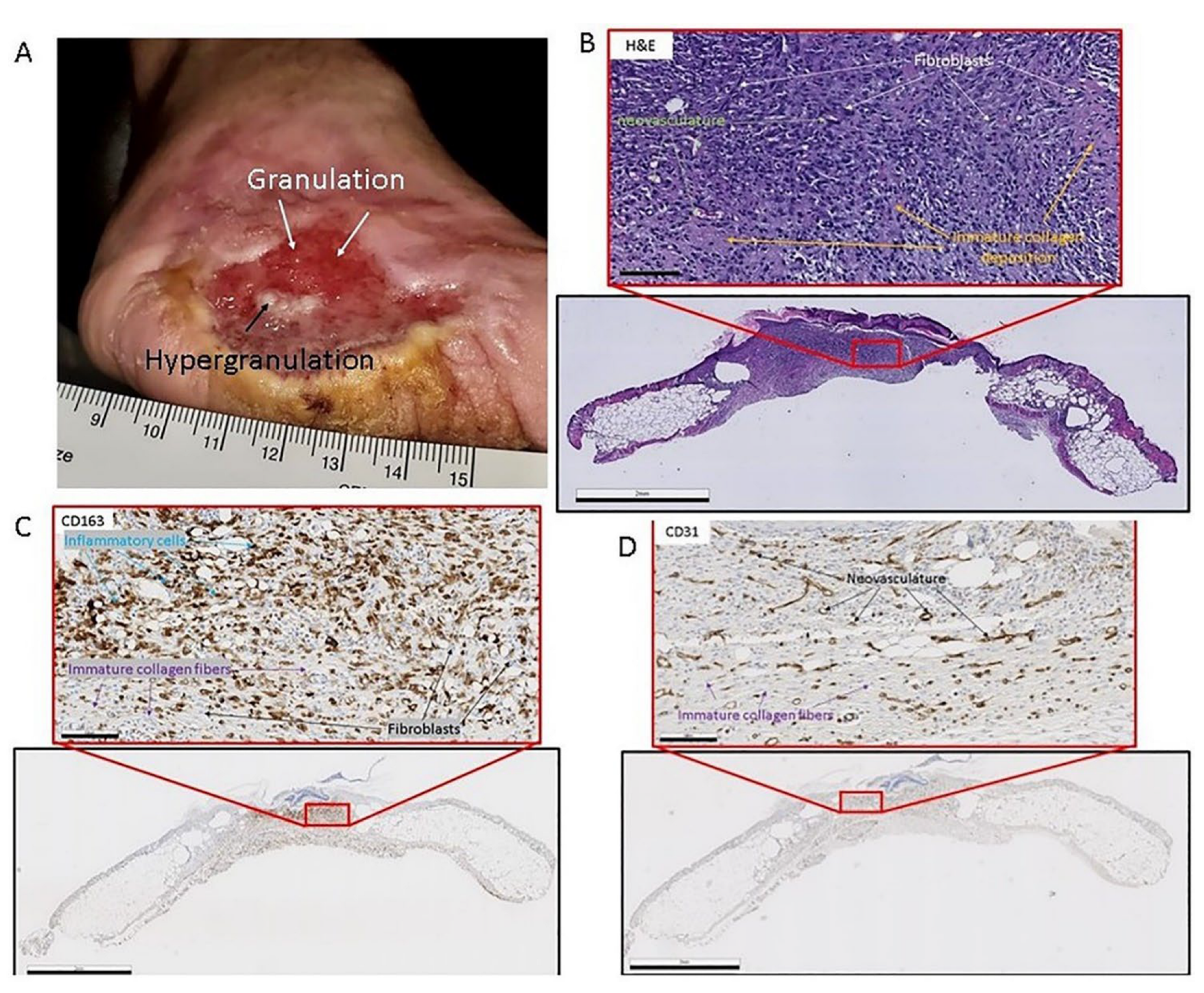
6. Remodeling
7. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Childs, D.R.; Murthy, A.S. Overview of wound healing and management. Surg. Clin. 2017, 97, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.A.; DiPietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Sanford, J.A.; Gallo, R.L. Functions of the skin microbiota in health and disease. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2013; Volume 25, pp. 370–377. [Google Scholar]
- Robson, M.C.; Steed, D.L.; Franz, M.G. Wound healing: Biologic features and approaches to maximize healing trajectories. Curr. Probl. Surg. 2001, 38, 72–140. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M. Biological responses to materials. Annu. Rev. Mater. Res. 2001, 31, 81–110. [Google Scholar] [CrossRef]
- Stevens, M.M.; George, J.H. Exploring and engineering the cell surface interface. Science 2005, 310, 1135–1138. [Google Scholar] [CrossRef]
- Traversa, E.; Mecheri, B.; Mandoli, C.; Soliman, S.; Rinaldi, A.; Licoccia, S.; Forte, G.; Pagliari, F.; Pagliari, S.; Carotenuto, F.; et al. Tuning hierarchical architecture of 3D polymeric scaffolds for cardiac tissue engineering. J. Exp. Nanosci. 2008, 3, 97–110. [Google Scholar] [CrossRef]
- Sen, C.K. Human wound and its burden: Updated 2020 compendium of estimates. Adv. Wound Care 2021, 10, 281–292. [Google Scholar] [CrossRef]
- Enoch, S.; Leaper, D.J. Basic science of wound healing. Surgery 2008, 26, 31–37. [Google Scholar]
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef]
- Alhajj, M.; Goyal, A. Physiology, granulation tissue. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Midwood, K.S.; Williams, L.V.; Schwarzbauer, J.E. Tissue repair and the dynamics of the extracellular matrix. Int. J. Biochem. Cell Biol. 2004, 36, 1031–1037. [Google Scholar] [CrossRef]
- Kushwaha, A.; Goswami, L.; Kim, B.S. Nanomaterial-based therapy for wound healing. Nanomaterials 2022, 12, 618. [Google Scholar] [CrossRef]
- Hess, C.T.; Kirsner, R.S. Orchestrating wound healing: Assessing and preparing the wound bed. Adv. Ski. Wound Care 2003, 16, 246–257. [Google Scholar] [CrossRef]
- Gantwerker, E.A.; Hom, D.B. Skin: Histology and physiology of wound healing. Clin. Plast. Surg. 2012, 39, 85–97. [Google Scholar] [CrossRef]
- Carr, N.J. The pathology of healing and repair. Surgery 2021, 40, 13–19. [Google Scholar] [CrossRef]
- Schultz, G.S.; Wysocki, A. Interactions between extracellular matrix and growth factors in wound healing. Wound Repair Regen. 2009, 17, 153–162. [Google Scholar] [CrossRef]
- Mustoe, T. Understanding chronic wounds: A unifying hypothesis on their pathogenesis and implications for therapy. Am. J. Surg. 2004, 187, S65–S70. [Google Scholar] [CrossRef]
- Brett, D. A Review of Collagen and Collagen-based Wound Dressings. Wounds 2008, 20, 347–356. [Google Scholar]
- Mees, J.; Mardin, W.A.; Senninger, N.; Bruewer, M.; Palmes, D.; Mees, S.T. Treatment options for postoperatively infected abdominal wall wounds healing by secondary intention. Langenbeck’s Arch. Surg. 2012, 397, 1359–1366. [Google Scholar] [CrossRef]
- Yamaoka, H.; Sumiyoshi, H.; Higashi, K.; Nakao, S.; Minakawa, K.; Sumida, K.; Saito, K.; Ikoma, N.; Mabuchi, T.; Ozawa, A.; et al. A novel small compound accelerates dermal wound healing by modifying infiltration, proliferation and migration of distinct cellular components in mice. J. Dermatol. Sci. 2014, 74, 204–213. [Google Scholar] [CrossRef]
- Nair, H.K.; Ahmad, N.W.; Lee, H.L.; Ahmad, N.; Othamn, S.; Mokhtar, N.S.H.M.; Chong, S.S.Y. Hirudotherapy in wound healing. Int. J. Low. Extrem. Wounds 2020. [Google Scholar] [CrossRef]
- Barnes, C.P.; Sell, S.A.; Boland, E.D.; Simpson, D.G.; Bowlin, G.L. Nanofiber technology: Designing the next generation of tissue engineering scaffolds. Adv. Drug Deliv. Rev. 2007, 59, 1413–1433. [Google Scholar] [CrossRef] [PubMed]
- Bonvallet, P.P.; Culpepper, B.K.; Bain, J.L.; Schultz, M.J.; Thomas, S.J.; Bellis, S.L. Microporous dermal-like electrospun scaffolds promote accelerated skin regeneration. Tissue Eng. Part A 2014, 20, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norouzi, M.; Boroujeni, S.M.; Omidvarkordshouli, N.; Soleimani, M. Advances in skin regeneration: Application of electrospun scaffolds. Adv. Healthc. Mater. 2015, 4, 1114–1133. [Google Scholar] [CrossRef] [PubMed]
- Bargavi, P.; Ramya, R.; Chitra, S.; Vijayakumari, S.; Chandran, R.R.; Durgalakshmi, D.; Rajashree, P.; Balakumar, S. Bioactive, degradable and multi-functional three-dimensional membranous scaffolds of bioglass and alginate composites for tissue regenerative applications. Biomater. Sci. 2020, 8, 4003–4025. [Google Scholar] [CrossRef] [PubMed]
- Underwood, P.; Cardinal, P.; Keller, E.; Goodfellow, R.; Scalea, T.; Henry, S.; Lauerman, M.H. Extending Limb Salvage after Fourth and Fifth Transmetatarsal Amputation in Diabetic Foot Infections Using ACell® Urinary Bladder Matrix. Am. Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, R.V.; James, S.L.; James, S.E. A review of tissue-engineered skin bioconstructs available for skin reconstruction. J. R. Soc. Interface 2010, 7, 229–258. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.; Kim, H.M.; Wang, Z.; Bingham, E.L.; Miyazawa, A.; Marcelo, C.L.; Feinberg, S.E. Comparison of two decellularized dermal equivalents. J. Tissue Eng. Regen. Med. 2018, 12, 983–990. [Google Scholar] [CrossRef]
- Tierney, B.P. Comparison of 30-day Clinical Outcomes with SimpliDerm and AlloDerm RTU in Immediate Breast Reconstruction. Plast. Reconstr. Surg. Glob. Open 2021, 9, e3648. [Google Scholar] [CrossRef]
- Vo, A.; Diller, R.; Kellar, R. Characterization and Clinical Applications of Amniotic Membranes. J. Pharmacol. Clin. Res. 2017, 4. [Google Scholar] [CrossRef]
- Valentin, J.E.; Badylak, J.S.; McCabe, G.P.; Badylak, S.F. Extracellular matrix bioscaffolds for orthopaedic applications: A comparative histologic study. JBJS 2006, 88, 2673–2686. [Google Scholar] [CrossRef]
- Chocarro-Wrona, C.; López-Ruiz, E.; Perán, M.; Gálvez-Martín, P.; Marchal, J.A. Therapeutic strategies for skin regeneration based on biomedical substitutes. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 484–496. [Google Scholar] [CrossRef]
- Lisa, A.V.E.; Galtelli, L.; Vinci, V.; Veronesi, A.; Cozzaglio, L.; Cananzi, F.C.M.; Sicoli, F.; Klinger, M. Adoption of a newly introduced dermal matrix: Preliminary experience and future directions. BioMed Res. Int. 2020, 2020, 3261318. [Google Scholar] [CrossRef]
- Dai, C.; Shih, S.; Khachemoune, A. Skin substitutes for acute and chronic wound healing: An updated review. J. Dermatol. Treat. 2020, 31, 639–648. [Google Scholar] [CrossRef]
- Yeh, D.D.; Nazarian, R.M.; Demetri, L.; Mesar, T.; Dijkink, S.; Larentzakis, A.; Velmahos, G.; Sadik, K.W. Histopathological assessment of OASIS ultra on critical-sized wound healing: A pilot study. J. Cutan. Pathol. 2017, 44, 523–529. [Google Scholar] [CrossRef]
- Kellar, R.S.; Diller, R.B.; Tabor, A.J.; Dominguez, D.D.; Audet, R.G.; Bardsley, T.A.; Talbert, A.J.; Cruz, N.D.; Ingraldi, A.L.; Ensley, B.D. Improved wound closure rates and mechanical properties resembling native skin in murine diabetic wounds treated with a tropoelastin and collagen wound healing device. J. Diabetes Clin. Res. 2020, 2, 86. [Google Scholar]
- Hsu, P.W.; Salgado, C.J.; Kent, K.; Finnegan, M.; Pello, M.; Simons, R.; Atabek, U.; Kann, B. Evaluation of porcine dermal collagen (Permacol) used in abdominal wall reconstruction. J. Plast. Reconstr. Aesthetic Surg. 2009, 62, 1484–1489. [Google Scholar] [CrossRef]
- Veves, A.; Sheehan, P.; Pham, H.T. A randomized, controlled trial of Promogran (a collagen/oxidized regenerated cellulose dressing) vs. standard treatment in the management of diabetic foot ulcers. Arch. Surg. 2002, 137, 822–827. [Google Scholar] [CrossRef]
- Thakar, H.; Sebastian, S.M.; Mandal, S.; Pople, A.; Agarwal, G.; Srivastava, A. Biomolecule-conjugated macroporous hydrogels for biomedical applications. ACS Biomater. Sci. Eng. 2019, 5, 6320–6341. [Google Scholar] [CrossRef]
- Patil, P.; Russo, K.A.; McCune, J.T.; Pollins, A.C.; Cottam, M.A.; Dollinger, B.R.; Colazo, J.M.; Yu, F.; Martin, J.R.; Gupta, M.K.; et al. ROS-Degradable Polythioketal Urethane Foam Dressings to Promote Porcine Skin Wound Repair. bioRxiv 2021. [Google Scholar] [CrossRef]
- Puhl, D.L.; Mohanraj, D.; Nelson, D.W.; Gilbert, R.J. Designing electrospun fiber platforms for efficient delivery of genetic material and genome editing tools. Adv. Drug Deliv. Rev. 2022, 183, 114161. [Google Scholar] [CrossRef]
- Clark, R.A.; Ashcroft, G.S.; Spencer, M.J.; Larjava, H.; Ferguson, M.W.J. Re-epithelialization of normal human excisional wounds is associated with a switch from αvβ5 to αvβ6 integrins. Br. J. Dermatol. 1996, 135, 46–51. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef]
- Ellis, S.; Lin, E.J.; Tartar, D. Immunology of wound healing. Curr. Dermatol. Rep. 2018, 7, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Becerra-Bayona, S.M.; Solarte-David, V.A.; Sossa, C.L.; Mateus, L.C.; Villamil, M.; Pereira, J.; Arango-Rodríguez, M.L. Mesenchymal Stem Cells Derivatives as a Novel and Potential Therapeutic Approach to Treat Diabetic Foot Ulcers. Endocrinol. Diabetes Metab. Case Rep. 2020, 2020, 19–0164. [Google Scholar] [CrossRef]
- Wallace, H.A.; Basehore, B.M.; Zito, P.M. Wound Healing Phases; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Lukashev, M.E.; Werb, Z. ECM signalling: Orchestrating cell behaviour and misbehaviour. Trends Cell Biol. 1998, 8, 437–441. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Pfisterer, K.; Shaw, L.E.; Symmank, D.; Weninger, W. The extracellular matrix in skin inflammation and infection. Front. Cell Dev. Biol. 2021, 9, 682414. [Google Scholar] [CrossRef] [PubMed]
- David, V.A.S.; Güiza-Argüello, V.R.; Arango-Rodríguez, M.L.; Sossa, C.L.; Becerra-Bayona, S.M. Decellularized Tissues for Wound Healing: Towards Closing the Gap Between Scaffold Design and Effective Extracellular Matrix Remodeling. Front. Bioeng. Biotechnol. 2022, 10, 821852. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almine, J.F.; Bax, D.V.; Mithieux, S.M.; Nivison-Smith, L.; Rnjak, J.; Waterhouse, A.; Wise, S.; Weiss, A.S. Elastin-based materials. Chem. Soc. Rev. 2010, 39, 3371–3379. [Google Scholar] [CrossRef] [PubMed]
- Maquart, F.X.; Monboisse, J.C. Extracellular matrix and wound healing. Pathol. Biol. 2014, 62, 91–95. [Google Scholar] [CrossRef]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular matrix and dermal fibroblast function in the healing wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Rousselle, P.; Montmasson, M.; Garnier, C. Extracellular matrix contribution to skin wound re-epithelialization. Matrix Biol. 2019, 75, 12–26. [Google Scholar] [CrossRef]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- Fidler, A.L.; Boudko, S.P.; Rokas, A.; Hudson, B.G. The triple helix of collagens–an ancient protein structure that enabled animal multicellularity and tissue evolution. J. Cell Sci. 2018, 131, jcs203950. [Google Scholar] [CrossRef] [Green Version]
- Har-Shai, Y.; Sela, E.; Rubinstien, I.; Lindenbaum, E.S.; Mitz, V.; Hirshowitz, B. Computerized morphometric quantitation of elastin and collagen in SMAS and facial skin and the possible role of fat cells in SMAS viscoelastic properties. Plast. Reconstr. Surg. 1998, 102, 2466–2470. [Google Scholar] [CrossRef]
- Uitto, J.; Olsen, D.R.; Fazio, M.J. Extracellular matrix of the skin: 50 years of progress. J. Investig. Dermatol. 1989, 92, S61–S77. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Garva, R.; Pickard, A.; Yeung, C.Y.C.; Mallikarjun, V.; Swift, J.; Holmes, D.F.; Calverley, B.; Lu, Y.; Adamson, A.; et al. Circadian control of the secretory pathway maintains collagen homeostasis. Nat. Cell Biol. 2020, 22, 74–86. [Google Scholar] [CrossRef]
- Weinstein, G.D.; Boucek, R.J. Collagen and elastin of human dermis. J. Investig. Dermatol. 1960, 35, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Uitto, J.; Paul, J.L.; Brockley, K.; Pearce, R.H.; Clark, J.G. Elastic fibers in human skin: Quantitation of elastic fibers by computerized digital image analyses and determination of elastin by radioimmunoassay of desmosine. Lab. Investig. J. Tech. Methods Pathol. 1983, 49, 499–505. [Google Scholar]
- Matsuoka, L.Y.; Wortsman, J.; Uitto, J.; Hashimoto, K.; Kupchella, C.E.; Eng, A.M.; Dietrich, J.E. Altered skin elastic fibers in hypothyroid myxedema and pretibial myxedema. Arch. Intern. Med. 1985, 145, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sephel, G.C.; Davidson, J.M. Elastin production in human skin fibroblast cultures and its decline with age. J. Investig. Dermatol. 1986, 86, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Nieves, D.; Chaikof, E.L. Collagen and Elastin Biomaterials for the Fabrication of Engineered Living Tissues. ACS Biomater. Sci. Eng. 2017, 3, 694–711. [Google Scholar] [CrossRef]
- Amadeu, T.P.; Braune, A.S.; Porto, L.C.; Desmoulière, A.; Costa, A.M.A. Fibrillin-1 and elastin are differentially expressed in hypertrophic scars and keloids. Wound Repair Regen. 2004, 12, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Andia, I.; Ardanza, B.; Nurden, P.; Nurden, A.T. Autologous platelets as a source of proteins for healing and tissue regeneration. Thromb. Haemost. 2004, 91, 4–15. [Google Scholar] [CrossRef]
- Baumann, L.; Bernstein, E.F.; Weiss, A.S.; Bates, D.; Humphrey, S.; Silberberg, M.; Daniels, R. Clinical relevance of elastin in the structure and function of skin. Aesthetic Surg. J. Open Forum 2021, 3, ojab019. [Google Scholar] [CrossRef]
- Swee, M.H.; Parks, W.C.; Pierce, R.A. Developmental Regulation of Elastin Production. Expression of Tropoelastin Pre-Mrna Persists after Down-Regulation of Steady-State mRNA Levels. J. Biol. Chem. 1995, 270, 14899–14906. [Google Scholar] [CrossRef] [Green Version]
- Mecham, R.P. Regulation of Matrix Accumulation; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Davidson, J.M. Biochemistry and turnover of lung interstitium. Eur. Respir. J. 1990, 3, 1048–1063. [Google Scholar]
- Rucker, R.B.; Dubick, M.A. Elastin metabolism and chemistry: Potential roles in lung development and structure. Environ. Health Perspect. 1984, 55, 179. [Google Scholar] [CrossRef]
- Shapiro, S.D.; Endicott, S.K.; Province, M.A.; Pierce, J.A.; Campbell, E.J. Marked longevity of human lung parenchymal elastic fibers deduced from prevalence of D-aspartate and nuclear weapons-related radiocarbon. J. Clin. Investig. 1991, 87, 1828. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, P.; Pulcinelli, F.M.; Lenti, L.; Gazzaniga, P.P.; Violi, F. Hydrogen peroxide is involved in collagen-induced platelet activation. Blood 1998, 91, 484–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, A.R.; Sands, D.; Sandberg, E.; Seitz, R.; Barrowcliffe, T.W. A multi-centre collaborative study on the potency estimation of ReFacto. Thromb. Haemost. 2003, 90, 1088–1093. [Google Scholar] [PubMed]
- Diegelmann, R.F.; Evans, M.C. The wound healing process: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef]
- Broughton, G.; Janis, J.E.; Attinger, C.E. The basic science of wound healing. Plast. Reconstr. Surg. 2006, 117 (Suppl. 7), 12S–34S. [Google Scholar] [CrossRef]
- Blair, P.; Flaumenhaft, R. Platelet α-granules: Basic biology and clinical correlates. Blood Rev. 2009, 23, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Montgomery, R.R. Platelets as delivery systems for disease treatments. Adv. Drug Deliv. Rev. 2010, 62, 1196–1203. [Google Scholar] [CrossRef] [Green Version]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Clark, R.A. Fibrin and wound healing. Ann. N. Y. Acad. Sci. 2001, 936, 355–367. [Google Scholar] [CrossRef]
- Adams, S.A.; Kelly, S.L.; Kirsch, R.E.; Robson, S.C.; Shephard, E.G. Role of neutrophil membrane proteases in fibrin degradation. Blood Coagul. Fibrinolysis 1995, 6, 693–702. [Google Scholar] [CrossRef]
- Ahmann, K.A.; Weinbaum, J.S.; Johnson, S.L.; Tranquillo, R.T. Fibrin degradation enhances vascular smooth muscle cell proliferation and matrix deposition in fibrin-based tissue constructs fabricated in vitro. Tissue Eng. Part A 2010, 16, 3261–3270. [Google Scholar] [CrossRef] [Green Version]
- Gray, A.J.; Bishop, J.E.; Reeves, J.T.; Mecham, R.P.; Laurent, G.J. Partially degraded fibrin (ogen) stimulates fibroblast proliferation in vitro. Am. J. Respir. Cell Mol. Biol. 1995, 12, 684–690. [Google Scholar] [CrossRef]
- Dvorak, H.F.; Harvey, V.S.; Estrella, P.; Brown, L.F.; McDonagh, J.; Dvorak, A.M. Fibrin containing gels induce angiogenesis. Implications for tumor stroma generation and wound healing. Lab. Investig. 1987, 57, 673–686. [Google Scholar]
- Pierce, G.F.; Mustoe, T.A.; Lingelbach, J.; Masakowski, V.R.; Griffin, G.L.; Senior, R.M.; Deuel, T.F. Platelet-derived growth factor and transforming growth factor-beta enhance tissue repair activities by unique mechanisms. J. Cell Biol. 1989, 109, 429–440. [Google Scholar] [CrossRef]
- Pierce, G.F.; Mustoe, T.A.; Altrock, B.W.; Deuel, T.F.; Thomason, A. Role of platelet-derived growth factor in wound healing. J. Cell. Biochem. 1991, 45, 319–326. [Google Scholar] [CrossRef]
- Kim, L.R.; Whelpdale, K.; Zurowski, M.; Pomeranz, B. Sympathetic denervation impairs epidermal healing in cutaneous wounds. Wound Repair Regen. 1998, 6, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.M.; Hunt, D.A.; Wakefield, L.M.; McCartney-Francis, N.; Wahl, L.M.; Roberts, A.B.; Sporn, M.B. Transforming growth factor type beta induces monocyte chemotaxis and growth factor production. Proc. Natl. Acad. Sci. USA 1987, 84, 5788–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ather, S.; Harding, K.G.; Tate, S.J. Wound management and dressings. In Advanced Textiles for Wound Care; Woodhead Publishing: Sawston, UK, 2019; pp. 1–22. [Google Scholar]
- Clark, R.A.; Wikner, N.E.; Doherty, D.E.; Norris, D.A. Cryptic chemotactic activity of fibronectin for human monocytes resides in the 120-kDa fibroblastic cell-binding fragment. J. Biol. Chem. 1988, 263, 12115–12123. [Google Scholar] [CrossRef]
- Brown, E.J.; Goodwin, J.L. Fibronectin receptors of phagocytes. J. Exp. Med. 1988, 167, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A. Regulation of fibroplasia in cutaneous wound repair. Am. J. Med. Sci. 1993, 306, 42–48. [Google Scholar] [CrossRef]
- Shaw, R.J.; Doherty, D.E.; Ritter, A.G.; Benedict, S.H.; Clark, R.A.F. Adherence-dependent increase in human monocyte PDGF (B) mRNA is associated with increases in c-fos, c-jun, and EGR2 mRNA. J. Cell Biol. 1990, 111, 2139–2148. [Google Scholar] [CrossRef]
- Mascharak, S.; desJardins-Park, H.E.; Davitt, M.F.; Griffin, M.; Borrelli, M.R.; Moore, A.L.; Chen, K.; Duoto, B.; Chinta, M.; Foster, D.S.; et al. Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring. Science 2021, 372, eaba2374. [Google Scholar] [CrossRef]
- Lin, F.; Ren, X.D.; Doris, G.; Clark, R.A. Three-dimensional migration of human adult dermal fibroblasts from collagen lattices into fibrin/fibronectin gels requires syndecan-4 proteoglycan. J. Investig. Dermatol. 2005, 124, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.; Young, A.; McNaught, C.E. The physiology of wound healing. Surgery 2014, 32, 445–450. [Google Scholar] [CrossRef]
- Mu, X.; Li, Y.; Fan, G.C. Tissue-resident macrophages in the control of infection and resolution of inflammation. Shock 2021, 55, 14–23. [Google Scholar] [CrossRef]
- Mast, B.A.; Diegelmann, R.F.; Krummel, T.M.; Cohen, I.K. Scarless wound healing in the mammalian fetus. Surg. Gynecol. Obstet. 1992, 174, 441–451. [Google Scholar]
- Pohlman, T.H.; Stanness, K.A.; Beatty, P.G.; Ochs, H.D.; Harlan, J.M. An endothelial cell surface factor(s) induced in vitro by lipopolysaccharide, interleukin 1, and tumor necrosis factor-alpha increases neutrophil adherence by a CDw18-dependent mechanism. J. Immunol. 1986, 136, 4548–4553. [Google Scholar]
- Young, A.; McNaught, C.E. The physiology of wound healing. Surgery 2011, 29, 475–479. [Google Scholar] [CrossRef]
- Martin, U.; Bock, D.; Arseniev, L.; Tornetta, M.A.; Ames, R.S.; Bautsch, W.; Köhl, J.; Ganser, A.; Klos, A. The human C3a receptor is expressed on neutrophils and monocytes, but not on B or T lymphocytes. J. Exp. Med. 1997, 186, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Parani, M.; Lokhande, G.; Singh, A.; Gaharwar, A.K. Engineered nanomaterials for infection control and healing acute and chronic wounds. ACS Appl. Mater. Interfaces 2016, 8, 10049–10069. [Google Scholar] [CrossRef]
- Bar-Shavit, R.; Kahn, A.; Fenton, J.W.; Wilner, G.D. Chemotactic response of monocytes to thrombin. J. Cell Biol. 1983, 96, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Janis, J.; Harrison, B. Wound healing: Part II. Clinical applications. Plast. Reconstr. Surg. 2014, 133, 383e–392e. [Google Scholar] [CrossRef]
- Viaña-Mendieta, P.; Sánchez, M.L.; Benavides, J. Rational selection of bioactive principles for wound healing applications: Growth factors and antioxidants. Int. Wound J. 2022, 19, 100–113. [Google Scholar] [CrossRef]
- Pierce, G.F.; Berg, J.V.; Rudolph, R.; Tarpley, J.; Mustoe, T.A. Platelet-derived growth factor-BB and transforming growth factor beta 1 selectively modulate glycosaminoglycans, collagen, and myofibroblasts in excisional wounds. Am. J. Pathol. 1991, 138, 629. [Google Scholar]
- Ramasastry, S.S. Acute wounds. Clin. Plast. Surg. 2005, 32, 195–208. [Google Scholar] [CrossRef]
- Szpaderska, A.M.; Zuckerman, J.D.; DiPietro, L.A. Differential injury responses in oral mucosal and cutaneous wounds. J. Dent. Res. 2003, 82, 621–626. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Baram, D.; Mekori, Y.A. Mast cells. Physiol. Rev. 1997, 77, 1033–1079. [Google Scholar] [CrossRef]
- Artuc, M.; Hermes, B.; Stckelings, U.M.; Grützkau, A.; Henz, B.M. Mast cells and their mediators in cutaneous wound healing? active participants or innocent bystanders? Exp. Dermatol. 1999, 8, 1–16. [Google Scholar] [CrossRef]
- Martin, P.; Leibovich, S.J. Inflammatory cells during wound repair: The good, the bad and the ugly. Trends Cell Biol. 2005, 15, 599–607. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A review of the contribution of mast cells in wound healing: Involved molecular and cellular mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef]
- Gethin, G. Understanding the inflammatory process in wound healing. Br. J. Community Nurs. 2012, 17. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Tonnesen, M.G.; Feng, X.; Clark, R.A. Angiogenesis in wound healing. J. Investig. Dermatol. Symp. Proc. 2000, 5, 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eppley, B.L.; Pietrzak, W.S.; Blanton, M. Platelet-rich plasma: A review of biology and applications in plastic surgery. Plast. Reconstr. Surg. 2006, 118, 147e–159e. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häkkinen, L.; Larjava, H.; Koivisto, L. Granulation tissue formation and remodeling. Endod. Top. 2011, 24, 94–129. [Google Scholar] [CrossRef]
- Stadelmann, W.K.; Digenis, A.G.; Tobin, G.R. Physiology and healing dynamics of chronic cutaneous wounds. Am. J. Surg. 1998, 176, 26S–38S. [Google Scholar] [CrossRef]
- Raghunath, M.; Bachi, T.; Meuli, M.; Altermatt, S.; Gobet, R.; Bruckner-Tuderman, L.; Steinmann, B. Fibrillin and elastin expression in skin regenerating from cultured keratinocyte autografts: Morphogenesis of microfibrils begins at the dermo-epidermal junction and precedes elastic fiber formation. J. Investig. Dermatol. 1996, 106, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Sarojini, H.; Bajorek, A.; Wan, R.; Wang, J.; Zhang, Q.; Billeter, A.T.; Chien, S. Enhanced Skin Incisional Wound Healing with Intracellular ATP Delivery via Macrophage Proliferation and Direct Collagen Production. Front. Pharmacol. 2021, 12, 594586. [Google Scholar] [CrossRef]
- Lee, J.H.; Parthiban, P.; Jin, G.Z.; Knowles, J.C.; Kim, H.W. Materials roles for promoting angiogenesis in tissue regeneration. Prog. Mater. Sci. 2021, 117, 100732. [Google Scholar] [CrossRef]
- Noskovicova, N.; Hinz, B.; Pakshir, P. Implant fibrosis and the underappreciated role of myofibroblasts in the foreign body reaction. Cells 2021, 10, 1794. [Google Scholar] [CrossRef]
- Guido, S.; Tranquillo, R.T. A methodology for the systemic and quantitative study of cell contact guidance in oriented collagen gels. J. Cell Sci. 1993, 105, 317–331. [Google Scholar] [CrossRef]
- Gailit, J.; Clark, R.A. Wound repair in the context of extracellular matrix. Curr. Opin. Cell Biol. 1994, 6, 717–725. [Google Scholar] [CrossRef]
- Assunção, M.; Yiu, C.H.K.; Wan, H.Y.; Wang, D.; Tuan, R.S.; Blocki, A. Hyaluronic acid drives mesenchymal stromal cell-derived extracellular matrix assembly by promoting fibronectin fibrillogenesis. J. Mater. Chem. B 2021, 9, 7205–7215. [Google Scholar] [CrossRef]
- Bainbridge, P. Wound healing and the role of fibroblasts. J. Wound Care 2013, 22, 407–411. [Google Scholar]
- Banks, R.E.; Forbes, M.A.; Kinsey, S.E.; Stanley, A.; Ingham, E.; Walters, C.; Selby, P.J. Release of the angiogenic cytokine vascular endothelial growth factor (VEGF) from platelets: Significance for VEGF measurements and cancer biology. Br. J. Cancer 1998, 77, 956. [Google Scholar] [CrossRef] [Green Version]
- DiPietro, L.A. Angiogenesis and scar formation in healing wounds. Curr. Opin. Rheumatol. 2013, 25, 87–91. [Google Scholar] [CrossRef]
- Pettet, G.J.; Byrne, H.M.; McElwain, D.L.S.; Norbury, J. A model of wound-healing angiogenesis in soft tissue. Math. Biosci. 1996, 136, 35–63. [Google Scholar] [CrossRef]
- Kurz, H. Physiology of angiogenesis. J. Neuro-Oncol. 2000, 50, 17–35. [Google Scholar] [CrossRef]
- Hunt, T.K. The physiology of wound healing. Ann. Emerg. Med. 1988, 17, 1265–1273. [Google Scholar] [CrossRef]
- Ghani, Q.P.; Wagner, S.; Becker, H.D.; Hunt, T.K.; Hussain, M.Z. Regulatory role of lactate in wound repair. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2004; Volume 381, pp. 565–575. [Google Scholar]
- Shen, Y.; Jiang, L.; Wen, P.; Ye, Y.; Zhang, Y.; Ding, H.; Luo, J.; Xu, L.; Zen, K.; Zhou, Y.; et al. Tubule-derived lactate is required for fibroblast activation in acute kidney injury. Am. J. Physiol.-Ren. Physiol. 2020, 318, F689–F701. [Google Scholar] [CrossRef]
- Schultz, G.S.; White, M.; Mitchell, R.; Brown, G.; Lynch, J.; Twardzik, D.R.; Todaro, G.J. Epithelial wound healing enhanced by transforming growth factor-alpha and vaccinia growth factor. Science 1987, 235, 350–352. [Google Scholar] [CrossRef]
- Yates, R.A.; Nanney, L.B.; Gates, R.E.; King, L.E. Epidermal growth factor and related growth factors. Int. J. Dermatol. 1991, 30, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Kurkinen, M.; Vaheri, A.N.T.T.I.; Roberts, P.J.; Stenman, S. Sequential appearance of fibronectin and collagen in experimental granulation tissue. Lab. Investig. 1980, 43, 47–51. [Google Scholar] [PubMed]
- Coelho, N.M.; McCulloch, C.A. Contribution of collagen adhesion receptors to tissue fibrosis. Cell Tissue Res. 2016, 365, 521–538. [Google Scholar] [CrossRef]
- Lee, Y.I.; Lee, S.G.; Kim, J.; Choi, S.; Jung, I.; Lee, J.H. Proteoglycan Combined with Hyaluronic Acid and Hydrolyzed Collagen Restores the Skin Barrier in Mild Atopic Dermatitis and Dry, Eczema-Prone Skin: A Pilot Study. Int. J. Mol. Sci. 2021, 22, 10189. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Overall, C.M.; López-Otín, C. Strategies for MMP inhibition in cancer: Innovations for the post-trial era. Nat. Rev. Cancer 2002, 2, 657–672. [Google Scholar] [CrossRef]
- Chen, P.; Parks, W.C. Role of matrix metalloproteinases in epithelial migration. J. Cell. Biochem. 2009, 108, 1233–1243. [Google Scholar] [CrossRef]
- Martins, V.L.; Caley, M.; O’Toole, E.A. Matrix metalloproteinases and epidermal wound repair. Cell Tissue Res. 2013, 351, 255–268. [Google Scholar] [CrossRef]
- Welch, M.P.; Odland, G.F.; Clark, R.A. Temporal relationships of F-actin bundle formation, collagen and fibronectin matrix assembly, and fibronectin receptor expression to wound contraction. J. Cell Biol. 1990, 110, 133–145. [Google Scholar] [CrossRef]
- Kumar, I.; Staton, C.A.; Cross, S.S.; Reed, M.W.R.; Brown, N.J. Angiogenesis, vascular endothelial growth factor and its receptors in human surgical wounds. Br. J. Surg. 2009, 96, 1484–1491. [Google Scholar] [CrossRef]
- Abercrombie, M.; Flint, M.H.; James, D.W. Wound contraction in relation to collagen formation in scorbutic guinea-pigs. Development 1956, 4, 167–175. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Products Manufactured Utilizing ECM Proteins | ||||
|---|---|---|---|---|
| Product Name | Source | Composition | ECM Proteins | Reference(s) |
| Acell Wound Powder | Porcine Urinary Bladder | Powdered Urinary Bladder Extracellular Matrix | Collagen I, Collagen III, and Collagen IV | [27] |
| AlloDerm | Human Dermis | Human acellular, lyophilized dermal matrix | collagen, elastin, basement membrane, hyaluronic acid glycosaminoglycan (GAG) | [28,29,30] |
| AlloPatch | Human Cadaveric Dermis | aseptically processed human reticular dermal tissue | collagen, elastin, basement membrane, hyaluronic acid glycosaminoglycan (GAG) | [29] |
| Axolotl DualGraft | Human Amnion | Dehydrated amniotic membrane | Collagen I, Collagen II, Collagen IV, Collagen V, Collagen VI, Proteoglycans, Fibronectin, Laminin | [31] |
| GraftJacket | Human cadaveric skin | Cryogenicaly preserved decellularized human dermis | Collagen and elastin | [32,33] |
| MatriDerm | Bovine Ligamentum nuchae | 3D matrix of type 1 collagen fiber coated with a 3% a-elastin hydrolysate | collagen fibrils types I, III, and V and alpha-elastin | [34,35] |
| OASIS | Porcine jejunum submucosa (PSIS) | Lyophilized small intestine submucosa | Collagen I, Collagen III, Collagen IV, Collagen VI, fibronectin, elastin, hyaluronan, chondroitin sulfate, decorin | [36,37] |
| Pelnac | Porcine Achilles tendon | Collagen sponge porous matrix | Collagen | [34] |
| PalinGen Membrane | Human Amnion | Dehydrated amniotic membrane | Collagen I, Collagen II, Collagen IV, Collagen V, Collagen VI, Proteoglycans, Fibronectin, Laminin | [31] |
| Permacol | Porcine Dermis | Collagen and Elastin crosslinked by diisocynate | Collagen I, Elastin | [38] |
| Promogran | Bovine Dermis | 55% Bovine Dermal Collagen 45% Oxidized regenerated cellulose | Collagen | [39] |
| SimliDerm | Human Cadaveric Dermis | pre-hydrated human acellular dermal matrix | Collagen, elastin, basement membrane, hyaluronic acid glycosaminoglycan (GAG) | [30] |
| Xcellistem | Porcine Spleen and Lung | wound powder composed of a blend of multiple porcine-based extracellular matrix material | Collagen I, Collagen III, Collagen IV, Sulfated glycosaminoglycan, Hyaluronic acid, Lipids, Elastin, Fibronectin, and Laminin | [40] |
| Novosorb | Synthetic polyers | polyurethane foam made from ethyl lysine diisocyanate, lactic acid/ethylene glycol chain extender, and PCL1000 polyol with a removable polyurethane overlayer | None | [41] |
| Phoenix Wound Matrix | Sythetic polymers | Electrospun nonwoven fibrous three-dimensional matrix comprised of Poly(lactide-co-caprolactone) and Polyglycolic acid fibers | None | [42] |
| Restrata Wound Matrix | Synthetic polymers | electrospun mat of polyglactin 910 (PGLA) and polydiaxonone (PDO) fibers | None | [41] |
| Protein | Percent Composition | Function(s) |
|---|---|---|
| Collagen(s) | 50–90 | Synthesized by fibroblasts. Gives structural and tissue integrity, aids in epidermal/dermal differentiation. |
| Elastin | 0.6–7.9 | Creates an intricate network for structural support allowing for elasticity of tissue. |
| Fibronectin | <1.0 | Involved in wound healing including platelet spreading and leukocyte migration to injured tissue(s). Aids in promotion of elastin deposition and mechanical strength of ECM. |
| Laminin | <1.0 | A glycoprotein that is a part of the basal lamina, aids in cellular signaling. |
| Vitronectin | <1.0 | A glycoprotein involved in hemostasis and cellular adhesion during tissue damage. |
| Tenascin | <1.0 | Glycoprotein family that aids in cellular migration adhesions and cell proliferation. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diller, R.B.; Tabor, A.J. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics 2022, 7, 87. https://doi.org/10.3390/biomimetics7030087
Diller RB, Tabor AJ. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics. 2022; 7(3):87. https://doi.org/10.3390/biomimetics7030087
Chicago/Turabian StyleDiller, Robert B., and Aaron J. Tabor. 2022. "The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review" Biomimetics 7, no. 3: 87. https://doi.org/10.3390/biomimetics7030087
APA StyleDiller, R. B., & Tabor, A. J. (2022). The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics, 7(3), 87. https://doi.org/10.3390/biomimetics7030087