
Optimization of Live Prey Enrichment Media for Rearing Juvenile Short-Snouted Seahorse, Hippocampus hippocampus




Abstract
:1. Introduction
2. Materials and Methods
2.1. Live Feed Culture and Enrichment
2.2. Copepod Harvesting
2.3. Feeding trials
2.4. Sampling, Data Collection, and Statistical Analysis
2.5. Dietary Treatments Proximate Analysis and Fatty Acid Composition
2.6. Ethical Statement
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vincent, A.C.J. The International Trade in Seahorses; TRAFFIC International: Cambridge, UK, 1996; ISBN 1 85850 098 2. [Google Scholar]
- Koldewey, H.J.; Martin-Smith, K.M. A global review of seahorse aquaculture. Aquaculture 2010, 302, 131–152. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021.1. Available online: http://www.iucnredlist.org (accessed on 4 August 2023).
- Pollom, R. Hippocampus hippocampus. In: The IUCN Red List of Threatened Species 2016: e.T10069A54904826. 2016. Available online: https://www.iucnredlist.org/species/10069/67618259(accessed on 2 August 2023).
- Otero-Ferrer, F.; Molina, L.; Socorro, J.; Herrera, R. Live prey first feeding regimes for short-snouted seahorse Hippocampus hippocampus (Linnaeus, 1758) juveniles. Aquacult. Res. 2010, 41, 8–19. [Google Scholar] [CrossRef]
- Leus, K. Captive breeding and conservation. Zool. Middle East 2011, 54 (Suppl. 3), 151–158. [Google Scholar] [CrossRef]
- Olivotto, I.; Planas, M.; Simões, N.; Holt, G.J.; Avella, M.A.; Calado, R. Advances in breeding and rearing marine ornamentals. J. World Aquac. Soc. 2011, 42, 135–166. [Google Scholar] [CrossRef]
- Rønnestad, I.; Thorsen, A.; Finn, R.N. Fish larval nutrition: A review of recent advances in the roles of amino acids. Aquaculture 1999, 177, 201–216. [Google Scholar] [CrossRef]
- Barclay, W.; Zeller, S. Nutritional enrichment of n-3 and n-6 fatty acids in rotifers and Artemia nauplii by feeding spray-dried Schizochytrium sp. J. World Aquac. Soc. 1996, 27, 314–322. [Google Scholar] [CrossRef]
- Furuita, H.; Konishi, K.; Takeuchi, T. Effect of different levels of eicosapentaenoic acid and docosahexaenoic acid in Artemia nauplii on growth, survival and salinity tolerances of larvae of the Japanese flounder, Paralichthys olivaceus. Aquaculture 1999, 170, 59–69. [Google Scholar] [CrossRef]
- Sorgeloos, P.; Dhert, P.; Candreva, P. Use of the brine shrimp, Artemia spp., in marine fish larviculture. Aquaculture 2001, 200, 147–215. [Google Scholar] [CrossRef]
- Watanabe, T. Nutritional quality of brine shrimp, Artemia salina, as a living feed from the viewpoint of essential fatty acids for fish. Bull. Japan Soc. Sci. Fish. 1978, 44, 1115. [Google Scholar] [CrossRef]
- Watanabe, T. Lipid nutrition in fish. Comp. Biochem. Physiol. 1982, 73, 3–15. [Google Scholar] [CrossRef]
- Job, S.; Do, H.; Meeuwig, J.; Hall, H. Culturing the oceanic seahorse, Hippocampus kuda. Aquaculture 2002, 214, 333–334. [Google Scholar] [CrossRef]
- Martínez-Cárdenas, L.; Purser, G.J. Effect of tank colour on Artemia ingestion, growth and survival in cultured early juvenile pot-bellied seahorses (Hippocampus abdominalis). Aquaculture 2007, 26, 92–100. [Google Scholar] [CrossRef]
- Olivotto, I.; Avella, M.A.; Sampaolesi, G.; Piccinetti, C.C.; Ruiz, P.N.; Carnevali, O. Breeding and rearing the longsnout seahorse Hippocampus reidi: Rearing and feeding studies. Aquaculture 2008, 283, 92–96. [Google Scholar] [CrossRef]
- Palma, J.; Bureau, D.P.; Andrade, J.P. Effect of different Artemia enrichments and feeding protocol for rearing juvenile long snout seahorse, Hippocampus guttulatus. Aquaculture 2011, 318, 439–443. [Google Scholar] [CrossRef]
- Payne, M.F.; Rippingale, R.J. Rearing West Australian seahorse, Hippocampus subelongatus, juveniles on copepod nauplii and enriched Artemia. Aquaculture 2000, 188, 353–361. [Google Scholar] [CrossRef]
- Sheng, J.; Lin, Q.; Chen, Q.; Gao, Y.; Shen, L.; Lu, J. Effects of food, temperature and light intensity on the feeding behavior of three-spot juvenile seahorses, Hippocampus trimaculatus Leach. Aquaculture 2006, 256, 596–607. [Google Scholar] [CrossRef]
- Willadino, L.; Souza-Santos, L.P.; Mélo, R.C.S.; Brito, A.P.; Barros, N.C.S.; Araújo-Castro, C.M.V.; Galvão, D.B.; Gouveia, A.; Regis, C.G.; Cavalli, R.O. Ingestion rate, survival and growth of newly released seahorse Hippocampus reidi fed exclusively on cultured live food items. Aquaculture 2012, 360, 10–16. [Google Scholar] [CrossRef]
- Woods, C.M.C. Preliminary observations on breeding and rearing the seahorse Hippocampus abdominalis (Teleostei: Syngnathidae) in captivity. N. Z. J. Mar. Freshw. Res. 2000, 34, 475–485. [Google Scholar] [CrossRef]
- Zhang, D.; Yin, F.; Lin, J. Criteria for assessing juvenile quality of the lined seahorse, Hippocampus erectus. Aquaculture 2011, 322, 255–258. [Google Scholar] [CrossRef]
- Chang, M.; Southgate, P.C. Effects of varying dietary fatty acid composition on growth and survival of seahorse, Hippocampus sp., juveniles. Aquarium Sci. Conserv. 2001, 3, 205–214. [Google Scholar] [CrossRef]
- Filleul, M.A. Optimizing growth of juveniles big bellied seahorse Hippocampus abdominalis Lesson. Master’s Thesis, University of Tasmania, Hobart, Australia, 1996. [Google Scholar]
- Woods, C.M.C. Effects of varying Artemia enrichment on growth and survival of juvenile seahorses, Hippocampus abdominalis. Aquaculture 2003, 220, 537–548. [Google Scholar] [CrossRef]
- Sargent, J.R.; Bell, J.G.; Bell, M.V.; Henderson, R.J.; Tocher, D.R. Dietary origins and functions of long-chain (n-3) polyunsaturated fatty acids in marine fish. J. Mar. Biotechnol. 1995, 3, 26–28. [Google Scholar]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–118. [Google Scholar] [CrossRef]
- Corraze, G. Lipid nutrition. In Nutrition and Feeding of Fish and Crustaceans; Guillaume, J., Kaushik, S., Bergot, P., Metailler, R., Eds.; Springer-Praxis: Chichester, UK, 2001; pp. 111–129, ISBN 10: 1852332417; ISBN 13: 9781852332419. [Google Scholar]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The lipids. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2002; pp. 182–246. [Google Scholar]
- Sargent, J.R.; McEvoy, L.A.; Bell, J.G. Requirements, presentation and sources of polyunsaturated fatty acids in marine fish larval feeds. Aquaculture 1997, 155, 117–127. [Google Scholar] [CrossRef]
- Faleiro, F.; Narciso, L. Lipid dynamics during early development of Hippocampus guttulatus seahorses: Searching for clues on fatty acid requirements. Aquaculture 2010, 307, 56–64. [Google Scholar] [CrossRef]
- Watanabe, T.; Kiron, V. Prospects in larval fish dietetics. Aquaculture 1994, 124, 223–251. [Google Scholar] [CrossRef]
- Watanabe, T.; Arakawa, T.; Takeuchi, T.; Satoh, S. Comparison between eicosapentaenoic and docosahexaenoic acids in terms of essential fatty acid efficiency in juvenile striped jack Pseudocaranx dentex. Nippon. Suisan Gakk. 1989, 55, 1989–1995. [Google Scholar] [CrossRef]
- Faleiro, F.; Narciso, L. Prey–predator dynamics in seahorses (Hippocampus guttulatus): Deciphering fatty acid clues. Aquacult. Res. 2013, 44, 618–633. [Google Scholar] [CrossRef]
- Sargent, J.; Bell, G.; McEvoy, L.; Tocher, D.; Estevez, A. Recent developments in the essential fatty acid nutrition of fish. Aquaculture 1999, 177, 191–219. [Google Scholar] [CrossRef]
- Webster, C.D.; Lovell, R.T. Response of striped bass larvae fed brine shrimp from different sources containing different fatty acid compositions. Aquaculture 1990, 90, 49–62. [Google Scholar] [CrossRef]
- Ostrowski, A.C.; Laidley, C.W. Application of marine food fish techniques in marine ornamental aquaculture: Reproduction and larval first feeding. Aquar. Sci. Conserv. 2001, 3, 191–204. [Google Scholar] [CrossRef]
- Holt, G.J. Research on culturing the early life stages of marine ornamental fish. In Marine Ornamental Species—Collection, Culture & Conservation; Cato, J.C., Brown, C.L., Eds.; Iowa State Press: Ames, IA, USA, 2003; pp. 251–254. [Google Scholar] [CrossRef]
- Aragão, C.; Conceição, L.E.; Dinis, M.T.; Fyhn, H.J. Amino acid pools of rotifers and Artemia under different conditions: Nutritional implications for fish larvae. Aquaculture 2004, 234, 429–445. [Google Scholar] [CrossRef]
- Hamre, K.; Srivastava, A.; Rønnestad, I.; Mangor-Jensen, A.; Stoss, J. Several micronutrients in the rotifer Brachionus sp. may not fulfil the nutritional requirements of marine fish larvae. Aquacult. Nut. 2008, 14, 51–60. [Google Scholar] [CrossRef]
- Conceição, L.E.; Yúfera, M.; Makridis, P.; Morais, S.; Dinis, M.T. Live feeds for early stages of fish rearing. Aquacult. Res. 2010, 41, 613–640. [Google Scholar] [CrossRef]
- Zeng, C.; Shao, L.; Ricketts, A.; Moorhead, J. The importance of copepods as live feed for larval rearing of the green mandarin fish Synchiropus splendidus. Aquaculture 2018, 491, 65–71. [Google Scholar] [CrossRef]
- Alajmi, F.; Zeng, C.; Jerry, D.R. Domestication as a novel approach for improving the cultivation of calanoid copepods: A case study with Parvocalanus crassirostris. PLoS ONE 2015, 10, e0133269. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Divakaran, S.; Brown, C.L.; Ostrowski, A. Comparative digestive enzyme ontogeny in two marine larval fishes, Pacific threadfin (Polydactylus sexfilis) and bluefin trevally (Caranx melampygus). Fish Physiol. Biochem. 2001, 24, 225–241. [Google Scholar] [CrossRef]
- Pérez-Casanova, J.C.; Murray, H.M.; Gallant, J.W.; Ross, N.W.; Douglas, S.E.; Johnson, S.C. Development of the digestive capacity in larvae of haddock (Melanogrammus aeglefinus) and Atlantic cod (Gadus morhua). Aquaculture 2006, 251, 377–401. [Google Scholar] [CrossRef]
- Bolasina, S.; Pérez, A.; Yamashita, Y. Digestive enzymes activity during ontogenetic development and effect of starvation in Japanese flounder, Paralichthys olivaceus. Aquaculture 2006, 252, 503–515. [Google Scholar] [CrossRef]
- Furné, M.; García-Gallego, M.; Hidalgo, M.C.; Morales, A.E.; Domezain, A.; Domezain, J.; Sanz, A. Effect of starvation and refeeding on digestive enzyme activities in sturgeon (Acipenser naccarii) and trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. A 2008, 149, 420–425. [Google Scholar] [CrossRef]
- Lazo, J.P.; Mendoza, R.; Holt, G.J.; Aguilera, C.; Arnold, C.R. Characterization of digestive enzymes during larval development of red drum (Sciaenops ocellatus). Aquaculture 2007, 265, 194–205. [Google Scholar] [CrossRef]
- Palma, J.; Stockdale, J.; Correia, M.; Andrade, J.P. Growth and survival of adult long snout seahorse (Hippocampus guttulatus) using frozen diets. Aquaculture 2008, 278, 55–59. [Google Scholar] [CrossRef]
- Woods, C.M.C. Improving initial survival in cultured seahorses, Hippocampus abdominalis Leeson, 1827 (Teleostei: Syngnathidae). Aquaculture 2000, 190, 377–388. [Google Scholar] [CrossRef]
- Sorgeloos, P.; Lavens, P.; Léger, P.; Tackaert, W.; Versichele, D. Manual for the Culture of Brine Shrimp Artemia in Aquaculture; University of Ghent: Ghent, Belgium, 1986; ISBN 92-5-103934-8. [Google Scholar]
- Palma, J.; Andrade, J.P.; Bureau, D.P. The impact of dietary supplementation with astaxanthin on egg quality and growth of long snout seahorse (Hippocampus guttulatus) juveniles. Aquacult. Nut. 2017, 23, 304–312. [Google Scholar] [CrossRef]
- Lourie, S.A.; Pritchard, J.C.; Casey, S.P.; Truong, S.K.; Hall, H.J.; Vincent, A.C.J. The taxonomy of Vietnam’s exploited seahorses (family Syngnathidae). Biol. J. Linn. Soc. 1999, 66, 231–256. [Google Scholar] [CrossRef]
- Iwama, G.K.; Tautz, A.F. A simple growth model for salmonids in hatcheries. Can. J. Fish. Aquat. Sci. 1981, 38, 649–656. [Google Scholar] [CrossRef]
- Cho, C.Y. Fish nutrition, feeds and feeding: With special emphasis on salmonid aquaculture. Food Rev. Int. 1990, 6, 333–357. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis of AOAC International. In Agricultural Chemicals; Contaminants Drugs, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995; Volume 1. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Jorge, A.F.N. Determinação do Perfil de Ácidos Gordos do Músculo de Achigã (Micropterus salmoides). Master’s Thesis, Universidade Nova de Lisboa, Lisboa, Portugal, 2018. [Google Scholar]
- Palma, J.; Bureau, D.; Andrade, J.P. The effect of diet on ontogenic development of the digestive tract in juvenile reared long snout seahorse Hippocampus guttulatus. Fish Physiol. Biochem. 2014, 40, 739–775. [Google Scholar] [CrossRef]
- Schubert, P.; Vogt, L.; Eder, K.; Hauffe, T.; Wilke, T. Effects of Feed Species and HUFA Composition on Survival and Growth of the Longsnout Seahorse (Hippocampus reidi). Front. Mar. Sci. 2016, 3, 53. [Google Scholar] [CrossRef]
- Gardner, T. The Copepod/Artemia Trade-Off in the Captive Culture of Hippocampus Erectus, a Vulnerable Species in New York State. Master’s Thesis, Hofstra University, Long Island, NY, USA, 2004. [Google Scholar]
- Cohen, F.P.; Valenti, W.C. Opportunities and constraints for developing low-cost aquaculture of seahorses in mangrove estuaries. Aquaculture 2019, 502, 121–127. [Google Scholar] [CrossRef]
- Lall, S.P.; Lewis-McCrea, L.M. Role of nutrients in skeletal metabolism and pathology in fish—An overview. Aquaculture 2007, 267, 3–19. [Google Scholar] [CrossRef]
- Blanco, A.; Planas, M. Mouth growth and prey selection in juveniles of the European long-snouted seahorse, Hippocampus guttulatus. J. World Aquacult. Soc. 2015, 46, 596–607. [Google Scholar] [CrossRef]
- Blanco, A.; Planas, M.; Moyano, F.J. Ontogeny of digestive enzymatic capacities in juvenile seahorses Hippocampus guttulatus fed on different live diets. Aquacult. Res. 2016, 47, 3558–3569. [Google Scholar] [CrossRef]
- Pham, N.; Lin, J. The effects of different feed enrichments on survivorship and growth of early juvenile long-snout seahorse, Hippocampus reidi. J. World Aquacult. Soc. 2013, 44, 435–446. [Google Scholar] [CrossRef]
- Castro, A.L.D.C.; Diniz, A.D.F.; Martins, I.Z.; Vendel, A.L.; Oliveira, T.P.R.D.; Rosa, I.M.D.L. Assessing diet composition of seahorses in the wild using a non-destructive method: Hippocampus reidi (Teleostei: Syngnathidae) as a study-case. Neotrop. Ichthyol. 2008, 6, 637–644. [Google Scholar] [CrossRef]
- Tipton, K.; Bell, S. Foraging patterns of two syngnathid fishes: Importance of harpacticoid copepods. Mar. Ecol. Prog. Ser. 1988, 47, 31–113. [Google Scholar] [CrossRef]
- Bodin, N.; Delfosse, G.; Thi, T.; Thu, N.; Le, E.; Abboudi, T.; Larondelle, Y.; Rollin, X. Effects of fish size and diet adaptation on growth performances and nitrogen utilization of rainbow trout (Oncorhynchus mykiss W.) juveniles given diets based on free and/or protein-bound amino acids. Aquaculture 2012, 356, 105–115. [Google Scholar] [CrossRef]
- Randazzo, B.; Rolla, L.; Ofelio, C.; Planas, M.; Gioacchini, G.; Vargas, A.; Giorgini, E.; Olivotto, I. The influence of diet on the early development of two seahorse species (H. guttulatus and H. reidi): Traditional and innovative approaches. Aquaculture 2018, 490, 75–90. [Google Scholar] [CrossRef]
- Planas, M.; Quintas, P.; Chamorro, A.; Balcázar, J.L. Husbandry and rearing of the seahorse Hippocampus guttulatus (project Hippocampus). In Proceedings of the World Aquaculture Society, World Aquaculture 2009, Veracruz, México, 25–29 September 2009; p. 668. [Google Scholar]
- Planas, M.; Blanco, A.; Chamorro, A.; Valladares, S.; Pintado, J. Temperature-induced changes of growth and survival in the early development of the seahorse Hippocampus guttulatus. J. Exp. Mar. Biol. Ecol. 2012, 438, 154–162. [Google Scholar] [CrossRef]
- Manning, C.G.; Foster, S.J.; Vincent, A.C.J. A review of the diets and feeding behaviours of a family of biologically diverse marine fishes (Family Syngnathidae). Rev. Fish Biol. Fish. 2019, 29, 197–221. [Google Scholar] [CrossRef]
- Rønnestad, I.; Yúfera, M.; Ueberschär, B.; Ribeiro, L.; Sæle, Ø.; Boglione, C. Feeding behaviour and digestive physiology in larval fish: Current knowledge, and gaps and bottlenecks in research. Rev. Aquac. 2013, 5, S59–S98. [Google Scholar] [CrossRef]
- Zambonino-Infante, J.L.; Cahu, C.L. Ontogeny of the gastrointestinal tract of marine fish larvae. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Celino, F.T.; Hilomen-Garcia, G.V.; del Norte-Campos, A.G. Feeding selectivity of the seahorse, Hippocampus kuda (Bleeker), juveniles under laboratory conditions. Aquacult. Res. 2012, 43, 1804–1815. [Google Scholar] [CrossRef]
- Zhang, D.; Lin, T.; Liu, X. A comparison of growth, survival, and fatty acid composition of the lined seahorse, Hippocampus erectus, juveniles fed enriched Artemia and a calanoid copepod, Schmackeria dubia. J. World Aquacult. Soc. 2015, 46, 608–616. [Google Scholar] [CrossRef]


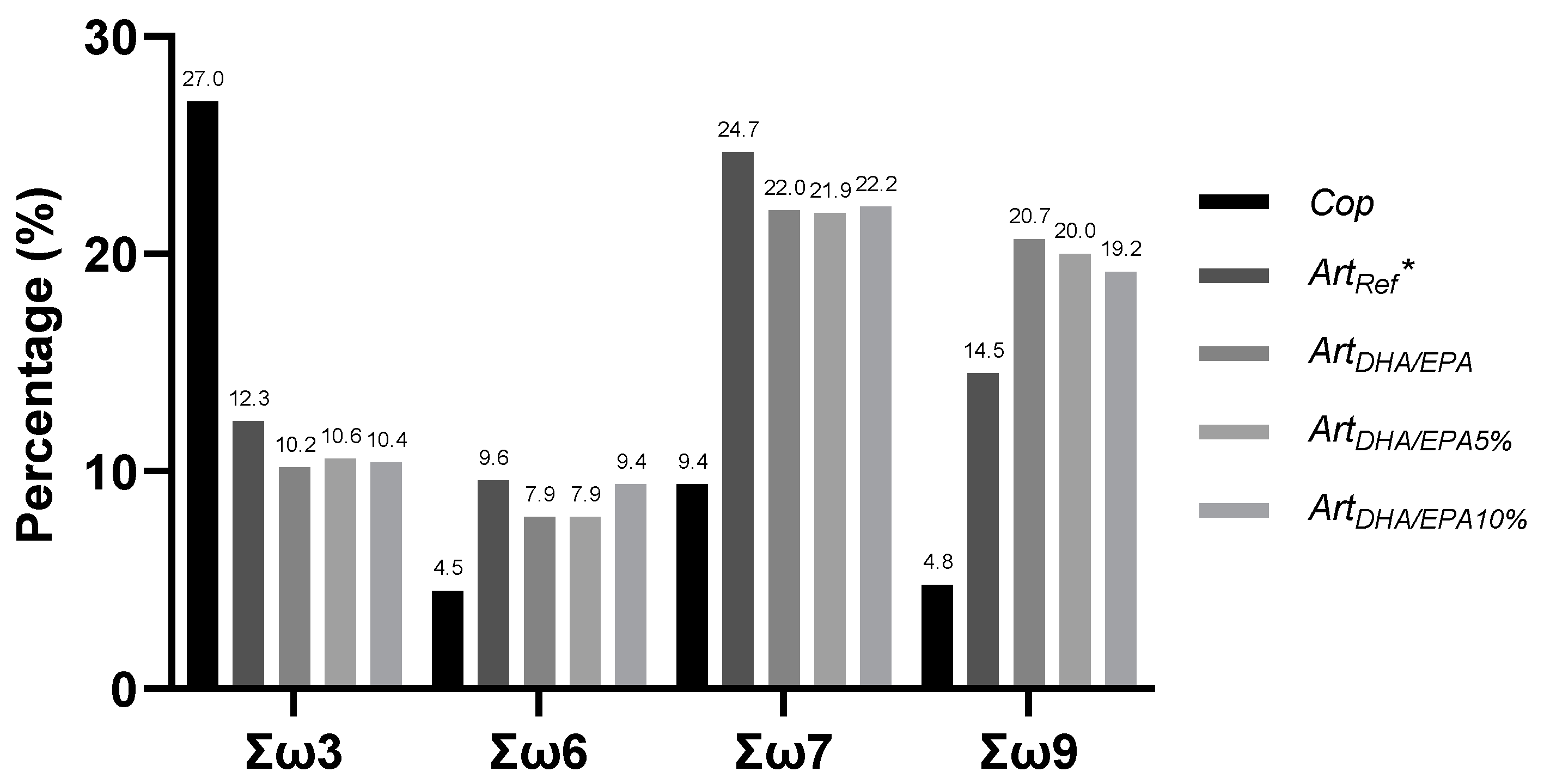{kind=link}
| Proximate Composition | Cop | ArtRef * | ArtDHA/EPA | ArtDHA/EPA5% | ArtDHA/EPA10% |
|---|---|---|---|---|---|
| Moisture (%) | 86.9 ± 1.1 a | 70.1 ± 0.7 b | 70.1 ± 0.6 b | 68.5 ± 0.8 b | 66.9 ± 0.8 b |
| Dry Matter (%) | 13.1 ± 0.9 b | 29.9 ± 1.1 a | 29.9 ± 1 a | 31.4 ± 1.3 a | 33.2 ± 1.1 a |
| Gross Protein (g/100 g DM) | 10.9 ± 0.7 c | 16.5 ± 0.6 b | 17.9 ± 0.8 ab | 19.1 ± 0.9 ab | 20 ± 1.1 a |
| Total Lipids (g/100 g DM) | 4.9 ± 0.9 b | 11.8 ± 0.7 a | 10.2 ± 0.7 a | 10 ± 0.8 a | 10.5 ± 0.9 a |
| Gross Energy (Kcal/100 DM) | 503.85 ± 1.1 a | 497.8 ± 0.6 ab | 489.8 ± 0.7 c | 495.6 ± 0.7 ab | 492.2 ± 1.2 b |
| Fatty Acid | Isochrysis galbana | Nannochloropsis sp. |
|---|---|---|
| C18:2ω6 (LA) | 7.5 | 4 |
| C18:3ω3 (ALA) | 15.5 | 0.2 |
| C20:4ω6 (ARA) | 0.2 | 5.3 |
| C20:5ω3 (EPA) | 0.1 | 36.1 |
| C22:6ω3 (DHA) | 16.5 | - |
| ΣSFA | 25.5 | 26.2 |
| ΣMUFA | 20.2 | 22.9 |
| ΣPUFA | 50.1 | 48.1 |
| ΣHUFA | 27.3 | 42.3 |
| ΣEFA | 39.8 | 45.6 |
| Σω3 | 39.8 | 37.2 |
| Σω6 | 10.6 | 10.9 |
| Σω9 | 15.2 | 3.2 |
| DHA/EPA | 165 | 0 |
| ω3/ω6 | 3.75 | 3.41 |
| PUFA/SFA | 1.96 | 1.84 |
| Fatty Acid | Cop | ArtRef * | ArtDHA/EPA | ArtDHA/EPA5% | ArtDHA/EPA10% |
|---|---|---|---|---|---|
| C12:0 | 0.19 ± 0.01 b | 0.16 ± 0.02 c | 0.15 ± 0.02 c | 0.18 ± 0.01 b | 0.23 ± 0.07 a |
| C14:0 | 10.25 ± 0.19 a | 3.06 ± 0.19 c | 4.26 ± 0.16 b | 4.25 ± 0.2 b | 4.24 ± 0.15 b |
| C16:0 | 31.23 ± 0.4 a | 15.69 ± 0.41 b | 15.94 ± 0.88 b | 16.91 ± 1.15 b | 17.07 ± 0.47 b |
| C18:0 | 6.13 ± 0.28 a | 4.84 ± 0.3 b | 5.35 ± 0.2 b | 5.19 ± 0.14 b | 5.07 ± 0.29 b |
| C20:0 | 0.11 ± 0 a | 0.11 ± 0.03 b | 0.14 ± 0 b | 0.14 ± 0.02 a | 0.09 ± 0.03 b |
| ΣSFA | 47.9 a | 23.87 c | 25.84 c | 26.65 c | 36.7 b |
| C14:1ω5 | - | 0.33 ± 0.02 a | 0.23 ± 0.01 b | 0.21 ± 0 b | 0.23 ± 0.02 b |
| C16:1ω7c | 8.88 ± 0.2 b | 13.79 ± 0.67 a | 12.74 ± 0.46 a | 13.06 ± 0.61 a | 13.58 ± 0.47 a |
| C16:1ω7t | 0.27 ± 0.07 b | 0.47 ± 0.09 a | 0.40 ± 0.04 a | 0.36 ± 0.02 a | 0.39 ± 0.02 a |
| C18:1ω9c | 2.91 ± 0.04 c | 6.07 ± 6.64 b | 12.36 ± 0.92 a | 12.21 ± 1.12 a | 11.82 ± 1.64 a |
| C18:1ω9t | 1.9 ± 0.09 b | 8.08 ± 1.8 a | 8.3 ± 1.3 a | 7.73 ± 1.44 a | 7.43 ± 1.29 a |
| C18:1ω7 | 0.29 ± 0.42 a | 0.12 ± 0.06 b | 0.10 ± 0.01 b | 0.09 ± 0.07 bc | 0.07 ± 0.02 c |
| C20:1ω9c | - | 0.34 ± 0.08 a | 0.33 ± 0.03 a | 0.33 ± 0.01 a | 0.32 ± 0.02 a |
| ΣMUFA | 14.26 c | 29.2 b | 34.46 a | 33.99 a | 33.82 a |
| C16:2ω4 | 0.27 ± 0.1d | 1.04 ± 0.05 a | 0.73 ± 0.45 b | 0.71 ± 0.38 b | 0.38 ± 0.22 c |
| C18:2ω6 (LA) | 2.09 ± 0.04 b | 5.05 ± 0.27 a | 4.56 ± 0.02 a | 4.49 ± 0.06 a | 4.52 ± 0.13 a |
| C18:2ω4 | - | 0.72 ± 0.02 a | 0.48 ± 0.03 b | 0.46 ± 0.02 b | 0.39 ± 0.08 b |
| C18:3ω6 | 0.34 ± 0.02d | 0.83 ± 0.13 b | 0.63 ± 0.09 c | 0.60 ± 0.06 c | 1.56 ± 1.6 a |
| C18:3ω3 (ALA) | 2.9 ± 0.02 a | 3.23 ± 0.22 a | 3.18 ± 0.1 a | 3.07 ± 0.08 a | 2.93 ± 0.18 a |
| C18:4ω3 | 1.5 ± 0.09 a | 0.56 ± 0.04 c | 1.14 ± 0.04 a | 1.07 ± 0.03 a | 0.85 ± 0.06 b |
| C20:3ω6 | 0.23 ± 0.02 a | 0.24 ± 0.08 a | 0.16 ± 0.05 a | 0.17 ± 0.04 a | 0.17 ± 0.08 a |
| C20:4ω6 (ARA) | 1.28 ± 0.1 c | 3.44 ± 0.17 a | 2.55 ± 0.02 b | 2.62 ± 0.01 b | 3.14 ± 0.57 a |
| C20:4ω3 | 0.25 ± 0.08 a | 0.24 ± 0.06 a | 0.11 ± 0.05 b | 0.14 ± 0.05 b | 0.08 ± 0.04 c |
| C20:5ω3 (EPA) | 10.48 ± 1.46 a | 8.26 ± 0.3 b | 5.58 ± 0.3 c | 5.86 ± 0.46 c | 5.94 ± 0.89 c |
| C22:6ω3 (DHA) | 11.67 ± 0.79 a | - | 0.23 ± 0.04 c | 0.47 ± 0.09 b | 0.58 ± 0.05 b |
| ΣPUFA | 31.01 a | 23.6 b | 19.35 c | 19.63 c | 20.78 c |
| ΣHUFA | 25.18 a | 12.51 b | 9.61 c | 10.13 c | 10.58 c |
| ΣEFA | 28.42 a | 19.98 b | 16.1 c | 16.51 c | 17.11 c |
| Σ Others | 6.83d | 23.33 a | 20.35 b | 19.73 b | 17.7 c |
| Fatty Acid Ratios | Cop | ArtRef * | ArtDHA/EPA | ArtDHA/EPA5% | ArtDHA/EPA10% |
|---|---|---|---|---|---|
| DHA/EPA | 1.11 | 0 | 0.04 | 0.08 | 0.1 |
| ω3/ω6 | 8 | 1.29 | 1.29 | 1.34 | 1.1 |
| PUFA/EFA | 0.65 | 0.99 | 0.75 | 0.74 | 0.78 |
| Cop | ArtDHA/EPA | ArtDHA/EPA5% | ArtDHA/EPA10% | |
|---|---|---|---|---|
| Survival | 72 | 88 | 87 | 77 |
| ML (cm) 0 DPP | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 |
| ML (cm) 28 DPP | 4.4 ± 0.3 a | 3.2 ± 0.4 c | 3.8 ± 0.3 b | 3.7 ± 0.3 b |
| MW (mg) 0 DPP | 2.4 ± 0.4 | 2.4 ± 0.4 | 2.4 ± 0.4 | 2.4 ± 0.4 |
| MW (mg) 28 DPP | 146 ± 39.6 a | 53.1 ± 19.3 c | 89 ± 22.7 b | 88.5 ± 28.4 b |
| LG (cm day−1) | 0.12 ± 0.003 a | 0.07 ± 0.004 c | 0.09 ± 0.004 b | 0.09 ± 0.004 b |
| WG (mg day−1) | 5.14 ± 0.33 a | 1.83 ± 0.33 c | 3.09 ± 0.82 b | 3.07 ± 0.23 b |
| TGC | 0.008 a | 0.003 c | 0.005 b | 0.005 b |
| CF | 0.17 a | 0.17 a | 0.17 a | 0.17 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palma, J.; Lima, R.; Andrade, J.P.; Lança, M.J. Optimization of Live Prey Enrichment Media for Rearing Juvenile Short-Snouted Seahorse, Hippocampus hippocampus. Fishes 2023, 8, 494. https://doi.org/10.3390/fishes8100494
Palma J, Lima R, Andrade JP, Lança MJ. Optimization of Live Prey Enrichment Media for Rearing Juvenile Short-Snouted Seahorse, Hippocampus hippocampus. Fishes. 2023; 8(10):494. https://doi.org/10.3390/fishes8100494
Chicago/Turabian StylePalma, Jorge, Ricardo Lima, José Pedro Andrade, and Maria João Lança. 2023. "Optimization of Live Prey Enrichment Media for Rearing Juvenile Short-Snouted Seahorse, Hippocampus hippocampus" Fishes 8, no. 10: 494. https://doi.org/10.3390/fishes8100494
APA StylePalma, J., Lima, R., Andrade, J. P., & Lança, M. J. (2023). Optimization of Live Prey Enrichment Media for Rearing Juvenile Short-Snouted Seahorse, Hippocampus hippocampus. Fishes, 8(10), 494. https://doi.org/10.3390/fishes8100494