
Biochemical Patterns and Genotoxicity of the Endocrine Disruptor Metformin in the Freshwater Fish Labeo rohita

 , , ,
, , , 
 and
and 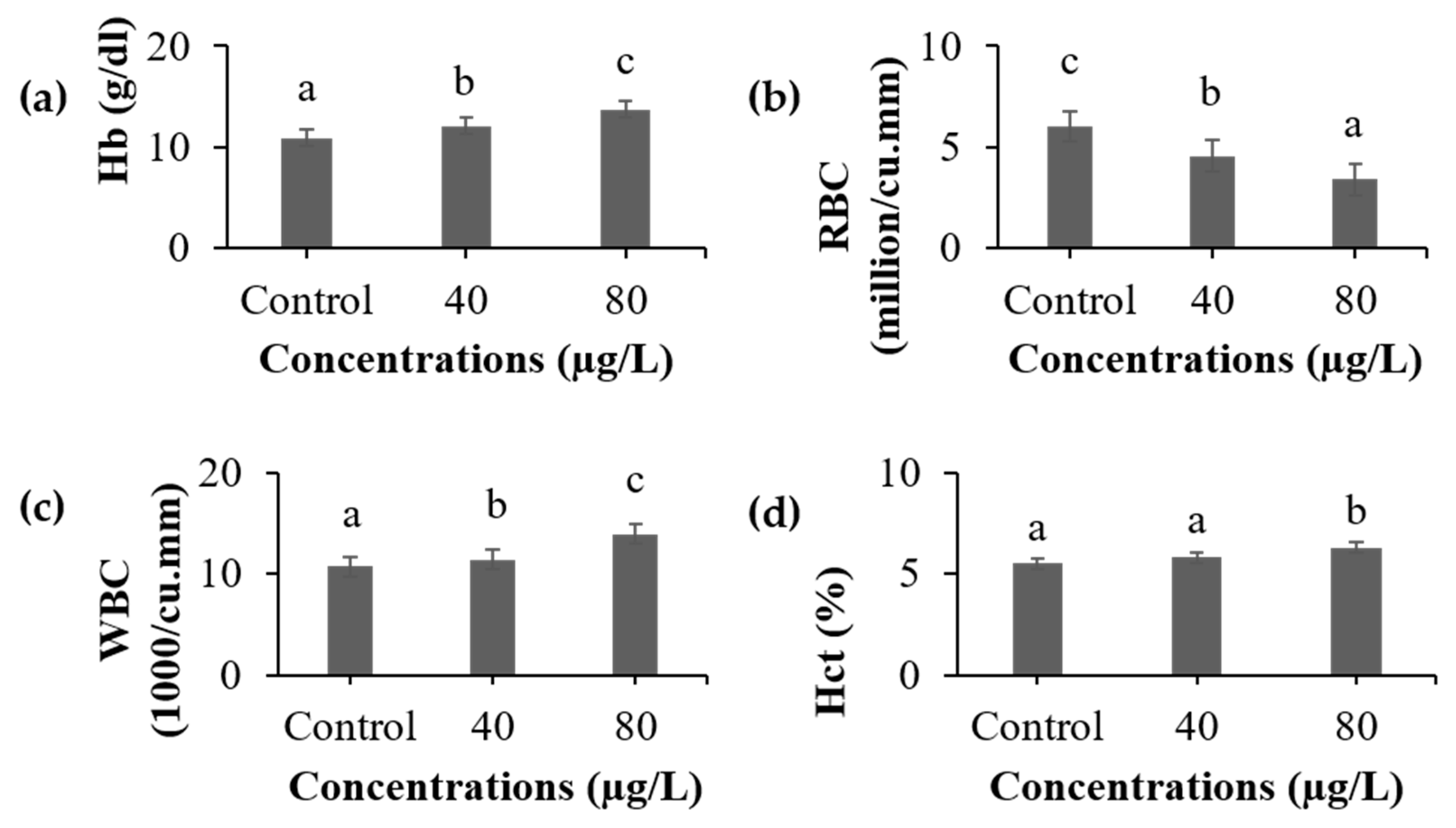{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Animals
2.3. Experimental Setup and Sample Collection
2.4. Hematological Analysis
2.5. Sample Preparation for Biochemical Assays
2.6. Indicators of Cellular Damage
2.7. Non-Specific Immune Parameters
2.8. Genotoxicity (Comet Assay)
2.9. Statistical Analysis
3. Results and Discussion
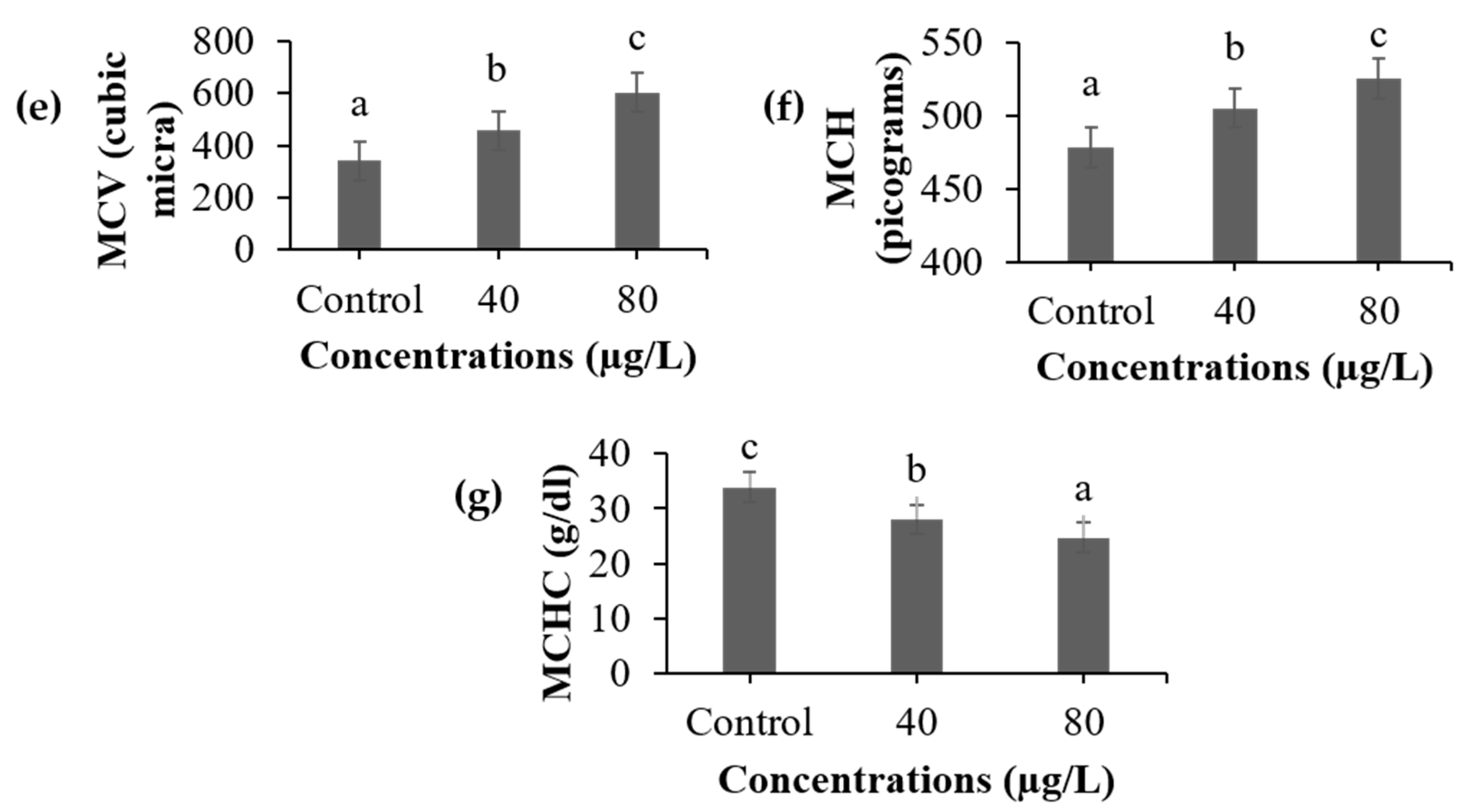
3.1. Hematological Analysis
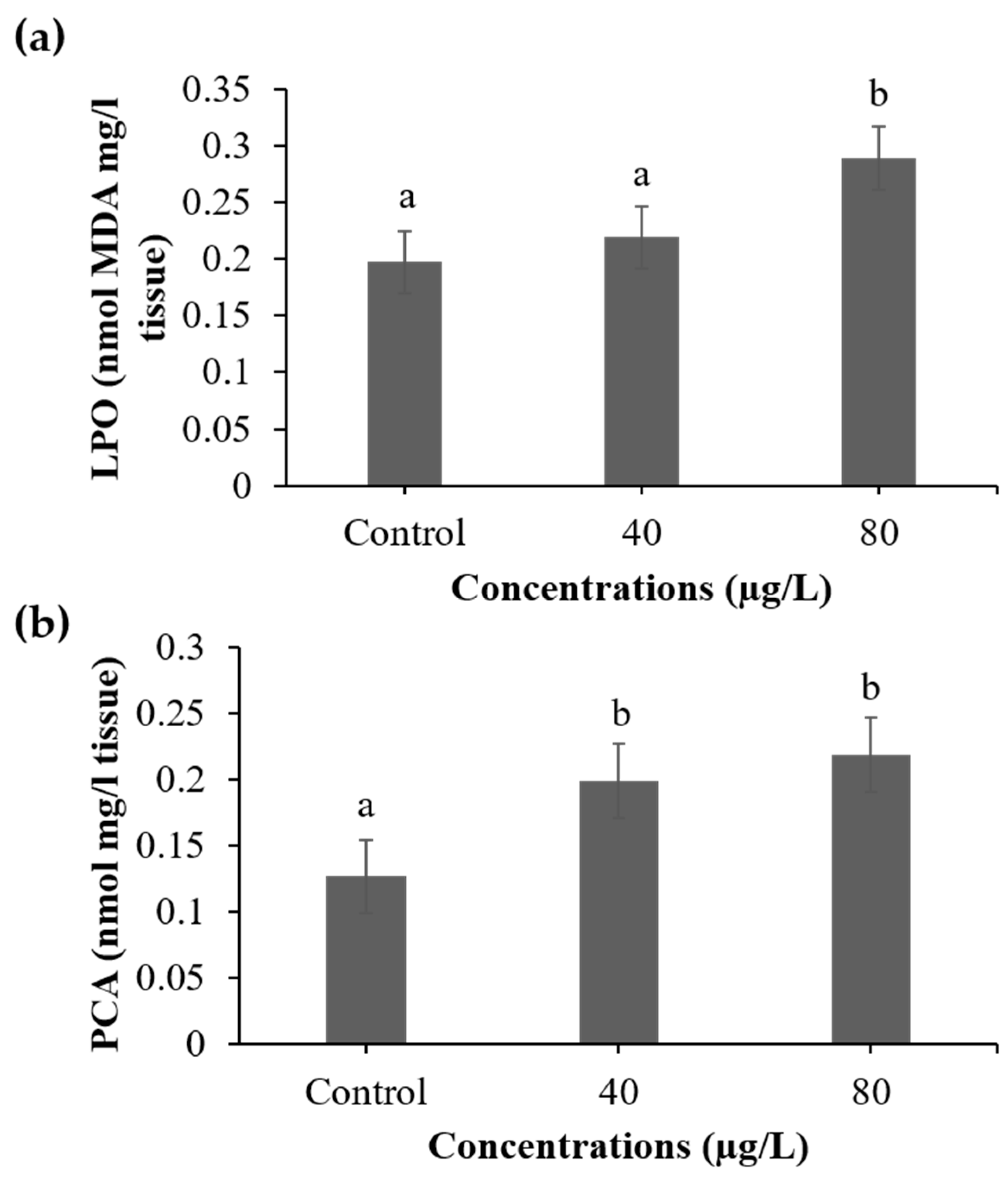
3.2. Indicators of Cellular Damage
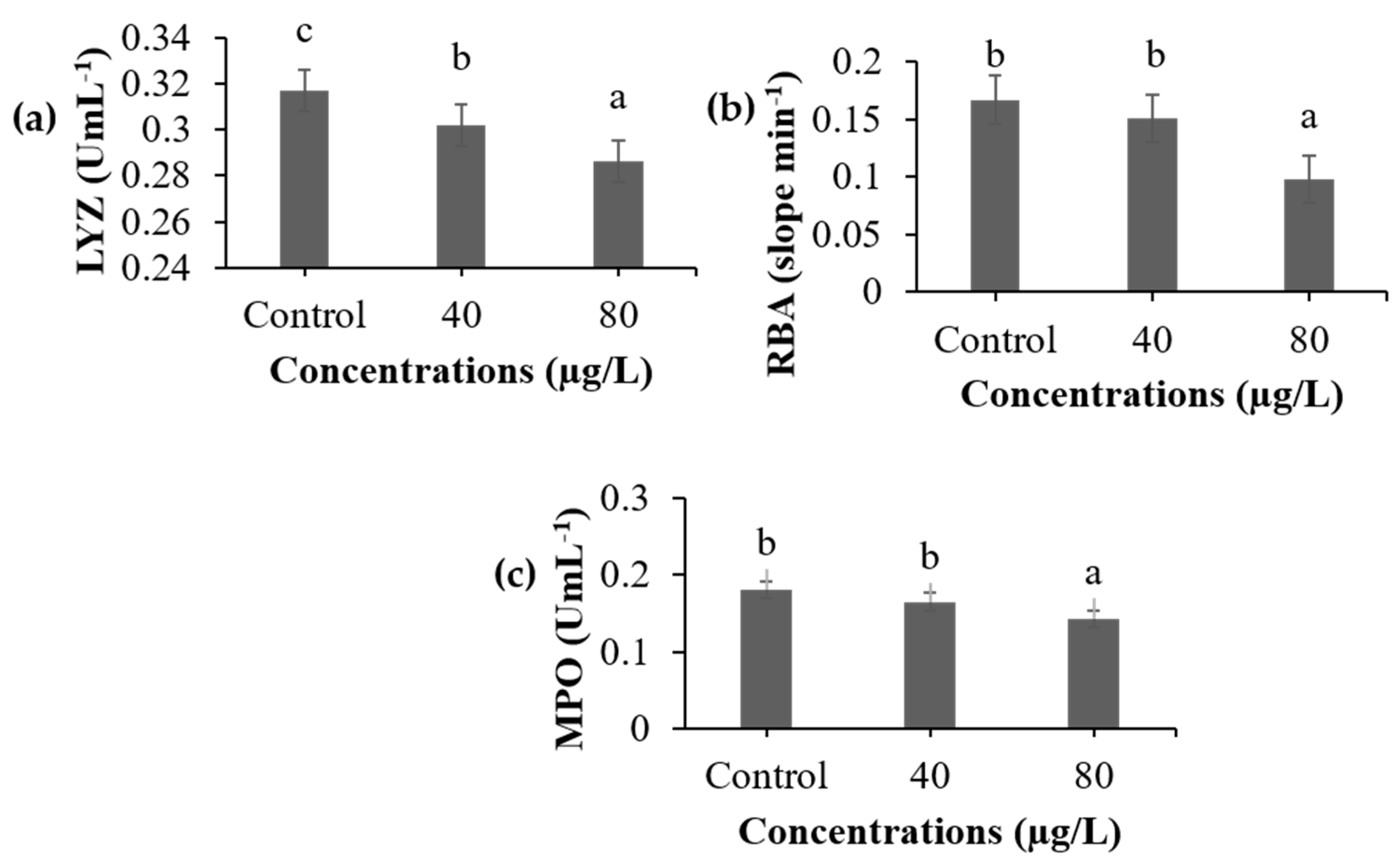
3.3. Non-Specific Immune Parameters
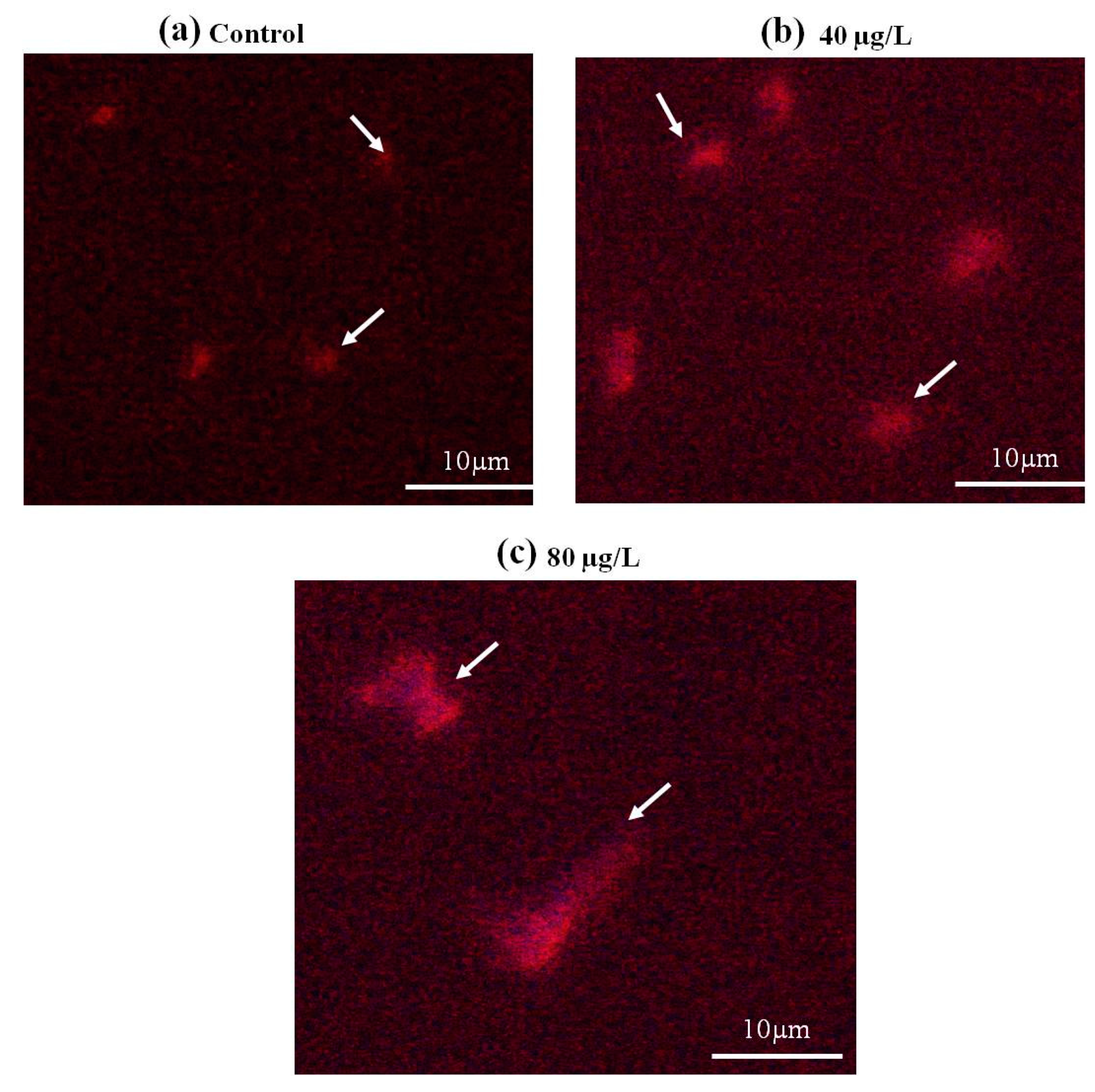
3.4. Genotoxicity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The worldwide epidemiology of type 2 diabetes mellitus present and future perspectives. Nat. Rev. Endocrinol. 2012, 4, 228–236. [Google Scholar] [CrossRef]
- Ghoshdastidar, A.J.; Fox, S.; Tong, A.Z. The presence of the top prescribed pharmaceuticals in treated sewage effluents and receiving waters in Southwest Nova Scotia, Canada. Environ. Sci. Pollut. Res. 2015, 1, 689–700. [Google Scholar] [CrossRef]
- Jacob, S.; Dötsch, A.; Knoll, S.; Köhler, H.R.; Rogall, E.; Stoll, D.; Tisler, S.; Huhn, C.; Schwartz, T.; Zwiener, C.; et al. Does the antidiabetic drug metformin affect embryo development and the health of brown trout (Salmo trutta f. fario)? Environ. Sci. Eur. 2018, 30, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oosterhuis, M.; Sacher, F.; Ter Laak, T.L. Prediction of concentration levels of metformin and other high consumption pharmaceuticals in wastewater and regional surface water based on sales data. Sci. Total Environ. 2013, 442, 380–388. [Google Scholar] [CrossRef]
- Viollet, B.; Guigas, B.; Garcia, N.S.; Leclerc, J.; Foretz, M.; Andreelli, F. Cellular and molecular mechanisms of metformin: An overview. Clin. Sci. 2012, 122, 253–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piltonen, T.; Morin-Papunen, L.; Koivunen, R.; Perheentupa, A.; Ruokonen, A.; Tapanainen, J.S. Serum anti-Müllerian hormone levels remain high until late reproductive age and decrease during metformin therapy in women with polycystic ovary syndrome. Hum Reprod. 2005, 20, 1820–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazer, V.S.; Iwanowicz, L.R.; Henderson, H.; Mazik, P.M.; Jenkins, J.A.; Alvarez, D.A.; Young, J.A. Reproductive endocrine disruption in smallmouth bass (Micropterus dolomieu) in the Potomac River basin: Spatial and temporal comparisons of biological effects. Environ. Monit. Assess. 2012, 184, 4309–4334. [Google Scholar] [CrossRef] [Green Version]
- Niemuth, N.J.; Klaper, R.D. Low-dose metformin exposure causes changes in expression of endocrine disruption-associated genes. Aquat. Toxicol. 2018, 195, 33–40. [Google Scholar] [CrossRef]
- Cui, J.; Hao, Z.; Zhou, Q.; Qiu, M.; Liu, Y.; Liu, Y.; Teng, X.; Kang, L. Chlorpyrifos induced autophagy and mitophagy in common carp livers through AMPK pathway activated by energy metabolism disorder. Ecotoxicol. Environ. Saf. 2023, 258, 114983. [Google Scholar] [CrossRef]
- Cui, J.; Qiu, M.; Liu, Y.; Liu, Y.; Teng, X.; Li, S. Nano-selenium protects grass carp hepatocytes against 4-tert-butylphenol-induced mitochondrial apoptosis and necroptosis via suppressing ROS-PARP1 axis. Fish Shellfish. Immunol. 2023, 135, 108682. [Google Scholar] [CrossRef]
- Zhao, C.; Teng, X.; Yue, W.; Suo, A.; Zhou, W.; Ding, D. The effect of acute toxicity from tributyltin on Liza haematocheila liver: Energy metabolic disturbance, oxidative stress, and apoptosis. Aquatic. Toxicol. 2023, 258, 106506. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Xu, W.; Zhang, Y.; Sun, Q.; Li, Z.; Geng, L.; Teng, X. Transcriptome analysis revealed the mechanism of Luciobarbus capito (L. capito) adapting high salinity: Antioxidant capacity, heat shock proteins, immunity. Marine Pollut. Bull. 2023, 192, 115017. [Google Scholar]
- Sibiya, A.; Gopi, N.; Jeyavani, J.; Mahboob, S.; Al-Ghanim, K.A.; Sultana, S.; Mustafa, A.; Govindarajan, M.; Vaseeharan, B. Comparative toxicity of silver nanoparticles and silver nitrate in freshwater fish Oreochromis mossambicus: A multi-biomarker approach. Comp. Biochem. Physiol. C Pharmacol. Toxicol. 2022, 2, 109391. [Google Scholar] [CrossRef] [PubMed]
- MacLaren, R.D.; Wisniewski, K.; MacLaren, C. Environmental concentrations of metformin exposure affect aggressive behavior in the Siamese fighting fish, Betta splendens. PLoS ONE 2018, 13, 0197259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drabkin, D.L. Spectrometric studies, XIV: The crystallographic and optimal properties of the hemoglobin of man in comparison with those of other species. J. Biol. Chem. 1946, 164, 703–723. [Google Scholar] [CrossRef] [PubMed]
- Sivanandan, J.M. Acute and sublethal intoxication of malathion in an Indian major carp, Labeo rohita: Haematological and biochemical responses. Environ. Anal. Health Toxicol. 2021, 36, e2021016. [Google Scholar] [CrossRef]
- Nwani, C.D.; Mkpadobi, B.N.; Onyishi, G.; Echi, P.C.; Chukwuka, C.O.; Oluah, S.N.; Ivoke, N. Changes in behavior and hematological parameters of freshwater African catfish Clarias gariepinus (Burchell 1822) following sublethal exposure to chloramphenicol. Drug Chem. Toxicol. 2014, 37, 107–113. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1978; Volume 52, pp. 302–310. [Google Scholar]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar]
- Ellis, A.E. Lysozyme Assays. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., Van Muiswinkel, W.B., Eds.; SOS Publications: New York, NY, USA, 1990; pp. 101–103. [Google Scholar]
- Secombes, C. Isolation of salmonid macrophages and analysis of their killing activity. Tech. Fish Immunol. 1990, 137–154. [Google Scholar]
- Kumari, J.; Sahoo, P.K. Dietary β-1, 3 glucan potentiates innate immunity and disease resistance of Asian catfish. Clarias batrachus (L.). J. Fish Dis. 2006, 29, 95–101. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantification of low levels of DNA damage in individual cells. Exp. Cell. Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, M.; Devi, K.U.; Malarvizhi, A.; Ramesh, M. Effects of Ibuprofen on hematological, biochemical and enzymological parameters of blood in an Indian major carp. Cirrhinus mrigala. Environ. Toxicol. Pharmacol. 2012, 34, 14–22. [Google Scholar] [CrossRef]
- Rezaei, M.; Mashinchian Moradi, A.; Mortazavi, P.; Jamili, S. Effects of chronic exposure to carbamazepine on hematological parameters in Cyprinus carpio. Iran. J. Fish. Sci. 2020, 19, 443–456. [Google Scholar]
- Ates, B.; Orun, I.; Talas, Z.S.; Durmaz, G.; Yilmaz, I. Effects of sodium selenite on some bio-chemical and hematological parameters of rainbow trout (Oncorhynchus mykiss Walbaum, 1792) exposed to Pb2+ and Cu2+. Fish Physiol. Biochem. 2008, 34, 53–59. [Google Scholar] [CrossRef]
- Li, Z.H.; Velisek, J.; Grabic, R.; Li, P.; Kolarova, J.; Randak, T. Use of hematological and plasma biochemical parameters to assess the chronic effects of a fungicide propiconazole on a freshwater teleost. Chemosphere 2011, 83, 572–578. [Google Scholar] [CrossRef]
- Li, Z.H.; Zlabek, V.; Velisek, J.; Grabic, R.; Machova, J.; Randak, T. Modulation of antioxidant defence system in brain of rainbow trout (Oncorhynchus mykiss) after chronic carbamazepine treatment. Comp. Biochem. Physiol. C Pharmacol. Toxicol. 2010, 1, 137–141. [Google Scholar] [CrossRef]
- Bols, N.C.; Brubacher, J.L.; Ganassin, R.C.; Lee, L.E. Ecotoxicology and innate immunity in fish. Dev. Comp. Immunol. 2001, 25, 853–873. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Esteban, M.Á.; Meseguer, J. Phagocytosis and peroxidase release by seabream (Sparus aurata L.) leucocytes in response to yeast cells. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 2003, 272, 415–423. [Google Scholar] [CrossRef]
- Yin, D.; Xu, Z.; Feng, M.; Zhao, Z.; Chen, D.; Song, L. Genotoxicity evaluation of metformin in freshwater planarian Dugesia japonica by the comet assay and RAPD analysis. Biomed Res Int. 2022, 2022, 2822605. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Tejada, S.; Blanco, A.; Caixach, J.; Deudero, S. Biochemical responses of Mytilus galloprovincialis as biomarkers of acute environmental pollution caused by the Don Pedro oil spill (Eivissa Island, Spain). Aquat. Toxicol. 2011, 101, 540–549. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sibiya, A.; Al-Ghanim, K.A.; Govindarajan, M.; Nicoletti, M.; Sachivkina, N.; Vaseeharan, B. Biochemical Patterns and Genotoxicity of the Endocrine Disruptor Metformin in the Freshwater Fish Labeo rohita. Fishes 2023, 8, 380. https://doi.org/10.3390/fishes8070380
Sibiya A, Al-Ghanim KA, Govindarajan M, Nicoletti M, Sachivkina N, Vaseeharan B. Biochemical Patterns and Genotoxicity of the Endocrine Disruptor Metformin in the Freshwater Fish Labeo rohita. Fishes. 2023; 8(7):380. https://doi.org/10.3390/fishes8070380
Chicago/Turabian StyleSibiya, Ashokkumar, Khalid A. Al-Ghanim, Marimuthu Govindarajan, Marcello Nicoletti, Nadezhda Sachivkina, and Baskaralingam Vaseeharan. 2023. "Biochemical Patterns and Genotoxicity of the Endocrine Disruptor Metformin in the Freshwater Fish Labeo rohita" Fishes 8, no. 7: 380. https://doi.org/10.3390/fishes8070380
APA StyleSibiya, A., Al-Ghanim, K. A., Govindarajan, M., Nicoletti, M., Sachivkina, N., & Vaseeharan, B. (2023). Biochemical Patterns and Genotoxicity of the Endocrine Disruptor Metformin in the Freshwater Fish Labeo rohita. Fishes, 8(7), 380. https://doi.org/10.3390/fishes8070380