

Tryptophan Reduces Intracohort Cannibalism Behavior in Tropical Gar (Atractosteus tropicus) Larvae

 , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Experimental Design
2.3. Formulation and Preparation of the Experimental Diets
2.4. Growth Indexes and Feed Quality
2.5. Collection of Biological Samples
2.6. Cannibalism
2.7. Digestive Enzyme Activity
2.8. RNA Extraction and Quantitative Reverse Transcription PCR (RT-qPCR)
2.9. Effect of the Mitigants on the Ethology of Cannibal Larvae
2.10. Statistics Analysis
3. Results
3.1. Growth Indexes and Survival
3.2. Cannibalism and Deformities
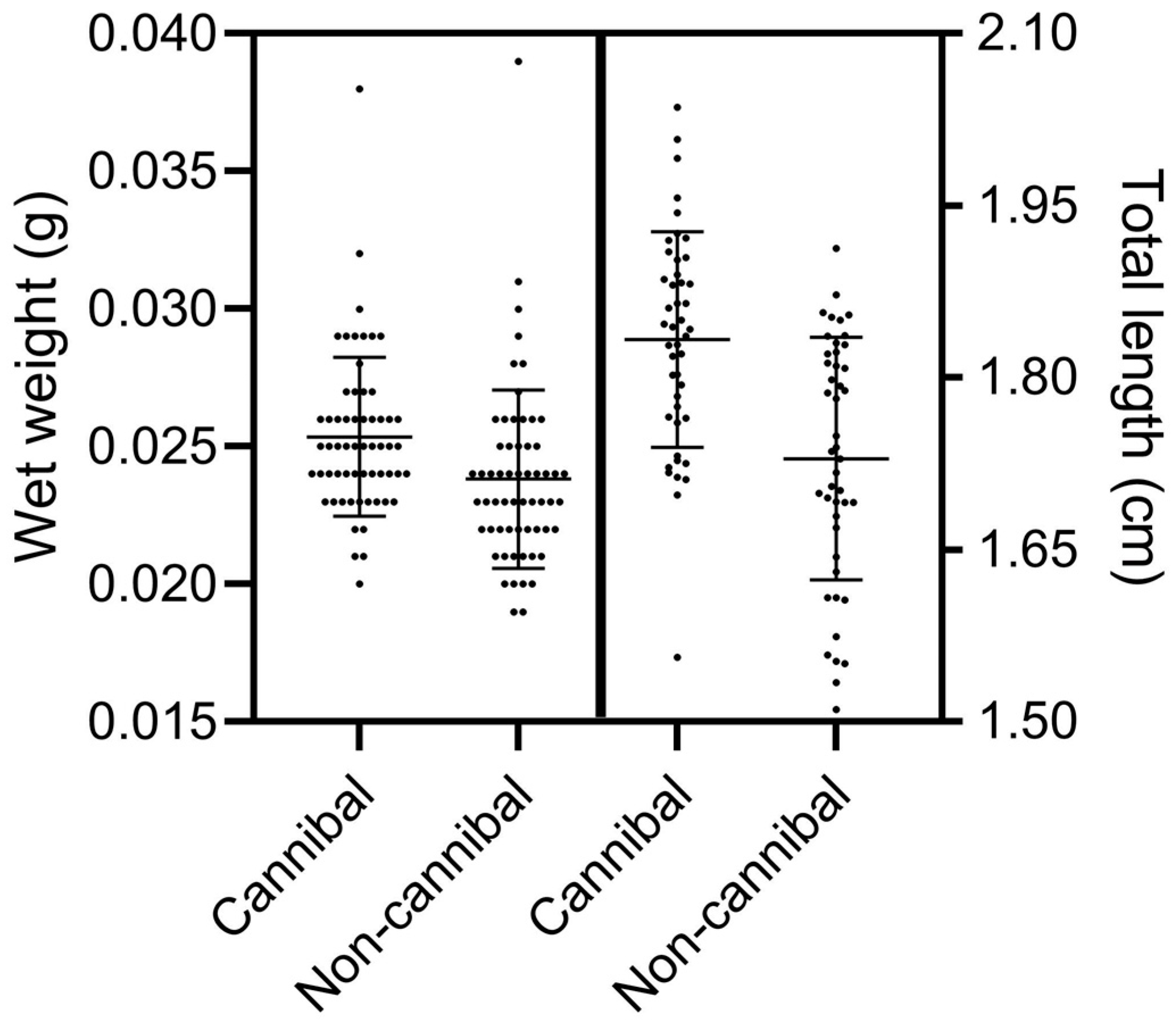
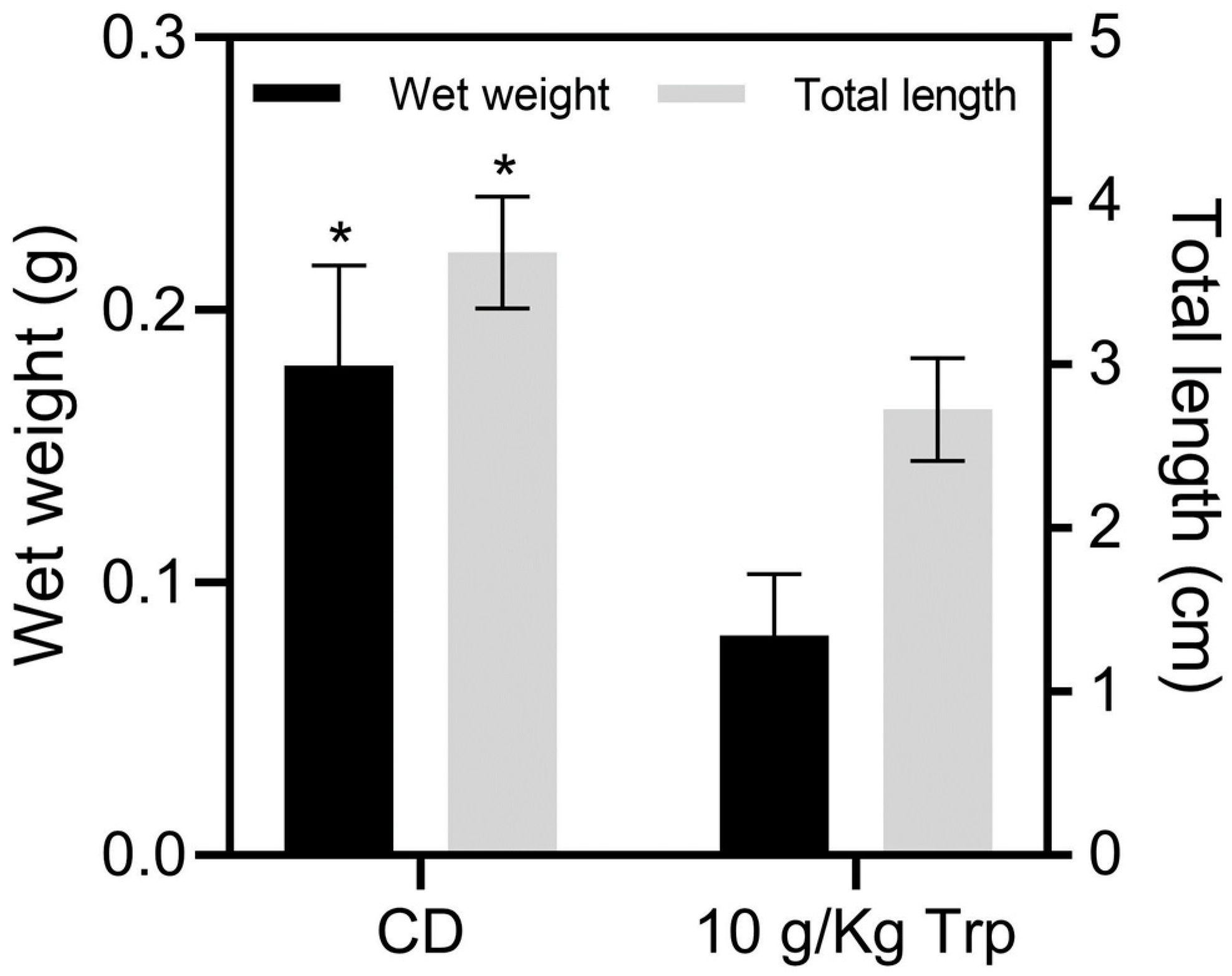
3.3. Growth Indexes and Survival of Cannibals
3.4. Cannibalism
3.5. Digestive Enzyme Activity
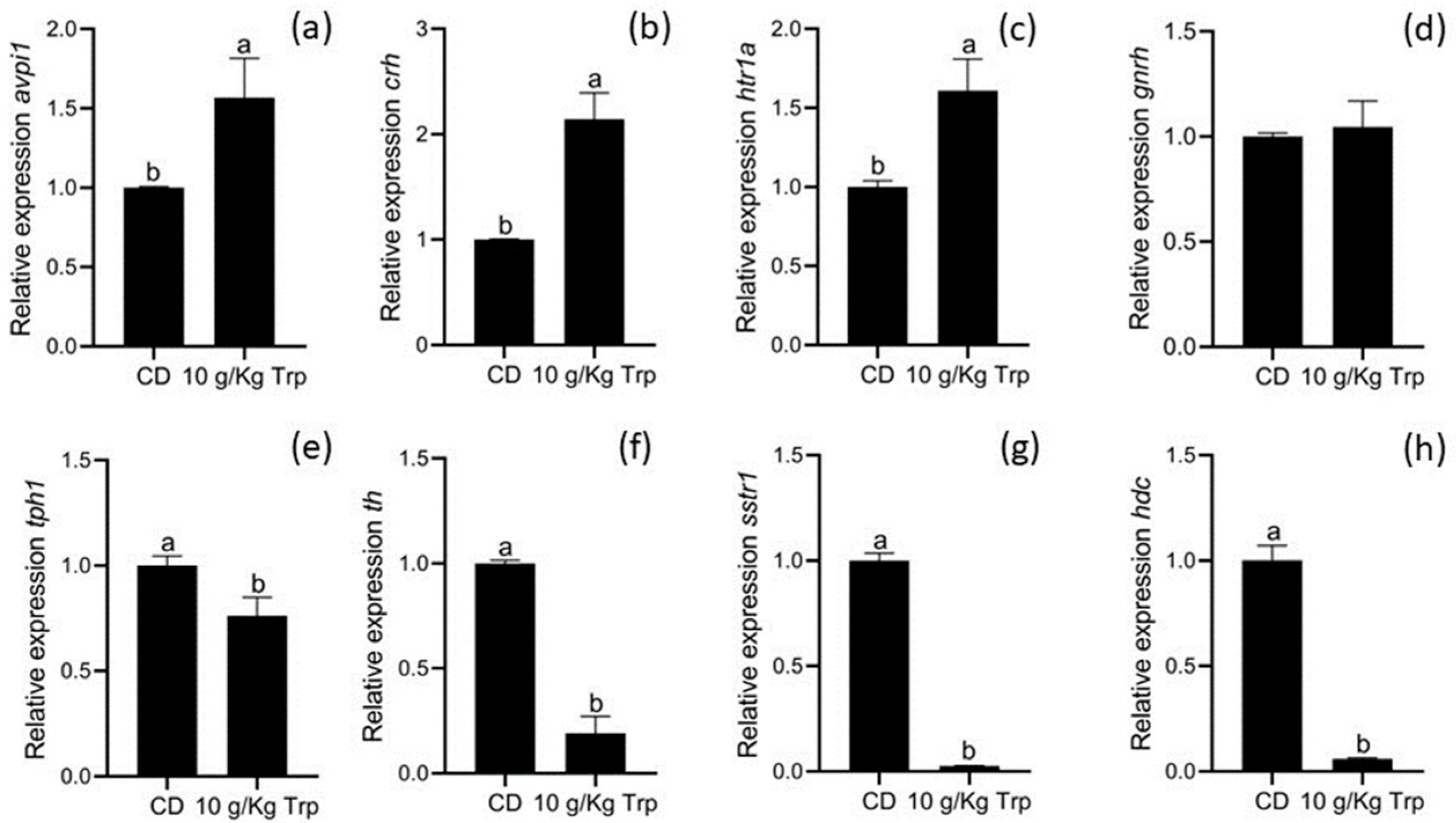
3.6. Gene Expression
3.7. Behavior
4. Discussion
4.1. Growth
4.2. Cannibalism
4.3. Digestive Enzymes
4.4. Gene Expression
4.5. Behavior
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira, L.S.; Agostinho, A.A.; Winemiller, K.O. Revisiting cannibalism in fishes. Rev. Fish Biol. Fish. 2017, 27, 499–513. [Google Scholar] [CrossRef]
- Aguilera, C.; Mendoza, R.; Iracheta, I.; Márquez, G. Digestive enzymatic activity on Tropical gar (Atractosteus tropicus) larvae fed different diets. Fish Physiol. Biochem. 2012, 38, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Márquez, C.G. Biología y tecnología para el cultivo del pejelagarto Atractosteus tropicus en el sureste de México. In Redes nacionales de Investigación en Acuicultura. Memorias de la IV Reunión. Secretaría de Medio Ambiente Recursos Naturales y Pesca (SEMARNAP); Alvarez, P., Guzman, M., Contreras, S., Silva, A., Eds.; Instituto Nacional de la Pesca: México City, Mexico, 2000; pp. 265–267. [Google Scholar]
- Márquez-Couturier, G.; Vázquez-Navarrete, C.J. Estado del arte de la biología y cultivo de pejelagarto (Atractosteus tropicus). Agroproductividad 2015, 8, 44–51. [Google Scholar]
- Nelson, J.S. Fishes of the World, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; 601p, ISBN 978-0471250319. [Google Scholar]
- Frías-Quintana, C.; Álvarez-González, C.; Tovar-Ramírez, D.; Martínez-García, R.; Camarillo-Coop, S.; Peña, E.; Galaviz, M. Use of potato starch in diets of tropical gar (Atractosteus tropicus, Gill 1863) larvae. Fishes 2017, 2, 3. [Google Scholar] [CrossRef]
- Palma-Cancino, D.J.; Martínez-García, R.; Álvarez-González, C.A.; Camarillo-Coop, S.; Peña-Marín, E.S. Evaluation of feeding strategies in tropical gar (Atractosteus tropicus Gill) larvae: Growth, survival and cannibalism. Ecosist. Recur. Agropecu. 2019, 6, 273–281. [Google Scholar] [CrossRef]
- Jiménez-Martínez, L.D.; Tovar-Ramírez, D.; Álvarez-González, C.A.; Peña-Marín, E.; Camarillo-Coop, S.; Martínez-García, R.; Palacios-Mechetnov, E.; Martínez-Yáñez, R.; Concha-Frias, B. Assessment of dietary lipid sources in tropical gar, Atractosteus tropicus larvae: Growth parameters and intermediary lipogenic gene expression. Aquac. Res. 2020, 51, 2629–2640. [Google Scholar] [CrossRef]
- Leathwood, P.D. Tryptophan availability and serotonin synthesis. Proc. Nutr. Soc. 1987, 46, 143–156. [Google Scholar] [CrossRef]
- Smith, T.K.; Seddon, I.R. Synergism demonstrated between Fusarium mycotoxins. Feedstuffs 1998, 70, 12–17. [Google Scholar]
- Akiyama, T.; Shiraaishi, M.; Yamamoto, T.; Unuma, T. Effect of dietary tryptophan on maturation of ayu Plecoglossus altivelis. Fish. Sci. 1996, 62, 776–782. [Google Scholar] [CrossRef]
- Comai, S.; Bertazzo, A.; Brughera, M.; Crotti, S. Tryptophan in health and disease. Adv. Clin. Chem. 2020, 95, 165–218. [Google Scholar] [CrossRef]
- Maler, L.; Ellis, W.G. Inter-male aggressive signals in weakly electric fish are modulated by monoamines. Behav. Brain Res. 1987, 25, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Munro, A.D. Effects of melatonin, serotonin, and naloxone on aggression in isolated cichlid fish (Aequiidens pulcher). J. Pineal Res. 1986, 3, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.F.; Liley, N.R.; Gorzalka, B.B. PCPA increases aggression in male firemouth Cichlids. Pharmacology 1996, 55, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Rawat, P.; Patel, A.B.; Jena, A. Dietary supplementation of L-tryptophan: Effect on growth and survival of Pabda, Ompok bimaculatus (Bloch) fry. J. Appl. Aquac. 2018, 31, 322–336. [Google Scholar] [CrossRef]
- Biswas, P.; Rawat, P.; Jena, A.K.; Patel, A.B.; Pandey, P.K. Effect of L-tryptophan on growth and survival of Pabda Fry, Ompok bimaculatus (Bloch, 1794). Fish. Technol. Soc. Fish. Technol. 2019, 56, 29–33. [Google Scholar]
- Álvarez-González, C.A.; Civera-Cerecedo, R.; Ortiz-Galindo, J.L.; Dumas, S.; Moreno-Legorreta, M.; Grayeb-Del, A.T. Effect of dietary protein level on growth and body composition of juvenile spotted sand bass, Paralabrax maculatofasciatus, fed practical diets. Aquaculture 2001, 194, 151–159. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2000. [Google Scholar]
- NOM-062-ZOO-1999, 2001. Norma Oficial Mexicana: Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. Available online: https://www.gob.mx/senasica/documentos/nom-062-zoo-1999 (accessed on 7 June 2022).
- Shuangyao, W.; Jiang, Z.; Mingguang, M.; Shoukang, M.; Yang, S.; Youzhen, S. Effects of seawater pH on survival, growth, energy budget and oxidative stress parameters of juvenile turbot scophthalmus maximus. Iran. J. Fish. Sci. 2018, 17, 675–689. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sarath, G.; de La Motte, R.S.; Wagner, F.W. Protease assay methods. In Proteolytic Enzymes: A Practical Approach; Beynon, R.J., Bonde, J.S., Eds.; Oxford University Press: Oxford, UK, 1989; pp. 25–54. [Google Scholar]
- Erlanger, B.F.; Kokowsky, N.; Cohen, W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys. 1961, 95, 271–278. [Google Scholar] [CrossRef]
- Maroux, S.; Louvard, D.; Barath, J. The aminopeptidase from hog intestinal brush border. Biochim. Biophys Acta Enzymol. 1973, 321, 282–295. [Google Scholar] [CrossRef]
- Versaw, W.; Cuppett, S.L.; Winters, D.D.; Williams, L.E. An improved colorimetric assay for bacterial lipase in nonfat dry milk. J. Food Sci. 1989, 54, 232–254. [Google Scholar] [CrossRef]
- Martínez-Burguete, T.; Peña-Marin, E.S.; García-Gasca, A.; Alvarez-González, C.A.; Llera-Herrera, R. Nutrigenomic marker discovery by de novo transcriptomic sequencing during early development of the tropical gar (Atractosteus tropicus). Aquac. Res. 2021, 52, 3829–3842. [Google Scholar] [CrossRef]
- Filby, A.L.; Paull, G.C.; Hickmore, T.F.A.; Tyler, C.R. Unravelling the neurophysiological basis of aggression in a fish model. BMC Genom. 2010, 11, 498. [Google Scholar] [CrossRef]
- de Abreu, M.S.; Giacomini, A.C.; Genario, R.; dos Santos, B.E.; da Rosa, L.G.; Demin, K.A.; Wappler-Guzzetta, E.A.; Kalueff, A.V. Neuropharmacology, pharmacogenetics and pharmacogenomics of aggression: The zebrafish model. Pharmacol. Res. 2019, 141, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Martínez, L.D.; Morales-Garcia, V.; Frias-Quintana, C.A.; Castillo-Collado, A.C.; Asencio-Alcudia, G.G.; Alvarez-Villagomez, C.S.; Peña-Marín, E.S.; Concha-Frias, B.; Alvarez-Gonzalez, C.A. Quality evaluation of reference gene expression on different tissues in adults of tropical gar Atractosteus tropicus. Pak. J. Zool. 2021, 54, 363–372. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔC(T) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Mai, K.; Xue, M.; He, G.; Xie, S.; Kaushik, S.J. Protein and amino acids. In Fish Nutrition; Academic Press: Cambridge, MA, USA, 2022; pp. 181–302. [Google Scholar]
- Walton, M.J.; Coloso, R.M.; Cowey, C.B.; Adron, J.W.; Knox, D. The effects of dietary tryptophan levels on growth and metabolism of rainbow trout (Salmo gairdneri). Br. J. Nutr. 1984, 51, 279. [Google Scholar] [CrossRef]
- Coloso, R.M.; Murillo-Gurrea, D.P.; Borlongan, I.G.; Catacutan, M.R. Tryptophan requirement of juvenile Asian sea bass Lates calcarifer. J. Appl. Ichthyol. 2004, 20, 43–47. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Mirghaed, A.T.; Mazandarani, M.; Zoheiri, F. Serum cortisol, glucose, thyroid hormones’ and non-specific immune responses of Persian sturgeon, Acipenser persicus to exogenous tryptophan and acute stress. Aquaculture 2016, 462, 17–23. [Google Scholar] [CrossRef]
- Hseu, J.; Lu, F.; Su, H.; Wang, L.; Tsai, C.; Hwang, P. Effect of exogenous tryptophan on cannibalism, survival and growth in juvenile grouper, Epinephelus coioides. Aquaculture 2003, 218, 251–263. [Google Scholar] [CrossRef]
- Jiang, W.D.; Wen, H.L.; Liu, Y.; Jiang, J.; Kuang, S.Y.; Wu, P.; Zhao, J.; Tang, L.; Tang, W.N.; Zhang, Y.A.; et al. The tight junction protein transcript abundance changes and oxidative damage by tryptophan deficiency or excess are related to the modulation of the signalling molecules, NF-κB p65, TOR, caspase-(3,8,9) and Nrf2 mRNA levels, in the gill of young grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2015, 46, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Wendelaar-Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Chacoff, L.; Nualart, D.; Vargas-Lagos, C.; Dann, F.; Muñoz, J.L.P.; Pontigo, J.P. Tryptophan and cortisol modulate the kynurenine and serotonin transcriptional pathway in the kidney of Oncorhynchus kisutch. Animals 2023, 13, 3562. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.C.; Shiu, Y.L.; Lin, H.L.; Liu, C.H. Immune response and disease resistance of barramundi, Lates calcarifer (Bloch), offered diets supplemented with replete levels of tryptophan or whey. J. World Aquac. Soc. 2017, 49, 127–140. [Google Scholar] [CrossRef]
- Król, J.; Zakęś, Z. Effect of dietary l-tryptophan on cannibalism, survival and growth in pikeperch Sander lucioperca (L.) post-larvae. Aquac. Int. 2015, 24, 441–451. [Google Scholar] [CrossRef]
- Ahmed, I. Dietary amino acid l-tryptophan requirement of fingerling Indian catfish, Heteropneustes fossilis (Bloch), estimated by growth and haemato-biochemical parameters. Fish Physiol. Biochem. 2012, 38, 1195–1209. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Khan, M.A. Dietary tryptophan requirement of fingerling Indian major carp, Cirrhinus mrigala (Hamilton). Aquac. Res. 2005, 36, 687–695. [Google Scholar] [CrossRef]
- Baras, E.; Silva Del Aguila, D.V.; Montalvan Naranjos, G.V.; Dugué, R.; Koo, F.C.; Duponchelle, F.; Renno, J.F.; Garcia-Dávila, C.; Nuñez, J. How many meals a day to minimize cannibalism when rearing larvae of the Amazonian catfish Pseudoplatystoma punctifer? The cannibal’s point of view. Aquat. Living Resour. 2011, 24, 379–390. [Google Scholar] [CrossRef]
- Kumar, P.; Kailasam, M.; Sethi, S.N.; Sukumaran, K.; Biswas, G.; Subburaj, R.; Thiagarajan, G.; Ghoshal, T.; Vijayan, K. Effect of dietary L-tryptophan on cannibalism, growth and survival of Asian seabass, Lates calcarifer (Bloch, 1790) fry. Indian J. Fish. 2017, 64, 28–32. [Google Scholar] [CrossRef]
- Baras, E.; Dugué, R.; Legendre, M. Do cannibalistic fish forage optimally? An experimental study of prey size preference, bioenergetics of cannibalism and their ontogenetic variations in the African catfish Heterobranchus longifilis. Aquat. Living Resour. 2014, 27, 51–62. [Google Scholar] [CrossRef]
- Folkvord, A. Ontogeny of cannibalism in larval and juvenile fishes with special emphasis on Atlantic cod. In Early Life History and Recruitment in Fish Populations; Chambers, R.C., Edward, A., Trippel, E.A., Eds.; Chapman & Hall: London, UK, 1992. [Google Scholar] [CrossRef]
- Hseu, J.R.; Hwang, P.P.; Ting, Y.Y. Morphometric model and laboratory analysis of intracohort cannibalism in giant grouper Epinephelus lanceolatus fry. Fish. Sci. 2004, 70, 482–486. [Google Scholar] [CrossRef]
- Xi, D.; Zhang, X.; Lü, H.; Zhang, Z. Cannibalism in juvenile black rockfish, Sebastes schlegelii (Hilgendorf, 1880), reared under controlled conditions. Aquaculture 2017, 479, 682–689. [Google Scholar] [CrossRef]
- Höglund, E.; Bakke, M.J.; ØVerli, Y.; Winberg, S.; Nilsson, G.E. Suppression of aggressive behaviour in juvenile Atlantic cod (Gadus morhua) by l-tryptophan supplementation. Aquaculture 2005, 249, 525–531. [Google Scholar] [CrossRef]
- Lepage, O.; Vílchez, I.M.; Pottinger, T.G.; Winberg, S. Time-course of the effect of dietaryl-tryptophan on plasma cortisol levels in rainbow trout Oncorhynchus mykiss. J. Exp. Biol. 2003, 206, 3589–3599. [Google Scholar] [CrossRef]
- Lepage, O.; Larson, E.T.; Mayer, I.; Winberg, S. Serotonin, but not melatonin, plays a role in shaping dominant–subordinate relationships and aggression in rainbow trout. Horm. Behav. 2005, 48, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Sahu, S.; Ngasotter, S.; Mog, M.; Tesia, S.; Sharma, S.; Dayakar, B.; Waikhom, D. A review on physiological, behavioral and metabolic role of dietary tryptophan in fish. Int. J. Chem. Stud. 2020, 8, 2411–2417. [Google Scholar] [CrossRef]
- Höglund, E.; ØVerli, Y.; Winberg, S. Tryptophan metabolic pathways and brain serotonergic activity: A comparative review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef]
- Winberg, S.; Nilsson, G.E. Roles of brain monoamine neurotransmitters in agonistic behaviour and stress reactions, with particular reference to fish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 1993, 106, 597–614. [Google Scholar] [CrossRef]
- Sepúlveda-Quiroz, C.; Alvarez-Villagomez, C.; Mendoza-Porras, O.; Peña-Marín, E.; Maytorena-Verdugo, C.; Pérez-Jiménez, G.; Jesus-Contreras, R.; Álvarez-González, C.; Martínez-García, R. Attack behavior leading cannibalism in tropical gar (Atractosteus tropicus) larvae under different tank colors and shelter type. Aquaculture 2023, 563, 738991. [Google Scholar] [CrossRef]
- Frías-Quintana, C.A.; Márquez-Couturier, G.; Álvarez-González, C.A.; Tovar-Ramírez, D.; Nolasco-Soria, H.; Galaviz-Espinosa, M.A.; Gisbert, E. Development of digestive tract and enzyme activities during the early ontogeny of the tropical gar Atractosteus tropicus. Fish Physiol. Biochem. 2015, 41, 1075–1091. [Google Scholar] [CrossRef] [PubMed]
- Svatos, A. The effect of amino acids, sugars and fats on activation of pancreatic enzymes. Cas. Lek. Cesk. 1994, 133, 140–143. [Google Scholar] [PubMed]
- Bone, Q.; Moores, R.H. Biology of Fishes, 3rd ed.; Taylor & Francis: London, UK, 2008. [Google Scholar] [CrossRef]
- Hakim, Y.; Uni, Z.; Hulata, G.; Harpaz, S. Relationship between intestinal brush border enzymatic activity and growth rate in tilapias fed diets containing 30% or 48% protein. Aquaculture 2006, 257, 420–428. [Google Scholar] [CrossRef]
- Moyano, F.J.; Díaz, M.; Alarcón, F.J.; Sarasquete, M.C. Characterization of digestive enzyme activity during larval development of gilthead seabream (Sparus aurata). Fish Physiol. Biochem. 1996, 15, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Cahu, C.; Zambonino Infante, J. Substitution of live food by formulated diets in marine fish larvae. Aquaculture 2001, 200, 161–180. [Google Scholar] [CrossRef]
- Tang, L.; Feng, L.; Sun, C.Y.; Chen, G.F.; Jiang, W.D.; Hu, K.; Liu, Y.; Jiang, J.; Li, S.H.; Kuang, S.Y.; et al. Effect of tryptophan on growth, intestinal enzyme activities and TOR gene expression in juvenile Jian carp (Cyprinus carpio var. Jian): Studies in vivo and in vitro. Aquaculture 2013, 412–413, 23–33. [Google Scholar] [CrossRef]
- Aldman, G.; Grove, D.; Holmgren, S. Duodenal acidification and intra-arterial injection of CCK8 increase gallbladder motility in the rainbow trout, Oncorhynchus mykiss. Gen. Comp. Endocrinol. 1992, 86, 20–25. [Google Scholar] [CrossRef]
- Jaworek, J. Ghrelin and metonin in the regulation of pancreatic exocrine secretion and maintaining of integrity. J. Physiol. Pharmacol. 2006, 57, 83–96. [Google Scholar]
- Zambonino-Infante, J.; Cahu, C. Ontogeny of the gastrointestinal tract of marine fish larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 477–487. [Google Scholar] [CrossRef]
- Pianesso, D.; Radünz Neto, J.; da Silva, L.; Goulart, F.; Adorian, T.; Mombach, P.; Loureiro, B.; Dalcin, M.; Rotili, D.; Lazzari, R. Determination of tryptophan requirements for juvenile silver catfish (Rhamdia quelen) and its effects on growth performance, plasma and hepatic metabolites and digestive enzymes activity. Anim. Feed Sci. Technol. 2015, 210, 172–183. [Google Scholar] [CrossRef]
- Mardones, O.; Oyarzún-Salazar, R.; Labbé, B.; Míguez, J.M.; Vargas-Chacoff, L.; Muñoz, J. Intestinal variation of serotonin, melatonin, and digestive enzymes activities along food passage time through GIT in Salmo salar fed with supplemented diets with tryptophan and melatonin. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 266, 111159. [Google Scholar] [CrossRef] [PubMed]
- Mardones, O.; Devia, E.; Labbé, B.; Oyarzún-Salazar, R.; Vargas-Chacoff, L.; Muñoz, J. Effect of L -tryptophan and melatonin supplementation on the serotonin gastrointestinal content and digestive enzymatic activity for Salmo salar and Oncorhynchus kisutch. Aquaculture 2018, 482, 203–210. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Lee, H.J.; Macbeth, A.H.; Young, W.S. Vasopressin: Behavioral roles of an “original” neuropeptide. Prog. Neurobiol. 2008, 84, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Conrad, J.L.; Weinersmith, K.L.; Brodin, T.; Saltz, J.B.; Sih, A. Behavioural syndromes in fishes: A review with implications for ecology and fisheries management. J. Fish Biol. 2011, 78, 395–435. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.E.; Watt, M.J.; Forster, G.L.; Øverli, Y.; Bockholt, C.; Renner, K.J.; Summers, C.H. Corticotropin releasing factor induces anxiogenic locomotion in trout and alters serotonergic and dopaminergic activity. Horm. Behav. 2007, 52, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.; Korzan, W.; Bockholt, C.; Watt, M.; Forster, G.; Renner, K.; Summers, C. Corticotropin releasing factor influences aggression and monoamines: Modulation of attacks and retreats. Neuroscience 2009, 158, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Loveland, J.L.; Uy, N.; Maruska, K.P.; Carpenter, R.E.; Fernald, R.D. Social status differences regulate the serotonergic system of a cichlid fish, Astatotilapia burtoni. J. Exp. Biol. 2014, 217, 2680–2690. [Google Scholar] [CrossRef]
- Clotfelter, E.D.; O’Hare, E.P.; McNitt, M.M.; Carpenter, R.E.; Summers, C.H. Serotonin decreases aggression via 5-HT1A receptors in the fighting fish Betta splendens. Pharmacol. Biochem. Behav. 2007, 87, 222–231. [Google Scholar] [CrossRef]
- Filby, A.L.; Paull, G.C.; Searle, F.; Ortiz-Zarragoitia, M.; Tyler, C.R. Environmental estrogen-induced alterations of male aggression and dominance hierarchies in fish: A mechanistic analysis. Environ. Sci. Technol. 2012, 46, 3472–3479. [Google Scholar] [CrossRef]
- Hecht, T.; Appelbaum, S. Observations on intraspecific aggression and coeval sibling cannibalism by larval and juvenile Claias gariepinus (Clariidae: Pisces) under controlled conditions. J. Zool. 1988, 214, 21–44. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2014, 17, 1–30. [Google Scholar] [CrossRef]
- Qin, J.G.; Mittiga, L.; Ottolenghi, F. Cannibalism reduction in juvenile Barramundi Lates calcarifer by providing refuges and low light. J. World Aquac. Soc. 2004, 35, 113–118. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, X.; Wang, Y.; Zhang, X. Effects of environmental enrichment on growth performance, aggressive behavior and stress-induced changes in cortisol release and neurogenesis of black rockfish Sebastes schlegelii. Aquaculture 2020, 528, 735483. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trp (g/kg) | ||||
|---|---|---|---|---|
| Ingredients | CD | 10 | 20 | 30 |
| Fish meal a | 305.4 | 305.4 | 305.4 | 305.4 |
| Renderer meal a | 300.0 | 300.0 | 300.0 | 300.0 |
| Soy meal a | 150.0 | 150.0 | 150.0 | 150.0 |
| Corn starch b | 67.1 | 67.1 | 67.1 | 67.1 |
| Oil soy c | 116.5 | 116.5 | 116.5 | 116.5 |
| Cellulose d | 30.0 | 20.0 | 10.0 | 0.0 |
| Tryptophan d | 0.0 | 10.0 | 20.0 | 30.0 |
| Premix vit-min e | 15.0 | 15.0 | 15.0 | 15.0 |
| Grenetine f | 10.0 | 10.0 | 10.0 | 10.0 |
| Vit C g | 5.0 | 5.0 | 5.0 | 5.0 |
| Vit E h | 1.0 | 1.0 | 1.0 | 1.0 |
| Proximate composition (g/100 g dry matter), except energy | ||||
| Energy (kJ/g) | 17.91 | 17.53 | 17.39 | 17.48 |
| Protein | 44.00 | 45.00 | 46.00 | 47.00 |
| Ether extract | 16.38 | 16.59 | 16.41 | 16.48 |
| Fibre | 1.05 | 1.12 | 1.07 | 1.01 |
| Ash | 13.43 | 13.23 | 13.02 | 12.93 |
| NFE 1 | 25.14 | 24.06 | 23.5 | 22.58 |
| Trp (g/kg) | ||||
|---|---|---|---|---|
| Fatty Acids (%) | CD | 10 | 20 | 30 |
| C13:0 | 7.0 | 10.2 | 9.7 | 7.9 |
| C14:0 | 1.2 | 1.3 | 1.2 | 1.2 |
| C16:0 | 16.7 | 17.0 | 17.0 | 17.4 |
| C17:0 | ND | ND | ND | ND |
| C18:0 | 5.8 | 6.0 | 6.0 | 6.1 |
| C23:0 | ND | ND | ND | ND |
| ƩSFA | 30.7 | 34.4 | 33.9 | 32.7 |
| C16:1n7 | 2.0 | 2.1 | 2.1 | 2.2 |
| C18:1n9 | 21.0 | 20.4 | 20.5 | 21.1 |
| C18:1n7 | 1.6 | 1.8 | 1.8 | 1.8 |
| ƩMUFAS | 24.6 | 24.3 | 24.5 | 25.1 |
| C18:2n6 | 34.9 | 32.0 | 32.3 | 32.6 |
| C18:3n3 | 0.3 | 4.3 | 4.3 | 4.3 |
| C18:4n3 | 4.7 | ND | ND | ND |
| C20:3n3 | 0.5 | 0.6 | 0.7 | 0.7 |
| C20:4n6 | 0.3 | ND | ND | ND |
| C20:5n3 | 1.6 | 1.7 | 1.8 | 1.7 |
| C22:5n3 | 0.4 | 0.4 | 0.4 | 0.4 |
| C22:6n3 | 1.9 | 2.2 | 2.2 | 2.4 |
| ƩPUFAS | 44.6 | 41.3 | 41.6 | 42.2 |
| NID | 0.0 | 0.0 | 0.0 | 0.0 |
| 100.0 | 100.0 | 100.0 | 100.0 | |
| Trp (g/kg) | ||||
|---|---|---|---|---|
| Amino Acid | CD | 10 | 20 | 30 |
| Essential amino acids | ||||
| HIS | 1.1 | 1.0 | 1.2 | 1.0 |
| ARG | 5.2 | 5.6 | 5.4 | 5.3 |
| THR | 1.7 | 1.6 | 1.7 | 1.7 |
| VAL | 1.7 | 1.6 | 1.6 | 1.6 |
| MET | 0.5 | 0.5 | 0.2 | 0.2 |
| LYS | 5.7 | 5.0 | 5.3 | 5.5 |
| ILE | 1.3 | 1.2 | 1.3 | 1.3 |
| LEU | 3.5 | 3.2 | 3.5 | 3.5 |
| PHE | 1.4 | 1.2 | 1.4 | 1.4 |
| subtotal | 22.1 | 20.9 | 21.5 | 21.7 |
| Non-essential amino acids | ||||
| ASP | 2.4 | 2.2 | 2.4 | 2.4 |
| SER | 2.3 | 2.2 | 2.4 | 2.3 |
| GLU | 5.8 | 5.7 | 6.0 | 6.0 |
| GLY | 7.0 | 8.2 | 7.1 | 6.9 |
| ALA | 3.4 | 3.7 | 3.6 | 3.5 |
| TYR | 1.5 | 1.3 | 1.4 | 1.5 |
| subtotal | 22.3 | 23.4 | 22.9 | 22.7 |
| Others | ||||
| TAU | 0.6 | 0.7 | 0.6 | 0.6 |
| Total | 45.0 | 45.0 | 45.0 | 45.0 |
| Tryptophan (mg/g) | 10.60 | 19.74 | 32.42 | 44.21 |
| Protein | Gen | Primers (5′-3′) | Alignment Temperature (°C) |
|---|---|---|---|
| Somatostatin receptor 1 | sstr1 | FW: CCTCAGCATTGACCGCTACA RV: AATACCGCCATCCACTGACG | 60 |
| Tyrosine hydroxylase | th | FW: GGACCAGATGTACCAGCCAG RV: GCAGTTCATCCCTCGCAGAT | 59 |
| Histidine decarboxylase | hdc | FW: GCATTTCGACTGCACTGCTT RV: CTTCGGCTGAGTGGGATCTG | 59 |
| Corticotropin-releasing hormone | crh | FW: AACGTCAACAGGGCTTTCCA RV: TCTTCCCGTCAGGTCTTCCA | 60 |
| 5-hydroxytryptamine (serotonin) receptor 1A, G protein-coupled | htr1a | FW: AAGCGCAGTGTGGAACCTAA RV: GCTGTCGGGGTATTAGGCAG | 60 |
| Gonadotropin-releasing hormone 1 | gnrh1 | FW: AGTCAGCACTGGTCATACGG RV: CTCACCTCCTCCGCAATGTC | 59 |
| Dopamine receptor D1 | drd1 | FW: TTTTTGGCCCTTTGGCTCATT RV: AAGTTCAAAATGGAGGCTGTGG | 59 |
| Arginine vasopressin-induced 1 | avpi1 | FW: AGGGAGGACCACTGAAGATGA RV: CCAGCAGAGGACAAGTCTGC | 60 |
| Tryptophan hydroxylase 1 | tph1 | FW: CCCCCGTATCGAGTTCACAG RV: AGGGGCAGGTTCTTGAGGTA | 60 |
| Trp (g/kg) | ||||
|---|---|---|---|---|
| CD | 10 | 20 | 30 | |
| Initial weight (g) | 0.018 ± 0.001 | 0.018 ± 0.001 | 0.018 ± 0.001 | 0.018 ± 0.001 |
| Final weight (g) | 0.022 ± 0.0003 | 0.023 ± 0.001 | 0.023 ± 0.001 | 0.023 ± 0.001 |
| Initial total length (cm) | 1.28 ± 0.09 | 1.28 ± 0.09 | 1.28 ± 0.09 | 1.28 ± 0.09 |
| Final total length (cm) | 1.78 ± 0.05 | 1.81 ± 0.02 | 1.83 ± 0.03 | 1.80 ± 0.03 |
| S (%) | 28.25 ± 1.76 | 34.75 ± 5.30 | 28.66 ± 1.04 | 29.75 ± 1.76 |
| FI (g/d) | 0.031 ± 0.001 | 0.031 ± 0.003 | 0.031 ± 0.003 | 0.031 ± 0.001 |
| AWG (g/fish) | 0.004 ± 0.0003 | 0.005 ± 0.0008 | 0.005 ± 0.001 | 0.005 ± 0.00004 |
| SGR (%/d) | 5.07 ± 0.52 | 5.35 ± 0.29 | 5.53 ± 0.38 | 5.24 ± 0.39 |
| FCR | 7.23 ± 0.85 | 6.68 ± 0.53 | 5.77 ± 0.46 | 5.64 ± 0.02 |
| PER | 0.13 ± 0.01 | 0.15 ± 0.1 | 0.15 ± 0.04 | 0.15 ± 0.04 |
| K | 0.39 ± 0.02 | 0.39± 0.01 | 0.37± 0.01 | 0.38± 0.02 |
| CV (%) | 6.43 ± 2.98 | 8.63 ± 2.23 | 8.20 ± 3.05 | 6.95 ± 0.05 |
| SH | 0.71 ± 0.33 | 0.96 ± 0.25 | 0.91 ± 0.34 | 0.77 ± 0.006 |
| CD | 10 g/kg Trp | |
|---|---|---|
| Initial weight (g) | 0.049 ± 0.004 | 0.048 ± 0.006 |
| Final weight (g) | 0.179 ± 0.03 * | 0.080 ± 0.02 |
| Initial total length (cm) | 2.16 ± 0.11 | 2.19 ± 0.08 |
| Final total length (cm) | 3.68 ± 0.34 * | 2.72 ± 0.18 |
| S (%) | 23.33 ± 5.77 | 75.0 ± 7.07 * |
| FI (g/d) | 0.72 ± 0.07 | 0.76 ± 0.08 |
| AWG (g/fish) | 0.13 ± 0.03 * | 0.03 ± 0.01 |
| SGR (%/d) | 14.62 ± 2.87 * | 5.81 ± 3.18 |
| FCR | 5.66 ± 0.62 | 12.43 ± 1.60 * |
| PER | 0.17 ± 0.01 * | 0.05 ± 0.03 |
| K | 0.35 ± 0.02 | 0.39 ± 0.01 |
| CV (%) | 55.64 ± 6.84 * | 26.74 ± 14.20 |
| SH | 6.07 ± 0.75 * | 1.90 ± 1.02 |
| Activities | Trp (g/kg) | |||||
|---|---|---|---|---|---|---|
| First stage | (u/mg Protein) | CD | 10 | 20 | 30 | |
| Acid protease | 12.233 ±0.924 a | 2.505 ± 2.272 b | 3.546 ± 1.207 b | 7.645 ± 0.792 a,b | ||
| Alkaline protease | 17.374 ± 1.550 a | 8.160 ± 2.257 b | 7.388 ± 0.852 b | 7.784 ± 0.859 b | ||
| Trypsin | 1.334 ± 0.088 a | 0.682 ± 0.067 a,b | 0.640 ± 0.018 b | 0.645 ± 0.236 a,b | ||
| Leucine aminopeptidase | 0.366 ± 0.014 | 0.290 ± 0.007 | 0.317 ± 0.057 | 0.337 ± 0.107 | ||
| Lipase | 1.711 ± 0.654 b | 4.529 ± 0.318 a | 4.514 ± 0.099 a | 2.413 ± 0.079 b | ||
| CD | 10 | |||||
| Second stage | Acid protease | 20.492 ± 4.162 | 35.869 ± 3.670 * (0.033) | |||
| Alkaline protease | 7.043 ± 0.835 | 14.395 ± 2.342 * (0.028) | ||||
| Trypsin | 0.755 ± 0.079 | 1.032 ± 0.086 * (0.0151) | ||||
| Leucine aminopeptidase | 0.432 ± 0.017 | 1.333 ± 0.203 * (0.0167) | ||||
| Lipase | 3.178 ± 0.640 | 3.072 ± 0.400 | ||||
| CD | 10 | 20 | 30 | |||
| Cannibals vs. Non-cannibals | Acid protease | C | 28.261 ± 0.028 b,* | 43.110 ± 0.045 a,* | 18.780 ± 0.102 d | 25.451 ± 0.017 c,* |
| N | 23.576 ± 0.062 b | 39.280 ± 0.049 a | 19.469 ± 0.031 d,* | 23.228 ± 0.020 d | ||
| Alkaline protease | C | 15.887 ± 0.016 a | 10.241 ± 0.218 c,* | 14.263 ± 0.035 b,* | 9.963 ± 0.072 c,* | |
| N | 18.021 ± 0.128 a,* | 9.722 ± 0.186 b | 8.393 ± 0.166 c | 9.762 ± 0.016 b | ||
| Trypsin | C | 1.919 ± 0.318 a,* | 1.491 ± 0.143 b,* | 1.203 ± 0.082 b,* | 1.154 ± 0.144 b,* | |
| N | 1.493 ± 0.232 a | 1.170 ± 0.078 b | 0.851 ± 0.087 c | 0.863 ± 0.069 c | ||
| Leucine aminopeptidase | C | 0.284 ± 0.044 c | 0.527 ± 0.079 a | 0.684 ± 0.164 a,b | 0.382 ± 0.051 b,c | |
| N | 0.184 ± 0.086 c | 0.494 ± 0.045 a,b | 0.514 ± 0.053 a | 0.379 ± 0.042 b | ||
| Lipase | C | 0.900 ± 0.011 b | 3.288 ± 0.038 a | 0.481 ± 0.025 c | 0.588 ± 0.032 c | |
| N | 3.324 ± 0.019 b,* | 3.889 ± 0.001 a,* | 1.759 ± 0.016 c,* | 1.502 ± 0.012 d,* | ||
| Lateral Attack | Frontal Attack | ||||||
|---|---|---|---|---|---|---|---|
| Escape | Head | Middle | Tail | Head | Middle | Total | |
| Without shelter | |||||||
| CD | 2 | 2 | 0 | 0 | 0 | 0 | 4 |
| 10 g/kg Trp | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| Total | 4 | 2 | 0 | 0 | 0 | 0 | 6 |
| Rocks | |||||||
| CD | 2 | 0 | 1 | 0 | 0 | 0 | 3 |
| 10 g/kg Trp | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 2 | 0 | 1 | 0 | 0 | 0 | 3 |
| Artificial Vegetation | |||||||
| CD | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| 10 g/kg Trp | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Total | 3 | 0 | 0 | 0 | 0 | 0 | 3 |
| SUMA total | 9 | 2 | 1 | 0 | 0 | 0 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepúlveda-Quiroz, C.A.; Pérez-Jiménez, G.M.; Asencio-Alcudia, G.G.; Mendoza-Porras, O.; Jiménez-Martínez, L.D.; Galaviz-Espinoza, M.A.; Tovar-Ramirez, D.; Martinez-Garcia, R.; Alvarez-Villagomez, C.S.; Alvarez-Gonzalez, C.A. Tryptophan Reduces Intracohort Cannibalism Behavior in Tropical Gar (Atractosteus tropicus) Larvae. Fishes 2024, 9, 40. https://doi.org/10.3390/fishes9010040
Sepúlveda-Quiroz CA, Pérez-Jiménez GM, Asencio-Alcudia GG, Mendoza-Porras O, Jiménez-Martínez LD, Galaviz-Espinoza MA, Tovar-Ramirez D, Martinez-Garcia R, Alvarez-Villagomez CS, Alvarez-Gonzalez CA. Tryptophan Reduces Intracohort Cannibalism Behavior in Tropical Gar (Atractosteus tropicus) Larvae. Fishes. 2024; 9(1):40. https://doi.org/10.3390/fishes9010040
Chicago/Turabian StyleSepúlveda-Quiroz, César Antonio, Graciela María Pérez-Jiménez, Gloria Gertrudis Asencio-Alcudia, Omar Mendoza-Porras, Luis Daniel Jiménez-Martínez, Mario A. Galaviz-Espinoza, Dariel Tovar-Ramirez, Rafael Martinez-Garcia, Carina Shianya Alvarez-Villagomez, and Carlos Alfonso Alvarez-Gonzalez. 2024. "Tryptophan Reduces Intracohort Cannibalism Behavior in Tropical Gar (Atractosteus tropicus) Larvae" Fishes 9, no. 1: 40. https://doi.org/10.3390/fishes9010040
APA StyleSepúlveda-Quiroz, C. A., Pérez-Jiménez, G. M., Asencio-Alcudia, G. G., Mendoza-Porras, O., Jiménez-Martínez, L. D., Galaviz-Espinoza, M. A., Tovar-Ramirez, D., Martinez-Garcia, R., Alvarez-Villagomez, C. S., & Alvarez-Gonzalez, C. A. (2024). Tryptophan Reduces Intracohort Cannibalism Behavior in Tropical Gar (Atractosteus tropicus) Larvae. Fishes, 9(1), 40. https://doi.org/10.3390/fishes9010040