

Effects of Myo-Inositol on the Growth Performance, Digestive Enzyme Activity, and Antioxidation of Juvenile Hucho taimen

 ,
, 
Abstract
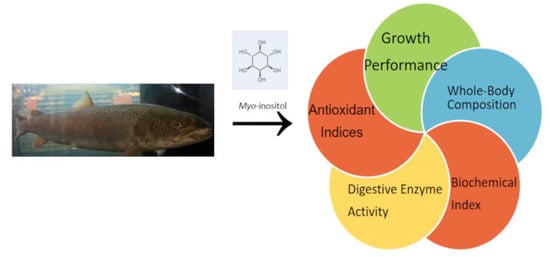
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Feeding Management
- Weight gain (WG, %) = (Wf − Wi)/Wi × 100;
- Specific growth rate (SGR, %/day) = (ln Wf − ln Wi)/days) × 100;
- Feed conversion ratio (FCR) = Feed consumed (g)/(Wf − Wi);
- Condition factor (CF, g·cm−3) = Wf × 100/Lf3;
- Viscerosomatic index (VSI, %) = (Wv/Wf) × 100;
- Hepatosmatic index (HSI, %) = (Wh/Wf) × 100.
2.2. Sample Collection
2.3. Whole-Body Composition Analyses
2.4. Biochemical Analysis
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Whole-Body Composition
3.3. Digestive Enzyme Activity
3.4. Biochemical Index
3.5. Antioxidant Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Liu, Y.; Zhao, X.; Wang, C.; Lu, S.; Liu, H.; Xu, Q. Effects of Aloe Powder on growth performance and serum indices of Taimen Hucho taimen. Chin. J. Fish. 2020, 33, 1–6. [Google Scholar]
- Wu, Q.; Wang, S.; Zhen, Y.; Wang, T.; Zhang, X.; Sun, Z. Research progress of preparation process and application of inositol. Heilongjiang Anim. Sci. Vet. Med. 2023, 20, 41–46. [Google Scholar]
- Li, S.; Jiang, W.; Feng, L.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.; Tang, L.; Tang, W.; Zhang, Y.; et al. Dietary myo-inositol deficiency decreased the growth performances and impaired intestinal physical barrier function partly relating to nrf2, jnk, e2f4 and mlck signaling in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2017, 67, 475–492. [Google Scholar] [CrossRef] [PubMed]
- Bu, X.; Lin, Z.; Liu, S.; Wang, C.; Wang, N.; Lei, Y.; Zhu, J.; Wang, X.; Qin, J.; Chen, L. Effects of myo -inositol on growth performance, body composition, antioxidant status, non-specific immunity and lipid metabolism of juvenile Chinese mitten crab (Eriocheir sinensis). Aquac. Nutr. 2020, 26, 1623–1635. [Google Scholar] [CrossRef]
- Li, R.; Kou, S.; Yang, Q.; Tan, B.; Chi, S.; Yi, Y.; Zhu, D. Effects of Myo-Inositol on Growth, Non-Specific Immunity and Intestinal Microflora Composition of Juvenile Litopenaeus Vannamei. Chin. J. Anim. Nutr. 2023, 35, 1182–1194. [Google Scholar]
- Feng, L.; Li, J.; Zhou, X.; Liu, Y. Effects of inoistol deficiency on digestive and immune function of juvenile jian carp(Cyprinus carpio var. Jian). Chin. J. Anim. Nutr. 2009, 21, 878–883. [Google Scholar]
- Jiang, W.; Liu, Y.; Feng, L.; Zhou, X. Histopathological observation of juvenile Cyprinus carpio var. Jian with myo-inositol deficiency and determinations of antioxidative ability and intestinal microbial population. Vet. Sci. China 2009, 31, 64–69. [Google Scholar]
- Cui, H.; Liu, B.; Ge, X.; Xie, J. Effects of dietary inositol on growth performance, physiological and biochemical indexes of serum and fat content in liver and muscle of juvenile Wuchang bream (Megalobrama amblycephala). J. Shanghai Ocean Univ. 2013, 22, 868–875. [Google Scholar]
- Deng, D.; Hemre, G.; Wilson, R.P. Juvenile sunshine bass (Morone chrysops × Morone saxatilis) do not require dietary myo-inositol. Aquaculture 2002, 213, 387–393. [Google Scholar] [CrossRef]
- Wen, H.; Zhao, Z.; Jiang, M.; Liu, A.; Wu, F.; Liu, W. Dietary myo-inositol requirement for grass carp, Ctenopharyngodon Idella Fingerling. J. Fish. Sci. China 2007, 14, 794–800. [Google Scholar]
- Khosravi, S.; Lim, S.; Rahimnejad, S.; Kim, S.; Lee, B.; Kim, K.; Han, H.; Lee, K. Dietary myo-inositol requirement of parrot fish. Oplegnathus fasciatus. Aquaculture 2015, 436, 1–7. [Google Scholar] [CrossRef]
- Lee, B.; Lee, K.; Lim, S.; Lee, S. Dietary myo-inositol requirement for olive flounder, Paralichthys olivaceus (Temminch et Schlegel). Aquac. Res. 2009, 40, 83–90. [Google Scholar] [CrossRef]
- Lin, K.; Feng, J.; Yang, H.; Chen, Y.; Chen, X.; Luo, H.; Huang, W.; Wang, H.; Luo, L. Effects of inositol supplementation to practical dietary on growth performance, lipid metabolism and antioxidant activity of Cteno pharyngodon Idella. J. Fish. China 2018, 42, 1428–1437. [Google Scholar]
- Wu, H.; Tang, Z.; Yang, H.; Luo, Y.; Huang, K.; Gan, X. Effects of dietary inositol on growth, liver and muscle fat content and biochemical indices of blood serum in tilapia (Oreoc hromis niloticus × Oreochromis aureus). J. South. Agric. 2011, 42, 1415–1419. [Google Scholar]
- Kanazawa, A.; Teshima, S.; Tanaka, N. Nutritional requirements of prawn V: Requirements for choline and inositol. Mem. Fac. Fish. Kagoshima Univ. 1976, 25, 47–51. [Google Scholar]
- Shiau, S.; Su, S. Dietary inositol requirement for juvenile grass shrimp, Penaeus Mondon. Aquac. 2004, 241, 1–8. [Google Scholar] [CrossRef]
- Liu, T.; Li, A.; Zhang, J. Studies on vitamin nutrition for the shrimp penaeus chinensis-X: Studies on the choline chloride and inositol requirements in the shrimp Penaeus chinensis. J. Ocean Univ. Qingdao 1993, 23, 71–78. [Google Scholar]
- Bu, X.; Zhu, J.; Liu, S.; Wang, C.; Xiao, S.; Lu, M.; Li, E.; Wang, X.; Qin, J.G.; Chen, L. Growth, osmotic response and transcriptome response of the euryhaline teleost, Oreochromis mossambicus fed different myo-inositol levels under long-term salinity stress. Aquaculture 2021, 534, 736294. [Google Scholar] [CrossRef]
- Chen, S.; Guo, Y.; Espe, M.; Yang, F.; Fang, W.; Wan, M.; Niu, J.; Liu, Y.; Tian, L. Growth performance, haematological parameters, antioxidant status and salinity stress tolerance of juvenile Pacific white shrimp (Litopenaeus vannamei) fed different levels of dietary myo-inositol. Aquac. Nutr. 2018, 24, 1527–1539. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhao, F.; Cairang, Z.; Zhou, Z.; Du, Q.; Wang, J.; Zhao, F.; Wang, Q.; Li, Z.; Zhang, X. Role of Dietary Tea Polyphenols on Growth Performance and Gut Health Benefits in Juvenile Hybrid Sturgeon (Acipenser baerii ♀ × A. schrenckii ♂). Fish Shellfish Immunol. 2023, 139, 108911–108921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, Q.; Li, M.G.; Li, F.; Qin, M.; Xie, Y.; Xu, J.; Liu, Y.; Tong, T. The Effects of Dietary Fermented Soybean Meal Supplementation on the Growth, Antioxidation, Immunity, and mTOR Signaling Pathway of Juvenile Coho Salmon (Oncorhynchus kisutch). Fishes 2023, 8, 448. [Google Scholar] [CrossRef]
- Yuan, H.; Hu, N.; Zheng, Y.; Hou, C.; Tan, B.; Shi, L.; Zhang, S. A Comparison of Three Protein Sources Used in Medium-Sized Litopenaeus vannamei: Effects on Growth, Immunity, Intestinal Digestive Enzyme Activity, and Microbiota Structure. Fishes 2023, 8, 449. [Google Scholar] [CrossRef]
- Chu, J.-H.; Weng, T.-S.; Huang, T.-W. The Effects of Replacing Fish Meal Protein with Black Soldier Fly Meal and Sodium Butyrate Supplementation on the Growth Performance, Lipid Peroxidation, and Intestinal Villi Status of Jade Perch, Scortum Barcoo Fingerlings. Fishes 2023, 8, 437. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, P.; Kong, Q.; Deng, Y.; Zhang, W.; Xu, G.; Tang, H. Effects of Co-Fermented Feed Using Lactobacillus acidophilus, Limosilactobacillus reuteri and Lactiplantibacillus plantarum on Growth, Antioxidant Capacity, Fatty Acids and Gut Microbiota of Largemouth Bass (Micropterus salmoides). Fishes 2023, 8, 433–450. [Google Scholar] [CrossRef]
- Aoac, I. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Maryland, MD, USA, 2010. [Google Scholar]
- Yin, G.; Zhen, Y.; Wang, T.; Zhang, X.; Sun, Z. The Mechanism of Inositol and Its Application in Animal Production. J. Econ. Anim. 2023, 11, 1–7. [Google Scholar]
- Levine, A.; Billington, C. Why do we eat? A neural systems approach. Annu. Rev. Nutr. 1997, 17, 597–619. [Google Scholar] [CrossRef]
- Chen, S.; Ji, W.; Lv, Y.; Chang, Q.; Pan, S. Nutrient function of inositol on the growth of black sea bream, juvenile, Sparus Marcocephalus. J. Mar. Sci. 1995, 5, 13–15. [Google Scholar]
- Burtle, G.; Lovell, R. Lack of response of channel catfish (Lctalurus punctatus) to dietary myo-inositol. Can. J. Fish. Aquat. Sci. 2011, 46, 218–222. [Google Scholar] [CrossRef]
- Waagbø, S.; Lie, R. Effects of inositol supplementation on growth, chemical composition and blood chemistry in Atlantic salmon (Salmo salar L.)fry. Aquac. Nutr. 1998, 4, 53–59. [Google Scholar] [CrossRef]
- Mai, K.; Wu, G.; Zhu, W. Abalone, Haloiotis discus hannai Ino, can synthesize myo-inositol de novo to meet physiological needs. Nuturition 2001, 131, 2898–2903. [Google Scholar]
- Chen, X.; Wang, J.; Zhao, W. Effects of dietary inositol supplementation on growth, digestive performance, antioxidant capacity, and body composition of golden pompano(Trachinotus ovatus). Front. Physiol. 2022, 13, 850470. [Google Scholar] [CrossRef] [PubMed]
- Peres, H.; Lim, C.; Klesius, P. Growth, chemical composition and resistance to Streptococcus iniae challenge of juvenile nile tilapia (Oreochromis niloticus) fed graded levels of dietary inositol. Aquaculture 2004, 235, 423–430. [Google Scholar] [CrossRef]
- Wang, Y.; Li, B.; Wang, J.; Wang, C.; Wang, X.; Ma, C.; Wang, S.; Sun, Y.; Hao, T. Dietary Myo-inositol Requirement of Juvenile Turbot(Scophthalmus maximus L.). Chin. J. Anim. Nutr. 2019, 31, 5122–5132. [Google Scholar]
- Jiang, W.; Feng, L.; Liu, Y.; Jiang, J.; Zhou, X. Growth, digestive capacity and intestinal microflora of juvenile Jian carp (Cyprinus carpio var. Jian) fed graded levels of dietary inositol. Aquac. Res. 2010, 40, 955–962. [Google Scholar] [CrossRef]
- Yu, Q.; Han, F.; Huang, M.; Wang, X.; Qin, J.G.; Chen, L.; Li, E. The effects of dietary myo-inositol on growth and physiological, biochemical, and molecular responses in the Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2023, 568, 739323. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, C.; Lu, S.; Liu, S.; Wang, Y.; Liu, H.; Yang, Y.; Xu, Q. Effects of dietary myo-Inositol on digestive physiology of Acipenser Schrenckii. Feed Ind. 2022, 43, 53–59. [Google Scholar]
- Yone, Y.; Furuichi, M.; Shitanda, K. Vitamin requirement of the red sea bream: Relationship between inositol requirements and glucose levels in diet. Bull. Jpn. Soc. Sci. Fish. 1971, 37, 149–155. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, X. Dietary glutamine supplementation improves structure and function of intestine of juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2006, 256, 389–394. [Google Scholar]
- Fang, Z.; Wang, C.; Wei, H. Effects of mercury and selenium on Na+,K+-ATPase activity in xiphophorus heller heckle. Chin. J. Appl. Environ. Biol. 2006, 12, 220–223. [Google Scholar]
- Xu, Q.; Wang, C.; Xu, H.; Zhu, Q.; Yin, J. Effects of glutamine dipeptide on intestine antioxidant, abilities of digestion and absorption of Hucho taimen larvae. J. Fish. Sci. China 2010, 17, 351–356. [Google Scholar]
- Ye, J.; Han, Y.; Zhao, J.; Lu, T.; Liu, H.; Yang, Y. Effects of dietary olaquindox on antioxidant enzymes system in hepatopancreas of Cyprinus Carpio. J. Fish. China 2004, 28, 231–235. [Google Scholar]
- Liu, Z.; Song, W.; Yu, B. Effects of inositol on tissue lipid and protein oxidation and antioxidant of crucian. China Feed 2019, 12, 39–43. [Google Scholar]
- Hu, K.; Li, S.; Feng, L.; Jiang, W.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Zhou, X. Protective effect of myo-inositol on oxidative damage of head kidney and spleen in juvenile grass carp (Ctenopharyngodon idella) induced by Aeromonas hydrophila. J. Fish. China 2019, 43, 2256–2267. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content |
|---|---|
| Casein | 34.00 |
| Fish meal | 23.00 |
| Gelatin | 10.00 |
| Dextrin | 15.30 |
| Fish oil | 11.00 |
| Phospholipids | 2.00 |
| Ca(H2PO4)2 | 1.00 |
| CMC | 2.60 |
| Vitamin premix 1 | 0.30 |
| Mineral premix 2 | 0.20 |
| Additive 3 | 0.60 |
| Total | 100.00 |
| Nutrient levels | |
| Crude protein | 52.3 |
| Crude lipid | 12.4 |
| MI (mg/kg) | 128 |
| Treatments | Initial Weight (g) | Final Weigh (g) | WG (%) | SGR (%) | FCR | CF (g·cm−3) | HSI (%) | VSI (%) |
|---|---|---|---|---|---|---|---|---|
| G1 | 2.45 ± 0.27 | 4.17 ± 0.76 | 70.20 ± 1.91 a | 0.94 ± 0.11 a | 1.98 ± 0.20 bc | 0.60 ± 0.10 b | 1.03 ± 0.18 | 7.11 ± 0.33 a |
| G2 | 2.55 ± 0.14 | 4.53 ± 1.02 | 77.65 ± 1.00 b | 1.03 ± 0.14 ab | 1.94 ± 0.14 ab | 0.62 ± 0.03 bc | 1.12 ± 0.28 | 7.34 ± 0.98 ab |
| G3 | 2.75 ± 0.20 | 5.02 ± 0.68 | 82.55 ± 2.99 bc | 1.11± 0.21 b | 1.89 ± 0.09 a | 0.58 ± 0.08 a | 1.61 ± 0.08 | 7.88± 0.70 ab |
| G4 | 2.55 ± 0.19 | 4.74 ± 0.41 | 86.00 ± 2.16 c | 1.07 ± 0.12 ab | 2.00 ± 0.07 bc | 0.62 ± 0.08 bc | 1.43 ± 0.15 | 9.88 ± 1.00 b |
| G5 | 2.54± 0.21 | 4.58± 1.27 | 80.31 ± 1.40 b | 1.05 ± 0.18 ab | 2.06 ± 0.15 c | 0.61 ± 0.07 b | 1.36 ± 0.04 | 7.59 ± 0.32 ab |
| G6 | 2.91± 0.38 | 5.14± 1.25 | 76.63 ± 3.51 b | 1.02 ± 0.09 ab | 2.06 ± 0.04 c | 0.63 ± 0.05 bc | 1.53 ± 0.13 | 7.43 ± 1.11 ab |
| Treatments | Moisture (%) | Crude Protein (%) | Crude Lipid (%) | Ash (%) |
|---|---|---|---|---|
| G1 | 80.19 ± 0.84 | 14.97 ± 1.04 | 2.10 ± 0.02 a | 2.21 ± 0.08 a |
| G2 | 80.74 ± 0.84 | 13.53 ± 1.10 | 2.16 ± 0.18 ab | 2.29 ± 0.16 a |
| G3 | 80.32 ± 0.40 | 13.70 ± 1.42 | 2.41 ± 0.19 b | 2.55 ± 0.51 b |
| G4 | 79.90 ± 0.12 | 14.38 ± 0.55 | 2.26 ± 0.02 ab | 2.33 ± 0.28 a |
| G5 | 80.30 ± 1.98 | 13.12 ± 0.75 | 2.28 ± 0.01 ab | 2.80 ± 0.24 c |
| G6 | 80.49 ± 0.25 | 14.13 ± 0.11 | 2.32 ± 0.03 ab | 2.22 ± 0.36 a |
| Tissue | Enzyme | G1 | G2 | G3 | G4 | G5 | G6 |
|---|---|---|---|---|---|---|---|
| Liver | Amylase | 30.37 ± 1.76 | 30.67 ± 4.21 | 32.29 ± 0.68 | 34.85 ± 2.89 | 33.38 ± 3.06 | 30.82 ± 1.77 |
| Protease | 24.60 ± 2.04 | 30.30 ± 5.54 | 31.55 ± 3.70 | 28.95 ± 1.26 | 27.96 ± 5.24 | 30.31 ± 4.61 | |
| Lipase | 8.54 ± 1.12 b | 6.44 ± 0.19 a | 7.61 ± 0.41 ab | 8.14 ± 0.82 b | 7.40 ± 0.38 ab | 8.02 ± 1.52 b | |
| Pyloric caeca | Amylase | 195.98 ± 12.01 a | 205.04 ± 8.67 ab | 236.07 ± 21.91 c | 222.71 ± 5.81 bc | 224.36 ± 7.13 bc | 218.60 ± 15.49 abc |
| Protease | 98.81 ± 9.59 | 105.28 ± 10.13 | 118.55 ± 14.21 | 120.66 ± 19.28 | 108.24 ± 8.38 | 113.07 ± 4.35 | |
| Lipase | 41.97 ± 0.42 a | 42.28 ± 1.92 a | 42.46 ± 1.32 a | 47.46 ± 2.91 b | 44.80 ± 4.75 ab | 41.94 ± 1.29 a | |
| Intestine | Amylase | 161.64 ± 4.44 | 166.58 ± 11.02 | 176.06 ± 14.47 | 164.21 ± 13.04 | 160.95 ± 6.76 | 160.07 ± 4.75 |
| Protease | 43.66 ± 5.39 a | 44.98 ± 5.02 a | 50.87 ± 8.44 ab | 56.66 ± 2.09 b | 56.21 ± 1.21 b | 49.83 ± 3.24 ab | |
| Lipase | 34.59 ± 6.51 | 34.28 ± 3.50 | 33.38 ± 1.66 | 30.26 ± 2.70 | 29.61 ± 0.94 | 33.18 ± 3.03 |
| Tissue | Biochemical Index | G1 | G2 | G3 | G4 | G5 | G6 |
|---|---|---|---|---|---|---|---|
| Pyloric caeca | AKP | 55.89 ± 4.78 | 56.60 ± 6.77 | 69.45 ± 9.50 | 65.55 ± 8.57 | 60.48 ± 2.91 | 61.63 ± 6.93 |
| Na+,K+-ATPase (μmolpi/gprot/h) | 1.32 ± 0.23 | 1.23 ± 0.05 | 1.25 ± 0.16 | 1.39 ± 0.30 | 1.21 ± 0.12 | 1.18 ± 0.27 | |
| Intestine | AKP | 42.16 ± 3.11 a | 49.58 ± 4.68 ab | 74.81 ± 11.62 c | 59.31 ± 8.73 b | 59.95 ± 8.06 b | 47.23 ± 8.55 ab |
| Na+,K+-ATPase (μmolpi/gprot/h) | 0.54 ± 0.09 a | 0.79 ± 0.05 a | 0.74 ± 0.05 a | 1.28 ± 0.27 b | 0.84 ± 0.05 a | 0.74 ± 0.12 a |
| Treatments | SOD (NU·mgprot−1) | MDA(nmol·mgprot−1) |
|---|---|---|
| G1 | 68.97 ± 8.31 a | 9.67 ± 0.42 b |
| G2 | 81.57 ± 12.81 ab | 6.20 ± 0.77 a |
| G3 | 82.26 ± 8.77 ab | 6.17 ± 1.57 a |
| G4 | 82.89 ± 7.70 ab | 7.32 ± 1.78 a |
| G5 | 89.59 ± 13.62 b | 7.47 ± 0.84 a |
| G6 | 76.32 ± 6.66 ab | 7.37 ± 0.81 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Yang, X.; Wang, C.; Shang, B.; Zhao, F.; Xu, H.; Xu, Q. Effects of Myo-Inositol on the Growth Performance, Digestive Enzyme Activity, and Antioxidation of Juvenile Hucho taimen. Fishes 2023, 8, 567. https://doi.org/10.3390/fishes8120567
Zhang M, Yang X, Wang C, Shang B, Zhao F, Xu H, Xu Q. Effects of Myo-Inositol on the Growth Performance, Digestive Enzyme Activity, and Antioxidation of Juvenile Hucho taimen. Fishes. 2023; 8(12):567. https://doi.org/10.3390/fishes8120567
Chicago/Turabian StyleZhang, Meiyan, Xing Yang, Changan Wang, Baodi Shang, Feng Zhao, Hong Xu, and Qiyou Xu. 2023. "Effects of Myo-Inositol on the Growth Performance, Digestive Enzyme Activity, and Antioxidation of Juvenile Hucho taimen" Fishes 8, no. 12: 567. https://doi.org/10.3390/fishes8120567
APA StyleZhang, M., Yang, X., Wang, C., Shang, B., Zhao, F., Xu, H., & Xu, Q. (2023). Effects of Myo-Inositol on the Growth Performance, Digestive Enzyme Activity, and Antioxidation of Juvenile Hucho taimen. Fishes, 8(12), 567. https://doi.org/10.3390/fishes8120567