
Sicklefin Chub (Macrhybopsis meeki) and Sturgeon Chub (M. gelida) Temporal and Spatial Patterns from Extant Population Monitoring and Habitat Data Spanning 23 Years
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
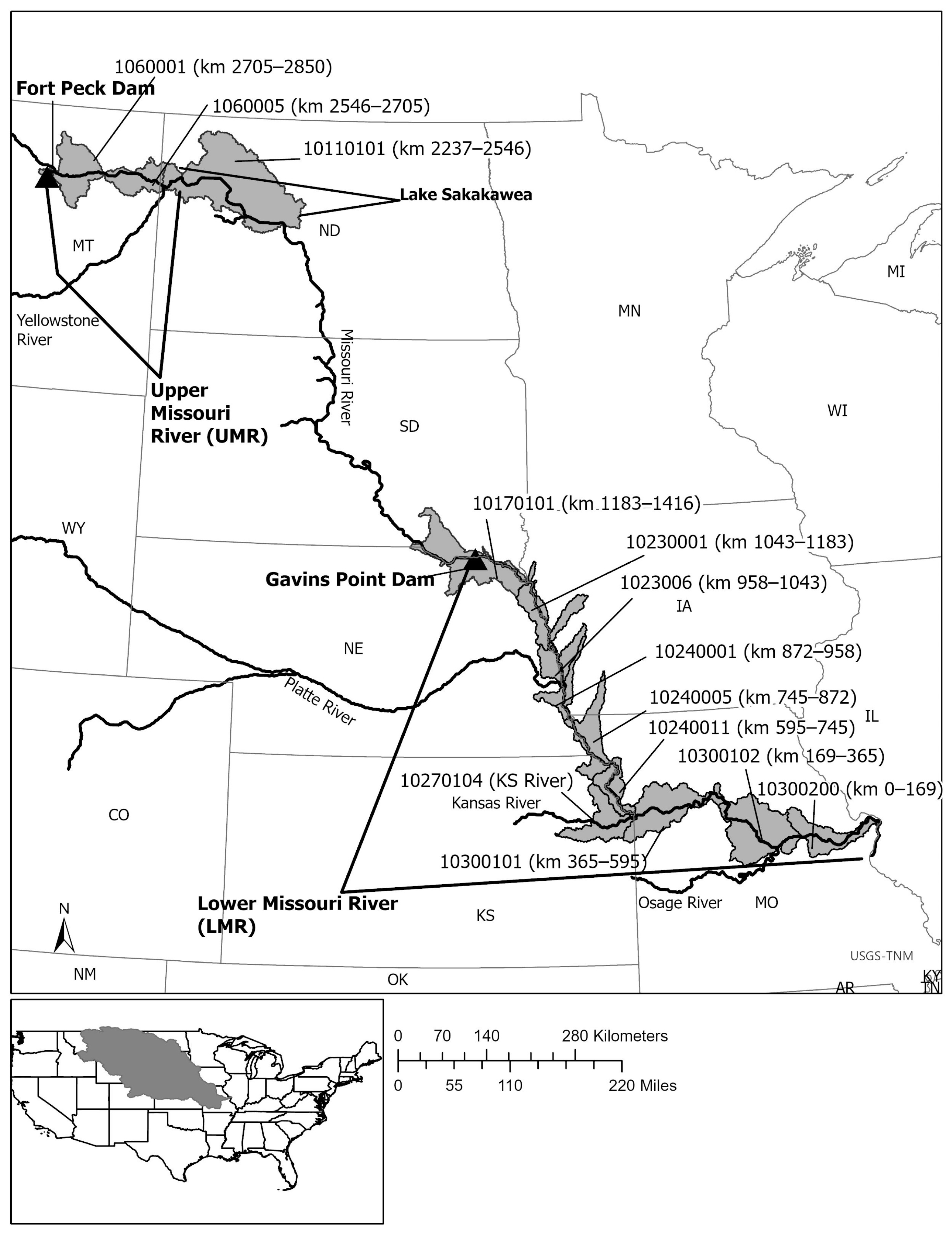
2.1. Extant Data and Study Area
2.2. Benthic Fish Species of Interest
2.3. Data Processing
2.4. Statistical Analyses
2.4.1. Occupancy Models
2.4.2. Post Hoc Missouri River Discharge Analyses
2.4.3. Observed Co-Occurrence and Simulations
3. Results
3.1. Two-Species Occupancy Models
3.1.1. Occupancy and Co-Occurrence
3.1.2. Model Performance and Detection
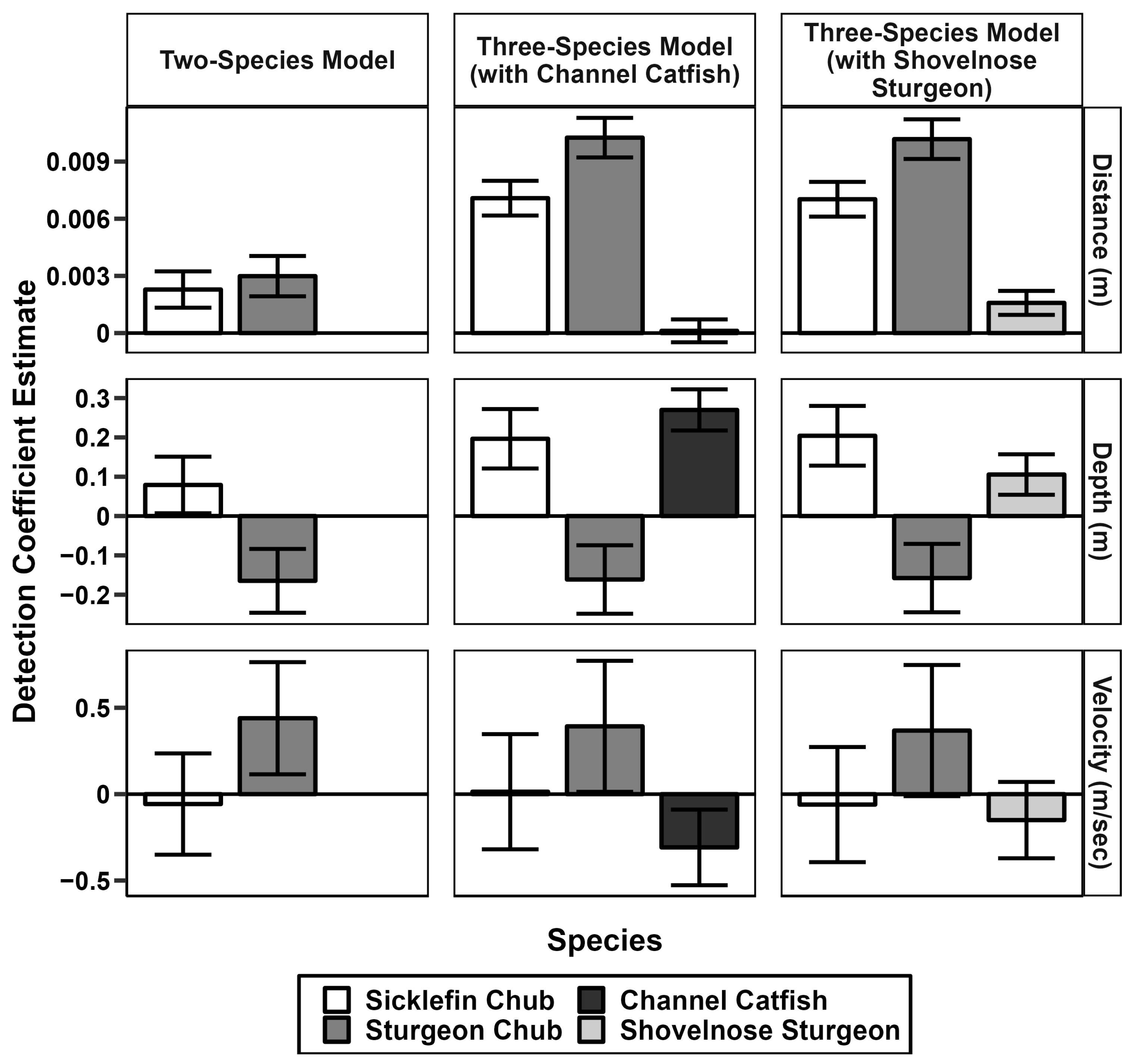
3.2. Three-Species Occupancy Models
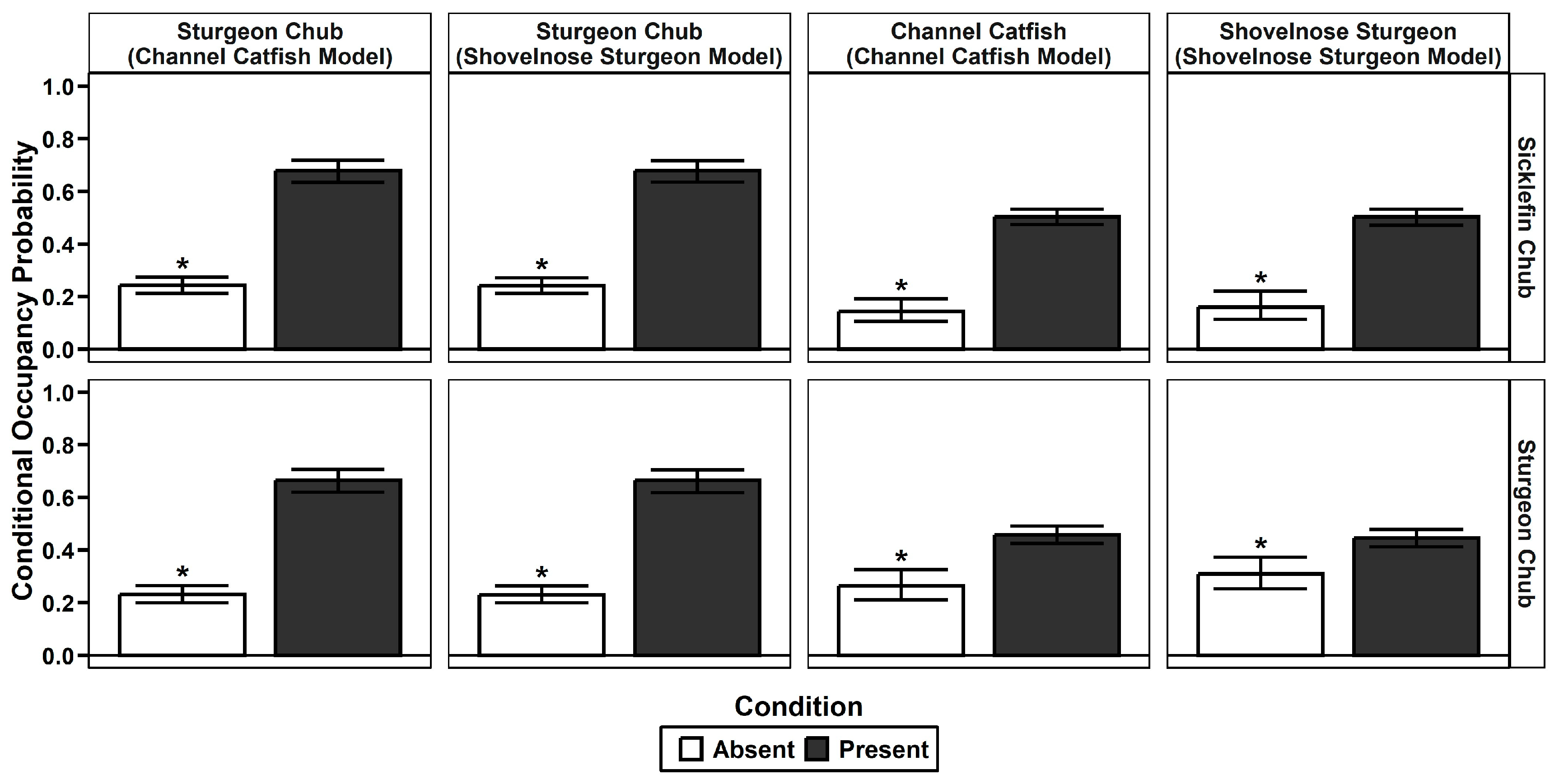
3.2.1. Occupancy and Co-Occurrence
3.2.2. AUC and Detection
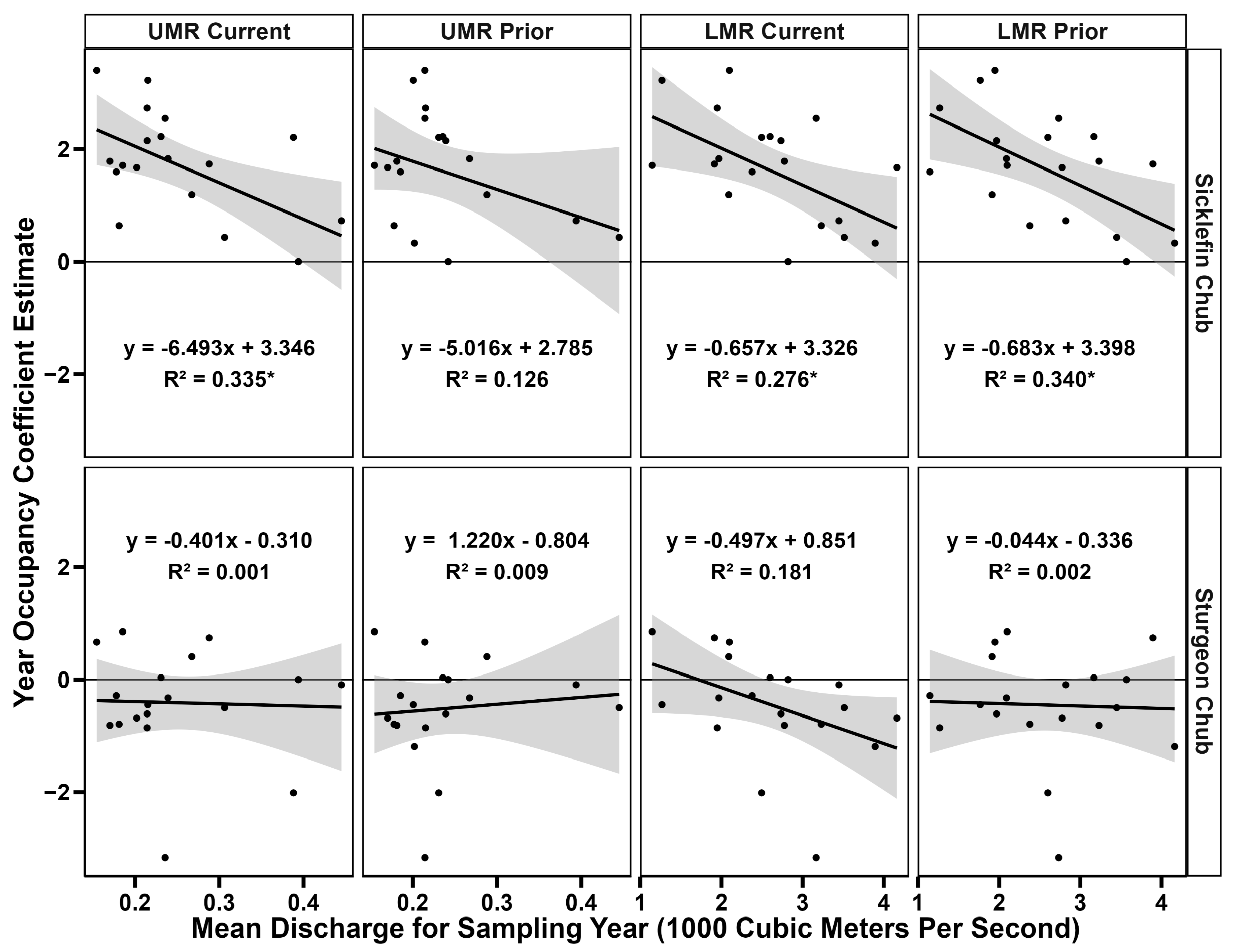
3.3. Missouri River Discharge Relationships
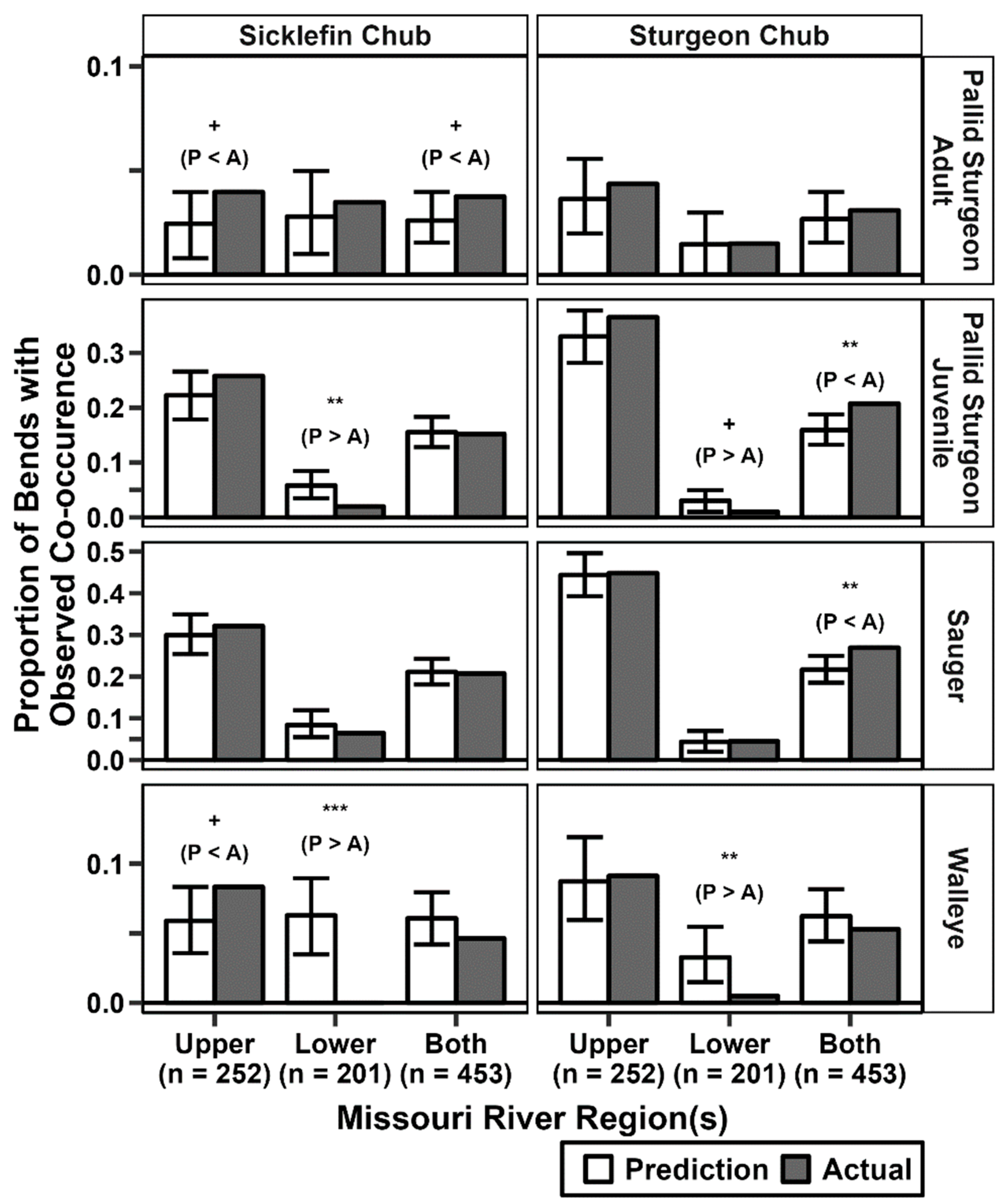
3.4. Observed Co-Occurrence and Simulations
4. Discussion
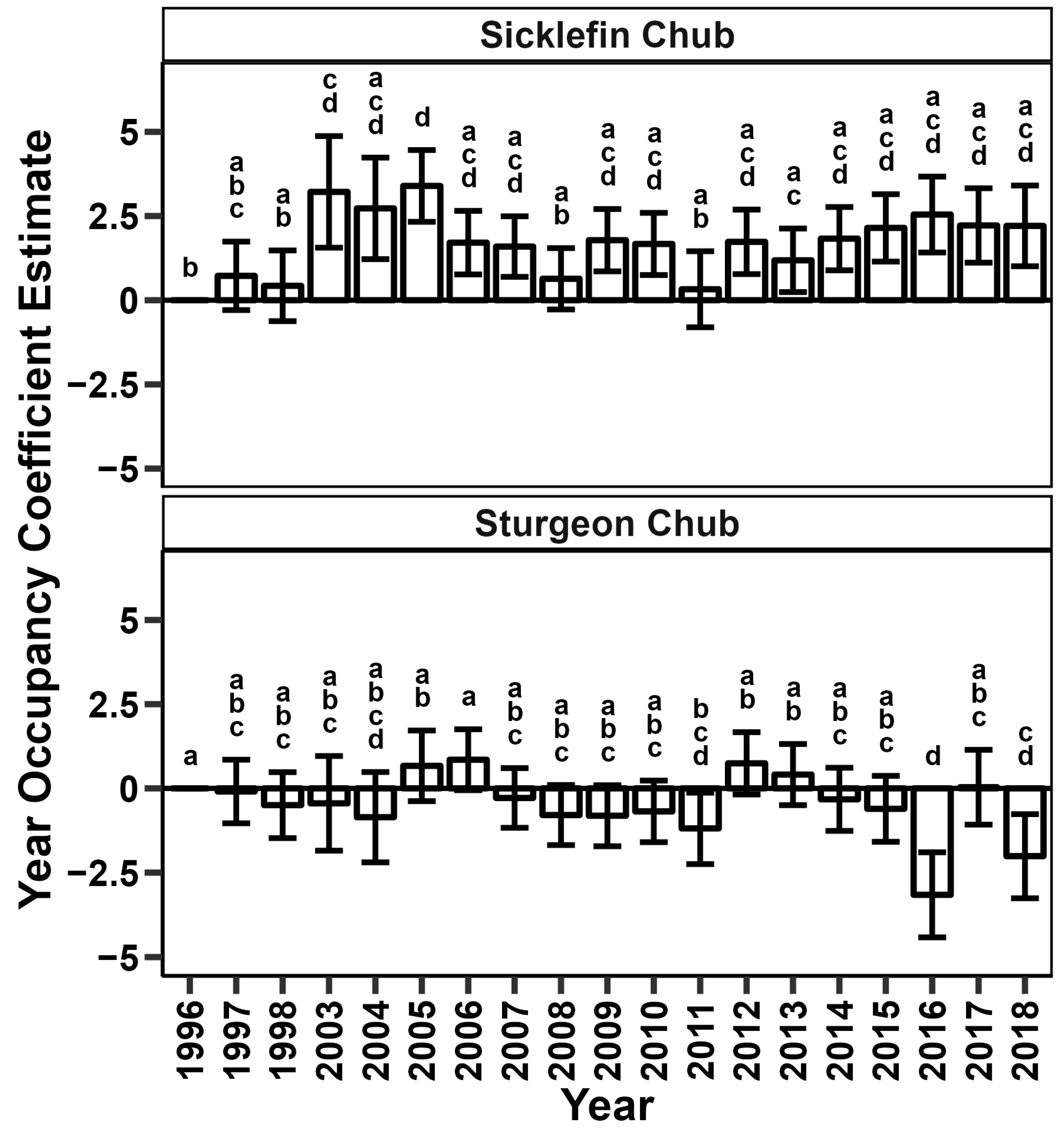
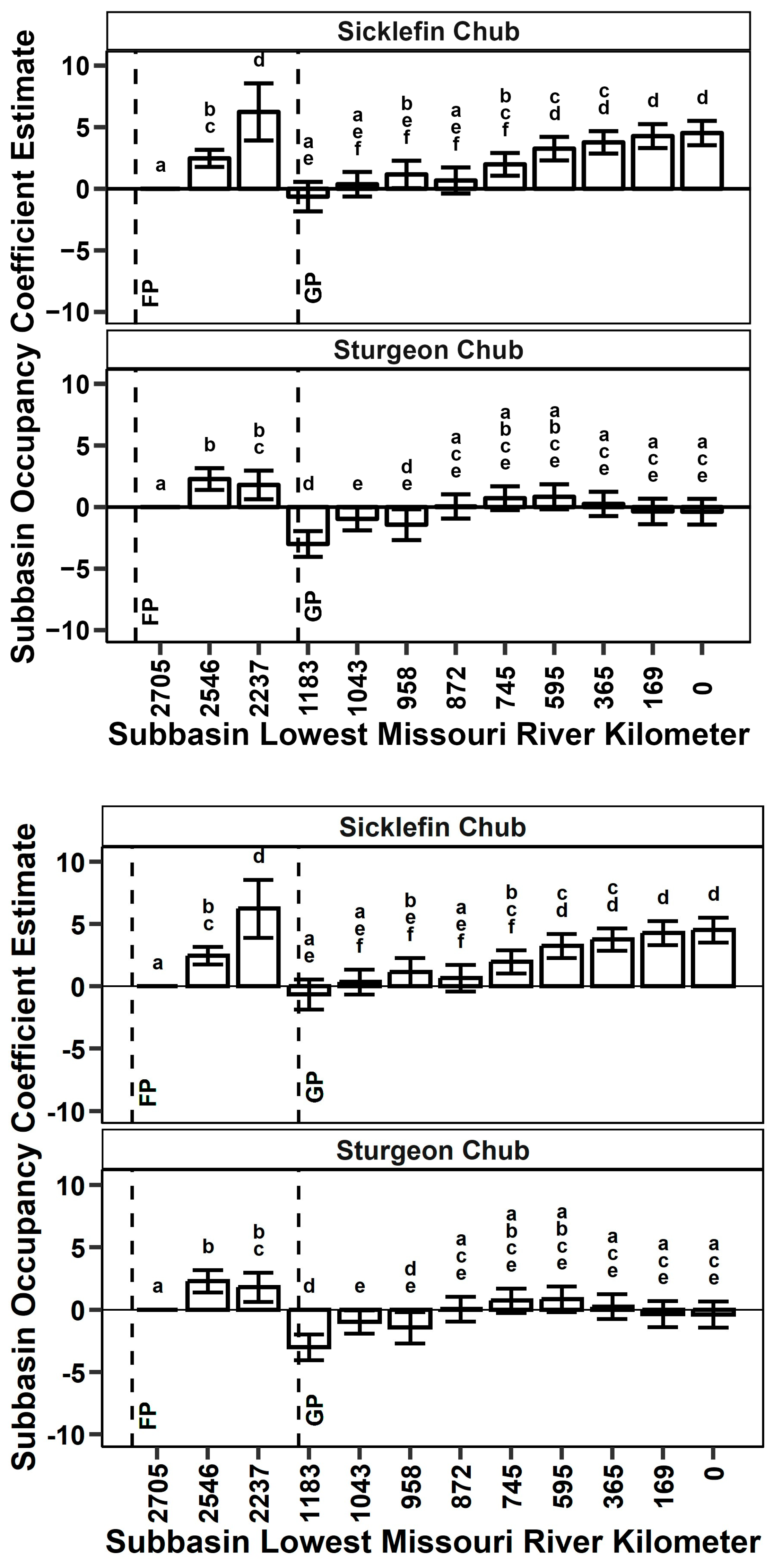
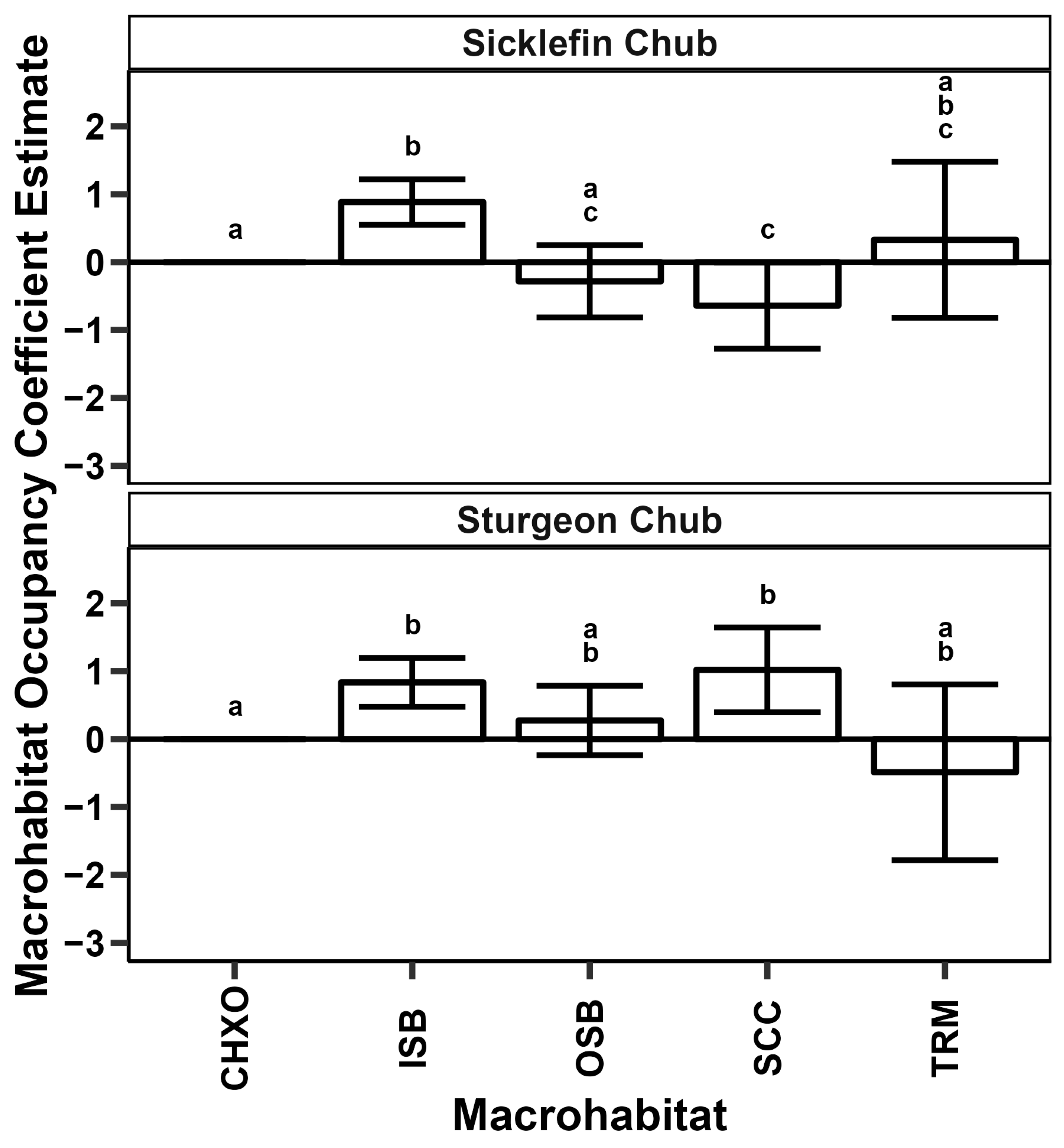
4.1. Occupancy Patterns
4.2. Possible Discharge Patterns
4.3. Co-Occurrence across Analyses
4.4. Other Potential Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Data Sources
Appendix B. Data Filtering and Processing
References
- Albers, J.L. Sicklefin chub: Macrhybopsis meeki (Jordan and Evermann 1896). In Kansas Fishes; University Press of Kansas: Lawrence, KS, USA, 2014; pp. 184–186. ISBN 978-0-7006-1961-0. [Google Scholar]
- Albers, J.L. Sturgeon chub: Macrhybopsis gelida (Girard 1856). In Kansas Fishes; University Press of Kansas: Lawerence, KS, USA, 2014; pp. 178–180. ISBN 978-0-7006-1961-0. [Google Scholar]
- Wildhaber, M.L.; Yang, W.H.; Arab, A. Population trends, bend use relative to available habitat and within-river-bend habitat use of eight indicator species of Missouri and Lower Kansas River benthic fishes: 15 years after baseline assessment. River Res. Appl. 2016, 32, 36–65. [Google Scholar] [CrossRef]
- William, B. Endangered and Threatened Wildlife and Plants; 12-Month Finding for a Petition to List the Sicklefin Chub (Macrhybopsis meeki) and the Sturgeon Chub (Macrhybopsis gelida) as Endangered; 01-9443; U.S. Fish and Wildlife Service: Washington, DC, USA, 2001; pp. 19910–19914. Available online: https://www.govinfo.gov/content/pkg/FR-2001-04-18/html/01-9443.htm (accessed on 2 November 2023).
- U.S. Fish and Wildlife Service. Species Status Assessment for the Sturgeon Chub (Macrhybopsis gelida) and Sicklefin Chub (Macrhybopsis meeki); U.S. Fish and Wildlife Service, Region 6: Lakewood, CO, USA, 2023. Available online: https://ecos.fws.gov/ServCat/DownloadFile/238509 (accessed on 29 September 2023).
- Gerrity, P.C.; Guy, C.S.; Gardner, W.M. Juvenile pallid sturgeon are piscivorous: A call for conserving native cyprinids. Trans. Am. Fish. Soc. 2006, 135, 604–609. [Google Scholar] [CrossRef]
- Huenemann, T. Central Lowlands and Interior Highlands Pallid Sturgeon Spawning and Stocking Summary 1992–2017; Middle Basin Pallid Sturgeon Workgroup: Lincoln, NE, USA, 2010; Available online: http://meridian.allenpress.com/jfwm/article-supplement/433030/pdf/10_3996022018-jfwm-013_s4 (accessed on 2 November 2023).
- Quist, M.C.; Boelter, A.M.; Lovato, J.M.; Korfanta, N.M.; Bergman, H.L.; Korschgen, C.E.; Galat, D.L.; Latka, D.C.; Krentz, S.; Oetker, M.; et al. Research and Assessment Needs for Pallid Sturgeon Recovery in the Missouri River: Final Report to the U.S. Geological Survey, U.S. Army Corps of Engineers, U.S. Fish and Wildlife Service, and U.S. Environmental Protection Agency; William D. Ruckelshaus Institute of Environment and Natural Resources, University of Wyoming: Laramie, WY, USA, 2004; p. 82. Available online: https://pallidsturgeon.org/wp-content/uploads/2018/01/28-Quist-et-al-2004.pdf (accessed on 19 December 2023).
- Rahel, F.J.; Thel, L.A. Sturgeon Chub (Macrhybopsis gelida): A Technical Conservation Assessment; USDA Forest Service, Rocky Mountain Region: Laramie, WY, USA, 2004; pp. 1–49. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5206786.pdf (accessed on 16 November 2023).
- Welker, T.L.; Drobish, M.R.; Williams, G.A. Pallid Sturgeon Population Assessment Project, Guiding Document, Volume 2.0; U.S. Army Corps of Engineers, Omaha District: Yankton, SD, USA, 2020. [Google Scholar]
- Ridenour, C.J.; Starostka, A.B.; Doyle, W.J.; Hill, T.D. Habitat used by chubs associated with channel modifying structures in a large regulated river: Implications for river modification. River Res. Appl. 2009, 25, 472–485. [Google Scholar] [CrossRef]
- Ridenour, C.J.; Wrasse, C.J.; Meyer, H.A.; Doyle, W.J.; Hill, T.D. Variation in Fish Assemblages among Four Habitat Complexes during a Summer Flood: Implications for Flow Refuge; U.S. Fish and Wildlife Service, Columbia Fish and Wildlife Conservation Office: Columbia, MO, USA, 2012. [Google Scholar]
- Wildhaber, M.L.; Gladish, D.W.; Arab, A. Distribution and habitat use of the Missouri River and Lower Yellowstone River benthic fishes from 1996 to 1998: A baseline for fish community recovery. River Res. Appl. 2012, 28, 1780–1803. [Google Scholar] [CrossRef]
- Steffensen, K.D.; Shuman, D.A.; Stukel, S. The status of fishes in the Missouri River, Nebraska: Shoal chub (Macrhybopsis hyostoma), sturgeon chub (M. gelida), sicklefin chub (M. meeki), silver chub (M. storeriana), Flathead chub (Platygobio gracilis), plains minnow (Hybognathus placitus), western silvery minnow (H. argyritis), and brassy minnow (H. hankinsoni). Trans. Neb. Acad. Sci. 2014, 34, 49–67. Available online: https://digitalcommons.unl.edu/tnas/470/ (accessed on 16 January 2024).
- Braaten, P.J.; Fuller, D.B.; Haddix, T.M.; Hunziker, J.R.; Colvin, M.E.; Holmquist, L.M.; Wilson, R.H. Catch rates for sturgeon chubs and sicklefin chubs in the Upper Missouri River 2004–2016 and correlations with biotic and abiotic variables. J. Fish Wildl. Manag. 2021, 12, 322–337. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Hines, J.E.; Knutson, M.G.; Franklin, A.B. Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology 2003, 84, 2200–2207. [Google Scholar] [CrossRef]
- Gu, W.; Swihart, R.K. Absent or undetected? Effects of non-detection of species occurrence on wildlife–habitat models. Biol. Conserv. 2004, 116, 195–203. [Google Scholar] [CrossRef]
- Royle, J.A.; Nichols, J.D.; Kéry, M. Modelling occurrence and abundance of species when detection is imperfect. Oikos 2005, 110, 353–359. [Google Scholar] [CrossRef]
- Specht, H.M.; Reich, H.T.; Iannarilli, F.; Edwards, M.R.; Stapleton, S.P.; Weegman, M.D.; Johnson, M.K.; Yohannes, B.J.; Arnold, T.W. Occupancy surveys with conditional replicates: An alternative sampling design for rare species. Methods Ecol. Evol. 2017, 8, 1725–1734. [Google Scholar] [CrossRef]
- Schloesser, J.T.; Paukert, C.P.; Doyle, W.J.; Hill, T.D.; Steffensen, K.D.; Travnichek, V.H. Heterogeneous detection probabilities for imperiled Missouri River Fishes: Implications for large-river monitoring programs. Endanger. Species Res. 2012, 16, 211–224. [Google Scholar] [CrossRef]
- Budy, P.; Conner, M.M.; Salant, N.L.; Macfarlane, W.W. An occupancy-based quantification of the highly imperiled status of desert fishes of the Southwestern United States. Conserv. Biol. 2015, 29, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.I.; Bailey, L.L.; Nichols, J.D. Investigating species co-occurrence patterns when species are detected imperfectly. J. Anim. Ecol. 2004, 73, 546–555. [Google Scholar] [CrossRef]
- Rota, C.T.; Ferreira, M.A.R.; Kays, R.W.; Forrester, T.D.; Kalies, E.L.; McShea, W.J.; Parsons, A.W.; Millspaugh, J.J. A multispecies occupancy model for two or more interacting species. Methods Ecol. Evol. 2016, 7, 1164–1173. [Google Scholar] [CrossRef]
- Lamothe, K.A.; Dextrase, A.J.; Drake, D.A.R. Characterizing species co-occurrence patterns of imperfectly detected stream fishes to inform species reintroduction efforts. Conserv. Biol. 2019, 33, 1392–1403. [Google Scholar] [CrossRef]
- Holt, A.R.; Gaston, K.J.; He, F.L. Occupancy-abundance relationships and spatial distribution: A review. Basic Appl. Ecol. 2002, 3, 1–13. [Google Scholar] [CrossRef]
- Berry, C.R.; Young, B.A. Introduction to the benthic fishes studies: Population structure and habitat. In Use of Benthic Fishes along the Missouri and Lower Yellowstone Rivers; United States Army Corps of Engineers, CENWO-PM-AE: Omaha, NE, USA, 2001; pp. 1–52. [Google Scholar]
- Welker, T.L.; Drobish, M.R.; Williams, G.A. Pallid Sturgeon Population Assessment Project, Guiding Document, Volume 1.8; U.S. Army Corps of Engineers, Omaha District: Yankton, SD, USA, 2017; Available online: https://meridian.allenpress.com/jfwm/article-supplement/433030/pdf/10_3996022018-jfwm-013_s14/ (accessed on 8 November 2023).
- Welker, T.L.; Drobish, M.R.; Williams, G.A. Pallid Sturgeon Population Assessment Project, Guiding Document, Volume 1.1; U.S. Army Corps of Engineers, Omaha District: Yankton, SD, USA, 2006. [Google Scholar]
- Wildhaber, M.L.; West, B.M.; Bennett, K.B.; Beaman, Z.D. Captures and Habitat Classification of Benthic Fishes along the Missouri and Lower Yellowstone Rivers, 1996–1998; U.S. Geological Survey Data Release: Columbia, MO, USA, 2023. [Google Scholar] [CrossRef]
- Jones, K.A.; Niknami, L.S.; Buto, S.G.; Decker, D. Federal standards and procedures for the national watershed boundary dataset (WBD). In Section A, Federal Standards, Book 11, Collection and Delineation of Spatial Data; U.S. Geological Survey: Reston, VA, USA, 2022; ISBN 2328-7055. [Google Scholar]
- Michaletz, P.H.; Dillard, J.G. Fisheries management—A survey of catfish management in the United States and Canada. Fisheries 1999, 24, 6–11. [Google Scholar] [CrossRef]
- Braun, A.P.; Phelps, Q.E. Channel catfish habitat use and diet in the Middle Mississippi River. Am. Midl. Nat. 2016, 175, 47–54. [Google Scholar] [CrossRef]
- Wildhaber, M.L.; Albers, J.L. Laboratory studies of potential competition for food and substrate among early juvenile Missouri River sturgeon and sympatric chub species. N. Am. J. Fish. Manag. 2023. [Google Scholar] [CrossRef]
- Grohs, K.L.; Klumb, R.A.; Chipps, S.R.; Wanner, G.A. Ontogenetic patterns in prey use by pallid sturgeon in the Missouri River, South Dakota and Nebraska. J. Appl. Ichthyol. 2009, 25, 48–53. [Google Scholar] [CrossRef]
- Wildhaber, M.L. Pallid Sturgeon: Scaphirhynchus albus (Forbes and Richardson 1905). In Kansas Fishes; University Press of Kansas: Lawrence, KS, USA, 2014; pp. 99–101. ISBN 978-0-7006-1961-0. [Google Scholar]
- Bellgraph, B.J. Competition Potential between Sauger and Walleye in Non-Native Sympatry: Historical Trends and Resource Overlap in the Middle Missouri River, Montana. Master’s Thesis, Montana State University, Bozeman, MT, USA, 2006. Available online: https://scholarworks.montana.edu/xmlui/handle/1/901 (accessed on 20 December 2023).
- Kelly, B.; Siepker, M.J.; Weber, M.J. Factors associated with detection and distribution of native Brook Trout and introduced Brown Trout in the driftless area of Iowa. Trans. Am. Fish. Soc. 2021, 150, 388–406. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence—Softcover, 2nd ed.; Academic Press: Cambridge, MA, USA, 2018; ISBN 9780128146910. [Google Scholar]
- R Core Team. Available online: https://www.R-project.org/ (accessed on 17 January 2023).
- Fiske, I.J.; Chandler, R.B. Unmarked: An R Package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Neebling, T.E.; Quist, M.C. Comparison of boat electrofishing, trawling, and seining for sampling fish assemblages in Iowa’s Nonwadeable Rivers. N. Am. J. Fish. Manag. 2011, 31, 390–402. [Google Scholar] [CrossRef]
- Steffensen, K.D.; Wilhelm, J.J.; Haas, J.D.; Adams, J.D. Conditional capture probability of pallid sturgeon in benthic trawls. N. Am. J. Fish. Manag. 2015, 35, 626–631. [Google Scholar] [CrossRef]
- Smith, C.D.; Quist, M.C.; Hardy, R.S. Detection probabilities of electrofishing, hoop nets, and benthic trawls for fishes in two Western North American Rivers. J. Fish Wildl. Manag. 2015, 6, 371–391. [Google Scholar] [CrossRef]
- Everett, S.R.; Scarnecchia, D.L.; Ryckman, L.F. Distribution and habitat use of sturgeon chubs (Macrhybopsis gelida) and sicklefin chubs (M. meeki) in the Missouri and Yellowstone Rivers, North Dakota. Hydrobiologia 2004, 527, 183–193. [Google Scholar] [CrossRef]
- Livernois, M.C.; Powers, S.P.; Albins, M.A.; Mareska, J.F. Habitat associations and co-occurrence patterns of two estuarine-dependent predatory fishes. Mar. Coast. Fish. 2020, 12, 64–77. [Google Scholar] [CrossRef]
- Fawcett, T. An introduction to roc analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: An open-source package for r and s plus to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S.; Sturdivant, R.X. Applied Logistic Regression, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; ISBN 9780470582473. [Google Scholar]
- U.S. Geological Survey. USGS Water Data for the Nation: U.S. Geological Survey National Water Information System Database; U.S. Geological Survey: Reston, VA, USA, 2023. [Google Scholar] [CrossRef]
- Wildhaber, M.L.; Tabor, V.M.; Whitaker, J.E.; Allert, A.L.; Mulhern, D.W.; Lamberson, P.J.; Powell, K.L. Ictalurid populations in relation to the presence of a main-stem reservoir in a Midwestern warmwater stream with emphasis on the threatened Neosho madtom. Trans. Am. Fish. Soc. 2000, 129, 1264–1280. [Google Scholar] [CrossRef]
- Herman, P.; Plauck, A.; Utrup, N.; Hill, T. Three Year Summary Age and Growth Report for Sicklefin Chub (Macrhybopsis meeki); U.S. Fish and Wildlife Service: Columbia, MO, USA, 2008; pp. 1–73. Available online: https://meridian.allenpress.com/jfwm/article-supplement/465707/pdf/jfwm-20-086.s5/ (accessed on 16 November 2023).
- Wildhaber, M.L.; DeLonay, A.J.; Papoulias, D.M.; Galat, D.L.; Jacobson, R.B.; Simpkins, D.G.; Braaten, P.J.; Korschgen, C.E.; Mac, M.J. A Conceptual Life-History Model for Pallid and Shovelnose Sturgeon; U.S. Geological Survey: Reston, VA, USA, 2007; p. iv. 18p, ISBN 9781411319059. [Google Scholar]
- Wildhaber, M.L.; DeLonay, A.J.; Papoulias, D.M.; Galat, D.L.; Jacobson, R.B.; Simpkins, D.G.; Braaten, P.J.; Korschgen, C.E.; Mac, M.J. Identifying structural elements needed for development of a predictive life-history model for pallid and shovelnose sturgeons. J. Appl. Ichthyol. 2011, 27, 462–469. [Google Scholar] [CrossRef]
- Wildhaber, M.L.; Dey, R.; Wilke, C.K.; Moran, E.H.; Anderson, C.J.; Franz, K.J. A stochastic bioenergetics model-based approach to translating large river flow and temperature into fish population responses: The pallid sturgeon example. Geol. Soc. Publ. Lond. 2017, 408, 101–118. [Google Scholar] [CrossRef]
- Bellgraph, B.J.; Guy, C.S.; Gardner, W.M.; Leathe, S.A. Competition potential between saugers and walleyes in nonnative sympatry. Trans. Am. Fish. Soc. 2008, 137, 790–800. [Google Scholar] [CrossRef]
- Wildhaber, M.L.; Albers, J.L.; Green, N.S.; Moran, E.H. A fully-stochasticized, age-structured population model for population viability analysis of fish: Lower Missouri River endangered pallid sturgeon example. Ecol. Model. 2017, 359, 434–448. [Google Scholar] [CrossRef]
- Muff, S.; Nilsen, E.B.; O’Hara, R.B.; Nater, C.R. Rewriting results sections in the language of evidence. Trends Ecol. Evol. 2022, 37, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Wildhaber, M.L.; West, B.M.; Green, N.S.; Albers, J.L.; Bennett, K.R.; May, J.H. Occupancy Model Coefficients and Observed Co-Occurrence Simulations for Sicklefin Chub, Sturgeon Chub, and Associated Fishes in the Missouri River; U.S. Geological Survey Data Release: Columbia, MO, USA, 2023. [Google Scholar] [CrossRef]
- Galat, D.L.; Berry, C.R.; Peter, E.J.W.; White, R.G. Missouri River Basin. In Rivers of North America, 2nd ed.; Delong, M.D.B., Benke, A.C., Jardine, T.D., Cushing, C.E., Eds.; Elsevier: Oxford, UK, 2005; pp. 410–461. ISBN 978-0-12-818847-7. [Google Scholar]
- Cross, F.B. Occurrence of the sturgeon chub Hybopsis gelida (Girard) in Kansas. Trans. Kans. Acad. Sci. 1953, 56, 90–91. [Google Scholar] [CrossRef]
- Dieterman, D.J.; Galat, D.L. Large-scale factors associated with sicklefin chub distribution in the Missouri and Lower Yellowstone Rivers. Trans. Am. Fish. Soc. 2004, 133, 577–587. [Google Scholar] [CrossRef]
- Pflieger, W.L.; Grace, T.B. Changes in the fish fauna of the Lower Missouri River, 1940–1983. In Community and Evolutionary Ecology of North Ameircan Stream Fishes; Matthews, W.J., Heins, D.C., Eds.; University of Oklahoma Press: Norman, OK, USA, 1987; pp. 166–177. ISBN 978-0806122359. [Google Scholar]
- Gelwicks, G.; Graham, K.; Galat, D. Status Survey for Sicklefin Chub, Sturgeon Chub, and Flathead Chub in the Missouri River, Missouri: Final Report; Missouri Department of Conservation, Fish and Wildlife Research Center: Columbia, MO, USA, 1996. [Google Scholar]
- Grady, J.M.; Milligan, J. Status of Selected Cyprinid Species and Gear Selectivity at Historic Lower Missouri River Sampling Sites; U.S. Fish and Wildlife Service, Columbia Fisheries Resources Office: Columbia, MO, USA, 1998; p. 52. [Google Scholar]
- Pflieger, W.L.; Smith, P.A. The Fishes of Missouri; Missouri Department of Conservation: Jefferson City, MO, USA, 1997; p. 343. ISBN 1887247114. [Google Scholar]
- Welker, T.L.; Scarnecchia, D.L. Habitat use and population structure of four native minnows (family Cyprinidae) in the Upper Missouri and Lower Yellowstone Rivers, North Dakota (USA). Ecol. Freshw. Fish 2004, 13, 8–22. [Google Scholar] [CrossRef]
- Wildhaber, M.L.; West, B.M.; Ditter, K.K.; Adrian, A.M.; Peterson, A.S. A review of grass carp and related species literature on diet, behavior, toxicology, and physiology focused on informing development of controls for invasive grass carp populations in North America. Fishes 2023, 8, 547. [Google Scholar] [CrossRef]
- Tornabene, B.J.; Smith, T.W.; Tews, A.E.; Beattie, R.P.; Gardner, W.M.; Eby, L.A. Trends in river discharge and water temperature cue spawning movements of blue sucker, Cycleptus elongatus, in an impounded Great Plains river. Copeia 2020, 108, 151–162. [Google Scholar] [CrossRef]
- Dieterman, D.J. Spatial Patterns in Phenotypes and Habitat Use of Sicklefin Chub, Macrhybopsis meeki, in the Missouri and Lower Yellowstone Rivers. Ph.D. Dissertation, University of Missouri, Columbia, MO, USA, 2000. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Type | Detection | Occupancy * | Co-Occurrence |
|---|---|---|---|
| Two-species | Categorical | Categorical | Categorical |
| Project (2) | Sampling year (19) | (none) | |
| HUC 8 subbasin (12) | |||
| Macrohabitat (5) | |||
| Continuous | Continuous | Continuous | |
| Trawl distance | Water temp. ** | Water temp. ** | |
| Water depth ** | Water depth ** | Water depth ** | |
| Water velocity ** | Water velocity ** | Water velocity ** | |
| Turbidity | Turbidity | ||
| Three-species | Continuous | Continuous | Continuous |
| Trawl distance | (none) | (none) | |
| Water depth ** | |||
| Water velocity ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wildhaber, M.L.; West, B.M.; Bennett, K.R.; May, J.H.; Albers, J.L.; Green, N.S. Sicklefin Chub (Macrhybopsis meeki) and Sturgeon Chub (M. gelida) Temporal and Spatial Patterns from Extant Population Monitoring and Habitat Data Spanning 23 Years. Fishes 2024, 9, 43. https://doi.org/10.3390/fishes9020043
Wildhaber ML, West BM, Bennett KR, May JH, Albers JL, Green NS. Sicklefin Chub (Macrhybopsis meeki) and Sturgeon Chub (M. gelida) Temporal and Spatial Patterns from Extant Population Monitoring and Habitat Data Spanning 23 Years. Fishes. 2024; 9(2):43. https://doi.org/10.3390/fishes9020043
Chicago/Turabian StyleWildhaber, Mark L., Benjamin M. West, Kendell R. Bennett, Jack H. May, Janice L. Albers, and Nicholas S. Green. 2024. "Sicklefin Chub (Macrhybopsis meeki) and Sturgeon Chub (M. gelida) Temporal and Spatial Patterns from Extant Population Monitoring and Habitat Data Spanning 23 Years" Fishes 9, no. 2: 43. https://doi.org/10.3390/fishes9020043
APA StyleWildhaber, M. L., West, B. M., Bennett, K. R., May, J. H., Albers, J. L., & Green, N. S. (2024). Sicklefin Chub (Macrhybopsis meeki) and Sturgeon Chub (M. gelida) Temporal and Spatial Patterns from Extant Population Monitoring and Habitat Data Spanning 23 Years. Fishes, 9(2), 43. https://doi.org/10.3390/fishes9020043