
Elevated Carbon Dioxide and Its Impact on Growth, Blood Properties, and Vertebral Column of Freshwater Fish Mahseer, Tor tambroides Juveniles



Abstract
:1. Introduction
2. Materials and Methods
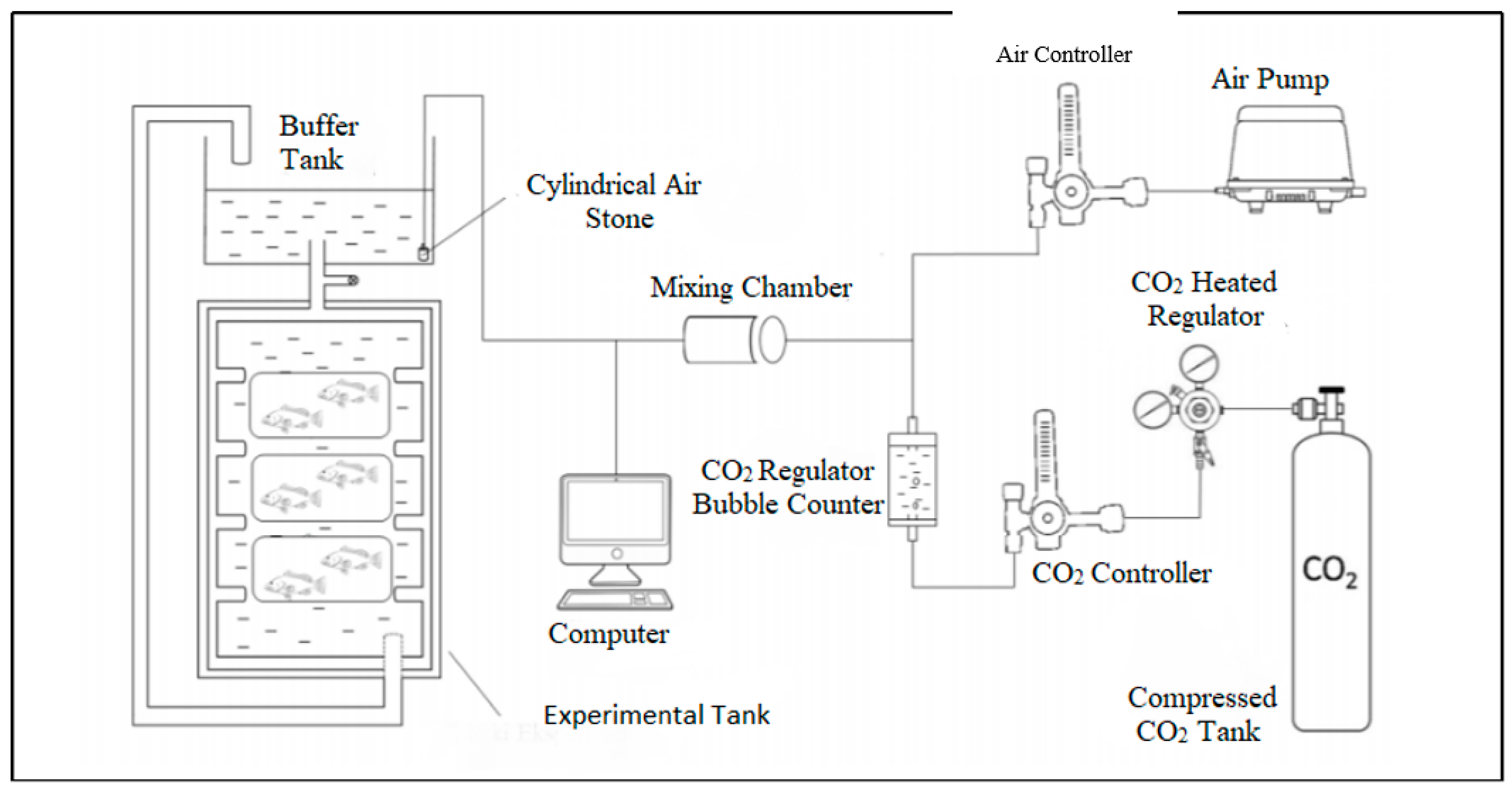
2.1. Fish Sampling and Experimental Design
2.2. Growth Properties
2.3. Vertebral Column Formation
2.4. Blood Haematology and Biochemical Parameters
2.5. Statistical Analysis
3. Results
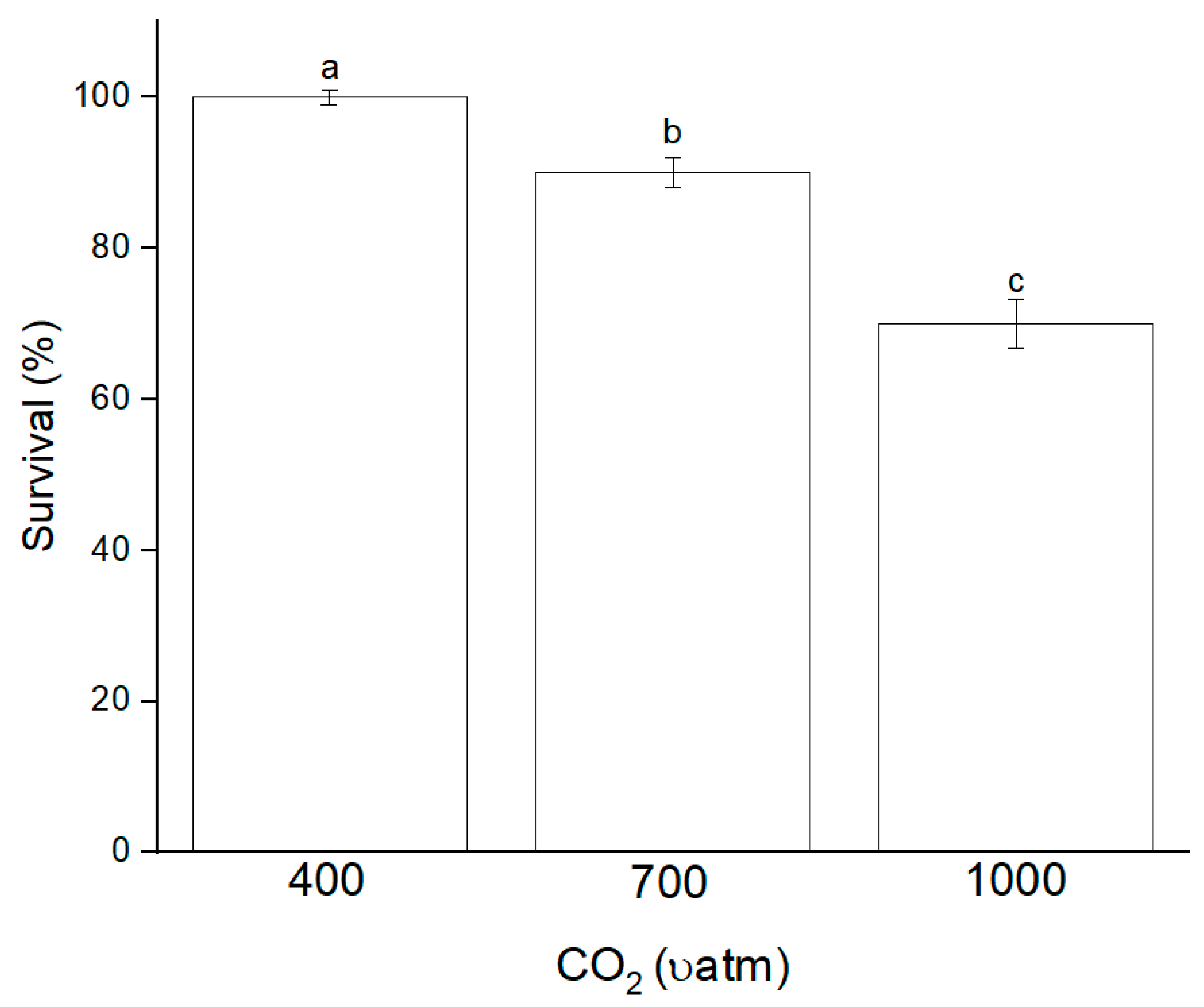
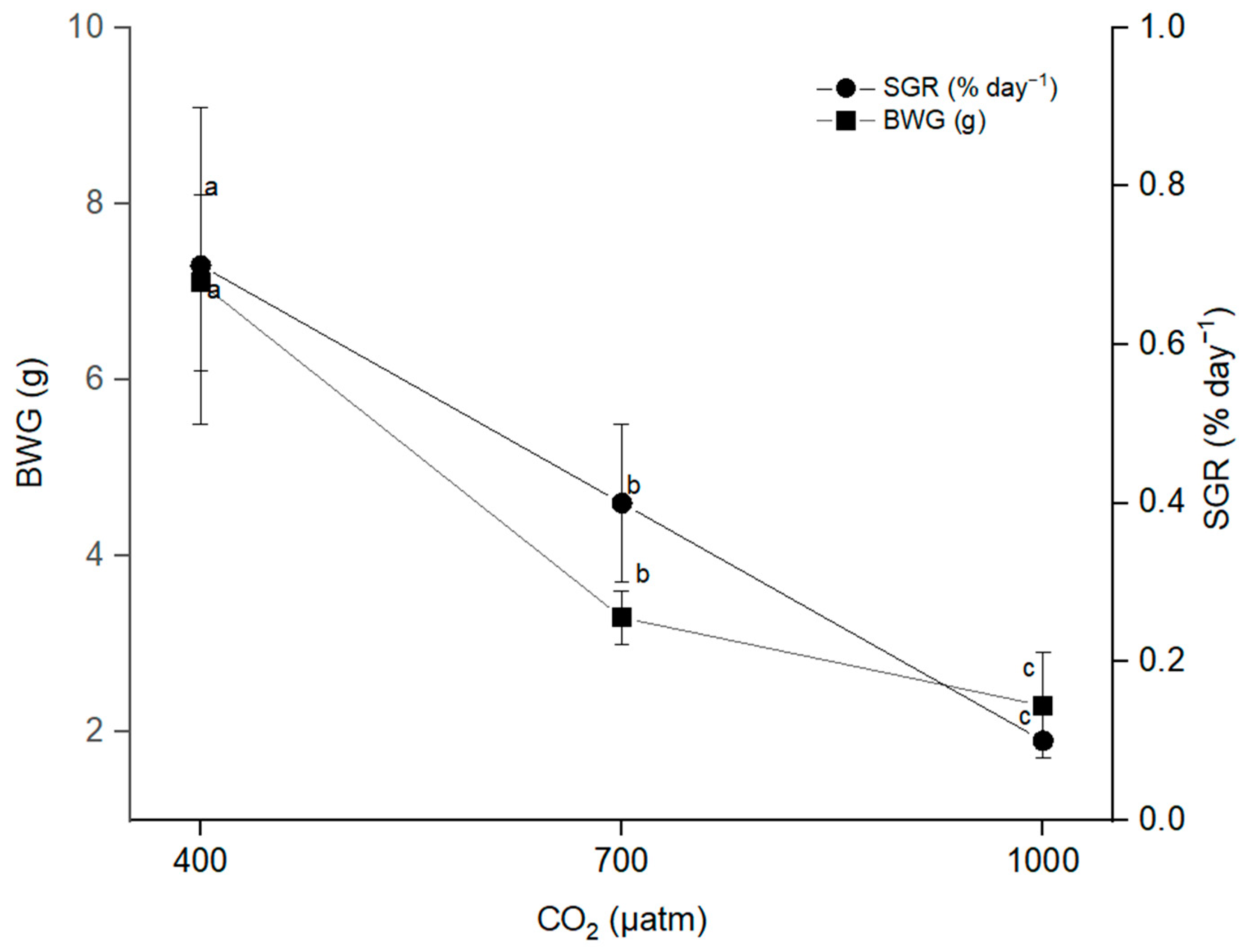
3.1. Growth
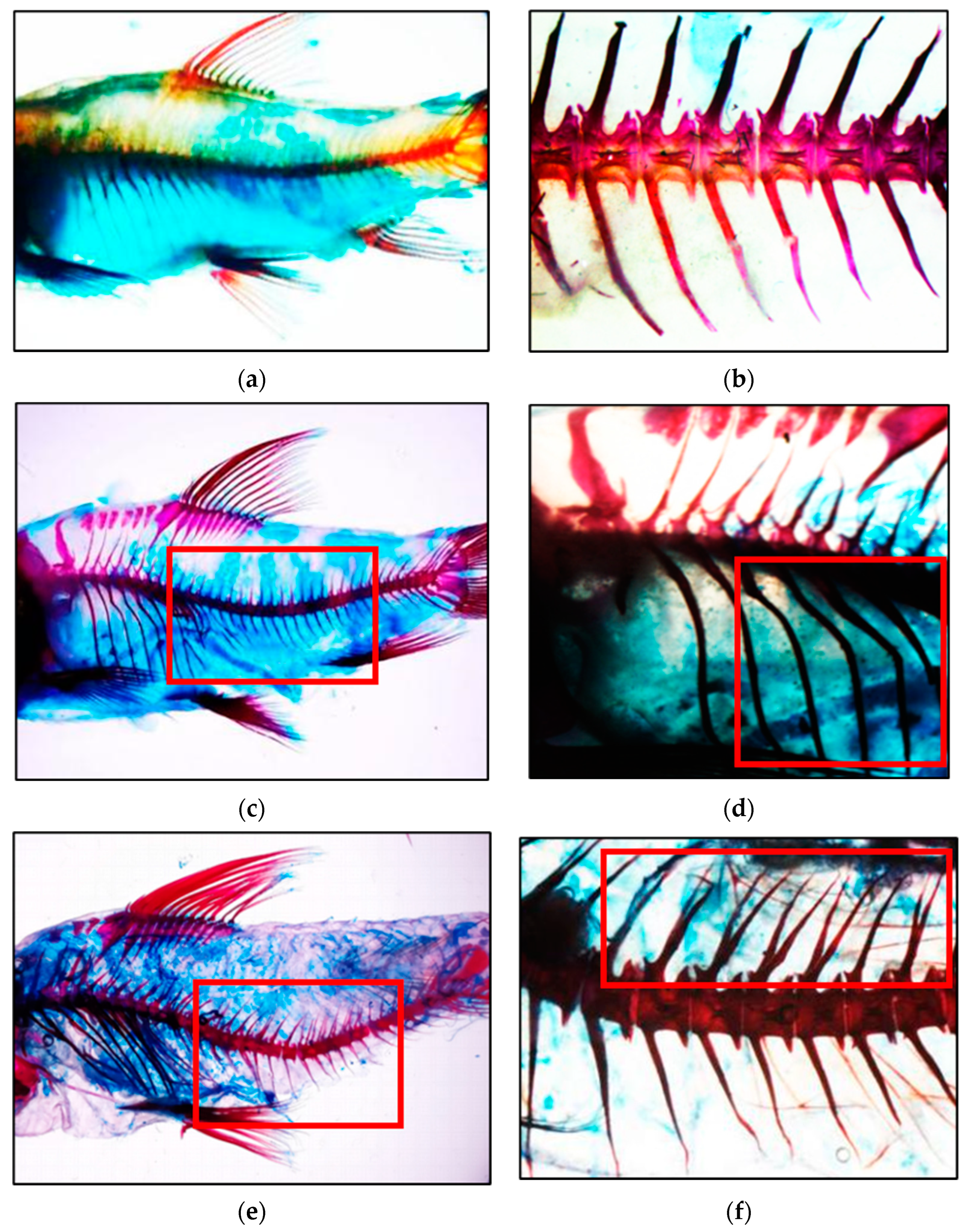
3.2. Vertebral Formation
3.3. Blood Haematological and Biochemical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parra, J.E.G.; Baldisserotto, B. Effect of water pH and hardness on survival and growth of freshwater teleosts. In Fish Osmoregulation; CRC Press: Boca Raton, FL, USA, 2019; pp. 135–150. [Google Scholar]
- Messina, S.; Costantini, D.; Eens, M. Impacts of rising temperatures and water acidification on the oxidative status and immune system of aquatic ectothermic vertebrates: A meta-analysis. Sci. Total Environ. 2023, 868, 161580. [Google Scholar] [CrossRef] [PubMed]
- Hasler, C.T.; Butman, D.; Jeffrey, J.D.; Suski, C.D. Freshwater biota and rising pCO2? Ecol. Lett. 2016, 19, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Walton, S.E.; Gan, H.M.; Raghavan, R.; Pinder, A.C.; Ahmad, A. Disentangling the taxonomy of the mahseers (Tor spp.) of Malaysia: An integrated approach using morphology, genetics and historical records. Rev. Fish. Sci. Aquac. 2017, 25, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Brauner, C.J.; Shartau, R.B.; Damsgaard, C.; Esbaugh, A.J.; Wilson, R.W.; Grosell, M. Acid-base physiology and CO2 homeostasis: Regulation and compensation in response to elevated environmental CO2. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2019; Volume 37, pp. 69–132. [Google Scholar]
- Jaafar, F.; Na-Nakorn, U.; Srisapoome, P.; Amornsakun, T.; Duong, T.Y.; Gonzales-Plasus, M.M.; Parhar, I.S. A current update on the distribution, morphological features, and genetic identity of the Southeast Asian mahseers, Tor species. Biology 2021, 10, 286. [Google Scholar] [CrossRef]
- Lopes, I.G.; Araújo-Dairiki, T.B.; Kojima, J.T.; Val, A.L.; Portella, M.C. Predicted 2100 climate scenarios affects growth and skeletal development of tambaqui (Colossoma macropomum) larvae. Ecol. Evol. 2018, 8, 10039–10048. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, A.; Pandey, M.; Kayastha, P.; Suwal, G.; Suwal, B. An Overview of Status and Development Trend of Aquaculture and Fisheries in Nepal. Adv. Agric. 2022, 2022, 4206401. [Google Scholar] [CrossRef]
- Stiasny, M.H.; Sswat, M.; Mittermayer, F.H.; Falk-Petersen, I.B.; Schnell, N.K.; Puvanendran, V.; Clemmesen, C. Divergent responses of Atlantic cod to ocean acidification and food limitation. Glob. Chang. Biol. 2019, 25, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Jesus, T.F.; Rosa, I.C.; Repolho, T.; Lopes, A.R.; Pimentel, M.S.; Almeida-Val, V.M.; Rosa, R. Different ecophysiological responses of freshwater fish to warming and acidification. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 216, 34–41. [Google Scholar] [CrossRef]
- Majeed, Z.; Ajab, Z.; Zuberi, A.; Akthar, S.; Muhammad, A. Meristic variations and skeletal deformities in natural population of mahseer fish, Tor putitora (Hamilton, 1822). Iran. J. Fish. Sci. 2018, 17, 208–216. [Google Scholar]
- Mazumder, S.K.; Das, S.K.; Bakar, Y.; Ghaffar, M.A. Effects of temperature and diet on length-weight relationship and condition factor of the juvenile Malabar blood snapper (Lutjanus malabaricus Bloch & Schneider, 1801). J. Zhejiang Univ. Sci. B 2016, 17, 580–590. [Google Scholar]
- Noor, N.M.; De, M.; Cob, Z.C.; Das, S.K. Welfare of scaleless fish, Sagor catfish (Hexanematichthys sagor) juveniles under different carbon dioxide concentrations. Aquac. Res. 2021, 52, 2980–2987. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.R.; Kim, S.K.; Kang, H.W. Effects of pH changes on blood physiology, antioxidant responses and Ig M of juvenile olive flounder, Paralichthys olivaceus. Aquac. Rep. 2021, 21, 100790. [Google Scholar] [CrossRef]
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Turak, E.; Heino, J.; Znachor, P. Characteristics, main impacts, and stewardship of natural and artificial freshwater environments: Consequences for biodiversity conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef] [Green Version]
- Greaver, T.L.; Clark, C.M.; Compton, J.E.; Vallano, D.; Talhelm, A.F.; Weaver, C.P.; Haeuber, R.A. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Chang. 2016, 6, 836–843. [Google Scholar] [CrossRef]
- Weiss, L.C.; Pötter, L.; Steiger, A.; Kruppert, S.; Frost, U.; Tollrian, R. Rising pCO2 in freshwater ecosystems has the potential to negatively affect predator-induced defenses in Daphnia. Curr. Biol. 2018, 28, 327–332. [Google Scholar] [CrossRef] [Green Version]
- De, M.; Ghaffar, M.A.; Bakar, Y.; Das, S.K. Effect of temperature and diet on growth and gastric emptying time of the hybrid, Epinephelus fuscoguttatus × E. lanceolatus. Aquac. Rep. 2016, 4, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Noor, N.M.; Kai, K.S.; Juan, Q.Z.; Iskandar, N.S.M.; De, M. Effects of temperature on the growth, gastric emptying time, and oxygen consumption rate of mahseer (Tor tambroides) under laboratory conditions. Aquac. Rep. 2018, 12, 20–24. [Google Scholar] [CrossRef]
- IPCC. AR6 Climate Change 2021: Impacts, Adaptation and Vulnerability—IPCC. 2021. Available online: https://www.ipcc.ch/report/sixth-assessment-report-working-group-ii/ (accessed on 28 December 2022).
- Phillips, J.C.; McKinley, G.A.; Bennington, V.; Bootsma, H.A.; Pilcher, D.J.; Sterner, R.W.; Urban, N.R. The potential for CO2-induced acidification in freshwater: A Great Lakes case study. Oceanography 2015, 28, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Kortazar, L.; Duval, B.; Liñero, O.; Olamendi, O.; Angulo, A.; Amouroux, D.; Fernandez, L.A. Accurate determination of the total alkalinity and the CO2 system parameters in high-altitude lakes from the Western Pyrenees (France–Spain). Microchem. J. 2020, 152, 104345. [Google Scholar] [CrossRef]
- Gupta, N.; Nigar, S. Detection of Blood Parasites and Estimation of Hematological Indices in Fish. In Experimental Protocols in Biotechnology; Humana: New York, NY, USA, 2020; pp. 43–73. [Google Scholar]
- Hoseini, S.M.; Mirghaed, A.T.; Iri, Y.; Ghelichpour, M. Effects of dietary cineole administration on growth performance, hematological and biochemical parameters of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 495, 766–772. [Google Scholar] [CrossRef]
- Kunwar, P.S.; Sinha, A.K.; De Boeck, G.; Sapkota, K. Modulations of blood biochemical parameters of golden mahseer, Tor putitora following exposures to single and mixed organophosphate. Comparat. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 251, 109207. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Howes, E.L.; Lacoue-Labarthe, T.; Forêt, S.; Hanna, B.; Medina, M.; Gattuso, J.P. Near-future pH conditions severely impact calcification, metabolism and the nervous system in the pteropod Heliconoides inflatus. Glob. Chang. Biol. 2016, 22, 3888–3900. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, J.D.; Hannan, K.D.; Hasler, C.T.; Suski, C.D. Responses to elevated CO2 exposure in a freshwater mussel, Fusconaia flava. J. Comparat. Physiol. B 2017, 187, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Tix, J.A.; Hasler, C.T.; Sullivan, C.; Jeffrey, J.D.; Suski, C.D. Elevated carbon dioxide has the potential to impact alarm cue responses in some freshwater fishes. Aquat. Ecol. 2017, 51, 59–72. [Google Scholar] [CrossRef]
- Mustapha, M.K.; Atolagbe, S.D. Tolerance level of different life stages of Nile tilapia Oreochromis niloticus (Linnaeus, 1758) to low pH and acidified waters. J. Basic Appl. Zool. 2018, 79, 46–54. [Google Scholar] [CrossRef]
- Chen, Y.; Bai, Y.; Hu, X.; Yang, X.; Xu, S. Energy metabolism responses in muscle tissue of rainbow trout Oncorhynchus mykiss fry to CO2-induced aquatic acidification based on metabolomics. Aquatic Toxicol. 2020, 220, 105400. [Google Scholar] [CrossRef]
- Ou, M.; Hamilton, T.J.; Eom, J.; Lyall, E.M.; Gallup, J.; Jiang, A. Responses of pink salmon to CO2-induced aquatic acidification. Nat. Clim. Chang. 2015, 5, 950–955. [Google Scholar] [CrossRef]
- Heino, J.; Virkkala, R.; Toivonen, H. Climate change and freshwater biodiversity: Detected patterns, future trends and adaptations in northern regions. Biol. Rev. 2009, 84, 39–54. [Google Scholar] [CrossRef]
- Wrona, F.J.; Prowse, T.D.; Reist, J.D.; Hobbie, J.E.; Lévesque, L.M.; Vincent, W.F. Climate change effects on aquatic biota, ecosystem structure and function. AMBIO 2006, 35, 359–369. [Google Scholar] [CrossRef] [Green Version]
- McMillan, J.R.; Katz, S.L.; Pess, G.R. Observational evidence of spatial and temporal structure in a sympatric anadromous (winter steelhead) and resident rainbow trout mating system on the Olympic Peninsula, Washington. Trans. Am. Fish. Soc. 2007, 136, 736–748. [Google Scholar] [CrossRef]
- Munday, P.L.; Watson, S.A.; Parsons, D.M.; King, A.; Barr, N.G.; Mcleod, I.M.; Pether, S.M. Effects of elevated CO2 on early life history development of the yellowtail kingfish, Seriola lalandi, a large pelagic fish. ICES J. Mar. Sci. 2016, 73, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Noor, N.M.; De, M.; Iskandar, A.; Keng, W.L.; Cob, Z.C.; Ghaffar, M.A.; Das, S.K. Effects of elevated carbon dioxide on the growth and welfare of Juvenile tiger grouper (Epinephelus fuscoguttatus) × giant grouper (E. lanceolatus) hybrid. Aquaculture 2019, 513, 734448. [Google Scholar] [CrossRef]
- Noor, N.M.; Das, S.K. Effects of elevated carbon dioxide on marine ecosystem and associated fishes. Thalassas 2019, 35, 421–429. [Google Scholar] [CrossRef]
- Wood, C.M. Internal spatial and temporal CO2 dynamics: Fasting, feeding, drinking, and the alkaline tide. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2019; Volume 37, pp. 245–286. [Google Scholar]
- Mazumder, S.K.; Das, S.K.; Rahim, S.M.; Abd Ghaffar, M. Temperature and diet effect on the pepsin enzyme activities, digestive somatic index and relative gut length of Malabar blood snapper (Lutjanus malabaricus Bloch & Schneider, 1801). Aquac. Rep. 2018, 9, 1–9. [Google Scholar]
- Kaya, H.; Hisar, O.; Yılmaz, S.; Gürkan, M.; Hisar, Ş.A. The effects of elevated carbon dioxide and temperature levels on tilapia (Oreochromis mossambicus): Respiratory enzymes, blood pH and hematological parameters. Environ. Toxicol. Pharmacol. 2016, 44, 114–119. [Google Scholar] [CrossRef]
- Sopinka, N.M.; Donaldson, M.R.; O’Connor, C.M.; Suski, C.D.; Cooke, S.J. Stress indicators in fish. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 405–462. [Google Scholar]
- Hoseini, S.M.; Mirghaed, A.T.; Mazandarani, M.; Zoheiri, F. Serum cortisol, glucose, thyroid hormones’ and non-specific immune responses of Persian sturgeon, Acipenser persicus to exogenous tryptophan and acute stress. Aquaculture 2016, 462, 17–23. [Google Scholar] [CrossRef]
- Vargas-Chacoff, L.; Regish, A.M.; Weinstock, A.; McCormick, S.D. Effects of elevated temperature on osmoregulation and stress responses in Atlantic salmon Salmo salar smolts in fresh water and seawater. J. Fish Biol. 2018, 93, 550–559. [Google Scholar] [CrossRef]
- Martens, L.G.; Witten, P.E.; Fivelstad, S.; Huysseune, A.; Sævareid, B.; Vikeså, V.; Obach, A. Impact of high water carbon dioxide levels on Atlantic salmon smolts (Salmo salar L.): Effects on fish performance, vertebrae composition and structure. Aquaculture 2006, 261, 80–88. [Google Scholar] [CrossRef]
- Hu, J.; Liu, Y.; Yu, G.; Yang, C.; Shan, B.; Liu, S.; Sun, D. Osteological development of the vertebral column and caudal complex in larval and juvenile blackhead seabream, Acanthopagrus schlegelii (Perciformes, Sparidae) (Bleeker, 1854). Pak. J. Zool. 2019, 51, 1859. [Google Scholar] [CrossRef]
- Crespel, A.; Dupont-Prinet, A.; Bernatchez, L.; Claireaux, G.; Tremblay, R.; Audet, C. Divergence in physiological factors affecting swimming performance between anadromous and resident populations of brook charr Salvelinus fontinalis. J. Fish Biol. 2017, 90, 2170–2193. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pCO2 (µatm) | Temperature (°C) | Total Alkalinity (mg L−1) | pH | Dissolved Oxygen (mg L−1) | Total Ammonia (mg/L) | Nitrite (mg/L) | Nitrate (mg/L) | |
|---|---|---|---|---|---|---|---|---|
| Current | 407.2 ± 10.5 a | 28.3 ± 0.1 | 19 ± 2 | 8.2 ± 0.1 | 5.8 ± 0.2 | 0.6 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.0 |
| Mild | 725.7 ± 20.1 b | 28.3 ± 0.2 | 12 ± 1 | 7.9 ± 0.1 | 5.6 ± 0.4 | 0.7 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| High | 1020.6 ± 9.2 c | 28.1 ± 0.3 | 5 ± 1 | 7.7 ± 0.1 | 5.5 ± 0.3 | 0.7 ± 0.2 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| pCO2 (µatm) | Lordosis (%) | Fusion (%) | Scoliosis (%) | Kyphosis (%) |
|---|---|---|---|---|
| 400 | ND | ND | ND | ND |
| 700 | 6.67 ± 0.0 | ND | ND | ND |
| 1000 | 13.33 ± 3.33 | 3.33 ± 00 | ND | ND |
| pCO2 (µatm) | Hb (g/dL) | Hct (%) | Glucose (mmol/L) | Protein (g/dL) | Cholesterol (mmol/L) |
|---|---|---|---|---|---|
| Reference interval [24] | 8.3–12.3 | 29.0–30.0 | 1.9–2.7 | 3.0–3.9 | 4.08–4.11 |
| 400 | 9.8 ± 1.1 a | 29.1 ± 1.5 a | 2.0 ± 0.1 a | 3.6 ± 0.3 a | 4.1 ± 0.1 a |
| 700 | 13.8 ± 0.7 b | 25.2 ± 2.0 b | 1.6 ± 0.2 b | 2.9 ± 0.1 b | 3.7 ± 0.1 b |
| 1000 | 14.6 ± 0.5 c | 20.5 ± 1.8 c | 1.1 ± 0.3 c | 2.7 ± 0.1 c | 3.5 ± 0.1 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iskandar, N.S.; Noor, N.M.; Cob, Z.C.; Das, S.K. Elevated Carbon Dioxide and Its Impact on Growth, Blood Properties, and Vertebral Column of Freshwater Fish Mahseer, Tor tambroides Juveniles. Fishes 2023, 8, 307. https://doi.org/10.3390/fishes8060307
Iskandar NS, Noor NM, Cob ZC, Das SK. Elevated Carbon Dioxide and Its Impact on Growth, Blood Properties, and Vertebral Column of Freshwater Fish Mahseer, Tor tambroides Juveniles. Fishes. 2023; 8(6):307. https://doi.org/10.3390/fishes8060307
Chicago/Turabian StyleIskandar, Nur Syuhada, Noorashikin Md Noor, Zaidi Che Cob, and Simon Kumar Das. 2023. "Elevated Carbon Dioxide and Its Impact on Growth, Blood Properties, and Vertebral Column of Freshwater Fish Mahseer, Tor tambroides Juveniles" Fishes 8, no. 6: 307. https://doi.org/10.3390/fishes8060307
APA StyleIskandar, N. S., Noor, N. M., Cob, Z. C., & Das, S. K. (2023). Elevated Carbon Dioxide and Its Impact on Growth, Blood Properties, and Vertebral Column of Freshwater Fish Mahseer, Tor tambroides Juveniles. Fishes, 8(6), 307. https://doi.org/10.3390/fishes8060307