
Evolution of Nipah Virus Infection: Past, Present, and Future Considerations

 ,
,
Abstract
:1. Introduction
2. Epidemiology

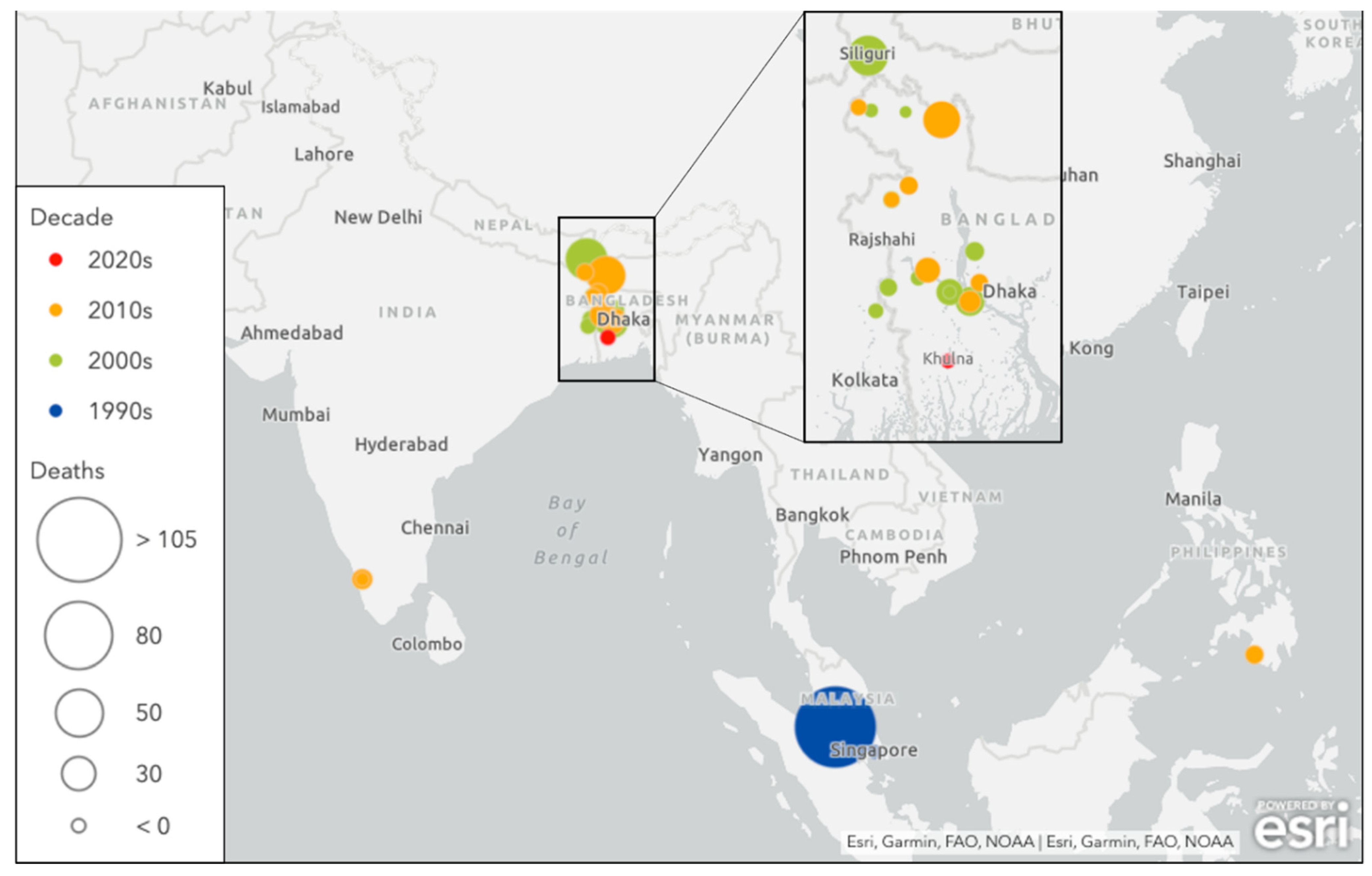{kind=link}
{kind=link}
| Country | Year(s) | Cases N | Fatalities (% CFR) | Exposure History, Transmission, Clinical Features | Reference |
|---|---|---|---|---|---|
| Malaysia | Sep 1998–Dec 1999 | 265 | 105 (38.5%) |
| Chua 2000 [1] Goh et al., 2000 [18] |
| Singapore | March 1999 | 11 | 1 (9.1%) |
| Paton et al., 1999 [6] |
| Bangladesh | Jan 2001–Feb 2015 (17 outbreaks) | 261 | 19 (75.9%) |
| Luby et al., 2006 [13] Gurley et al., 2017 [10] Nikolay et al., 2019 [3] |
| India | Jan–Feb 2001 Apr 2007 May 2018 June 2019 | 92 | 68 (73.9%) |
| Chadha et al., 2006 [7] Banerjee et al., 2019 [19] Arunkumar et al., 2019 [20] |
| Philippines | Mar–Apr 2014 | 17 | 9 (52.9%) |
| Ching et al., 2015 [8] |
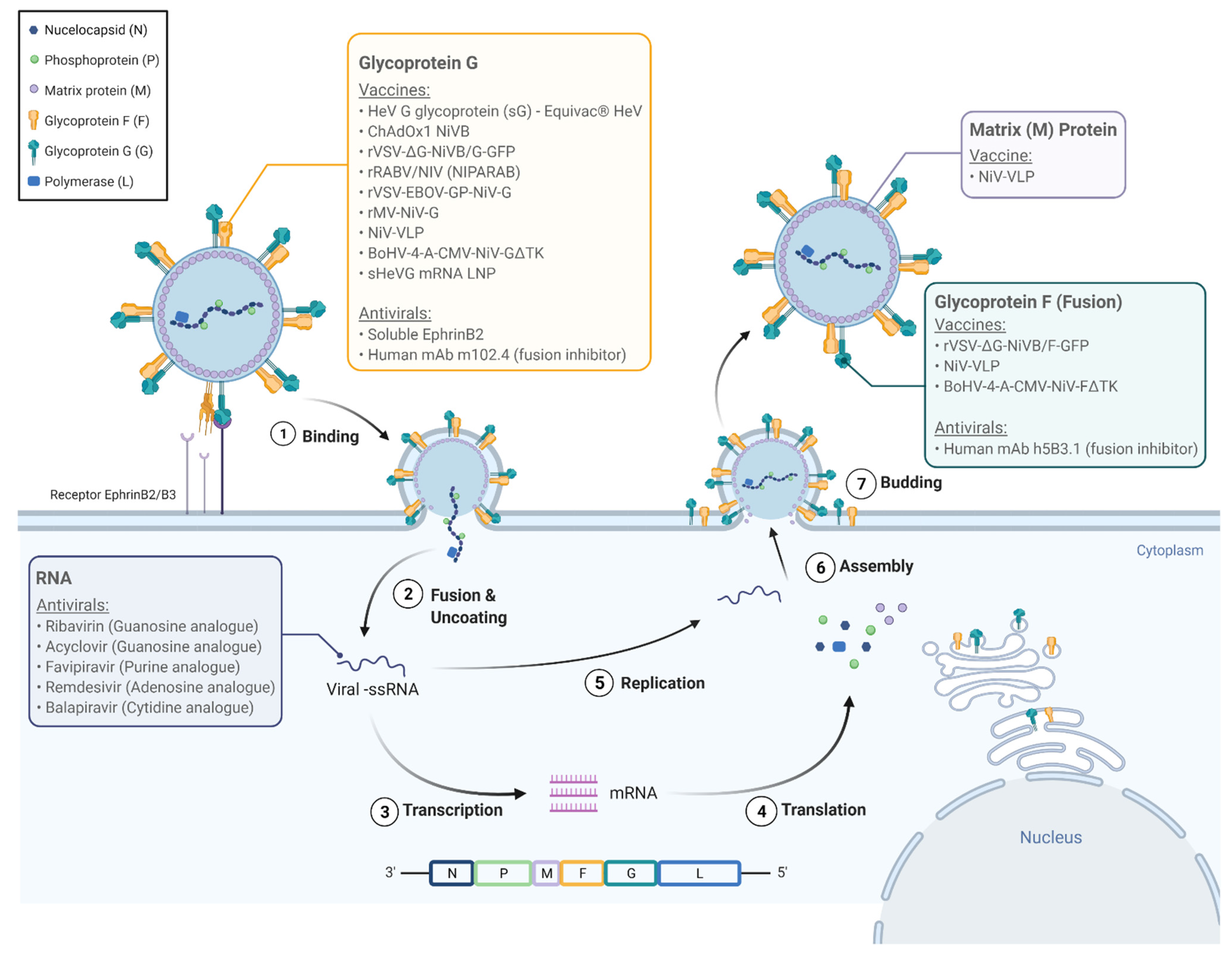
3. Molecular Biology of the Virus
4. Pathogenesis and Clinical Features
5. Antiviral Therapies in Development
6. Vaccines in Development
| Vaccines | Description | Animal Model | Reference |
|---|---|---|---|
| Subunit-based | |||
| HeV-sG (Equivac® HeV) | Subunit vaccine based on soluble HeV G glycoprotein. Elicits cross protective immune response against HeV and NiV. Available for horses in Australia | Ferret | Pallister et al., 2013 [46] |
| Vector-based | |||
| ChAdOx1 NiVB | Recombinant simian adenovirus-based vaccine encoding NiV-B glycoprotein G | Golden Syrian hamster | Van Doremalen et al., 2019 [47] |
| rVSV-ΔG-NiVB/F-GFP rVSV-ΔG-NiVB/G-GFP | Recombinant vesicular stomatitis virus (VSV) vaccine expressing NiV-B F or G | African green monkey challenge | Mire et al., 2019 [48] |
| rRABV/NIV (NIPARAB) | Recombinant rabies virus vector expressing NiV G | C57BL/6 mice | Keshwara et al., 2019 [49] |
| rVSV-EBOV-GP-NiV-G | Recombinant VSV vector expressing Ebola virus glycoprotein and NiV G | African green monkey challenge | Prescott et al., 2015 [50] |
| rMV-NiV-G | Recombinant measles virus vaccine (rMV) expressing NiV G | African green monkey challenge | Yoneda et al., 2013 [51] |
| BoHV-4-A-CMV-NiV-GΔTK BoHV-4-A-CMV-NiV-FΔTK | Recombinant bovine herpesvirus vaccine (BoHV) expressing NiV G or NiV F | Pig | Pedrera et al., 2020 [52] |
| Virus-like particle-based | |||
| NiV-VLP vaccine | Purified Nipah virus-like particles G, F, and M proteins | Golden Syrian hamster challenge | Walpita et al., 2017 [53] |
| mRNA-based | |||
| sHeVG mRNA LNP | mRNA vaccine encoding soluble HeV glycoprotein (sHeVG) subunit | Syrian hamster | Lo et al., 2020 [54] |
7. Non-Pharmacologic Interventions
8. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chua, B. Nipah Virus: A Recently Emergent Deadly Paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Enserink, M. New Virus Fingered in Malaysian Epidemic. Science 1999, 284, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B.; Salje, H.; Hossain, M.J.; Khan, A.K.M.D.; Sazzad, H.M.S.; Rahman, M.; Daszak, P.; Ströher, U.; Pulliam, J.R.C.; Kilpatrick, A.M.; et al. Transmission of Nipah Virus—14 Years of Investigations in Bangladesh. N. Engl. J. Med. 2019, 380, 1804–1814. [Google Scholar] [CrossRef] [PubMed]
- Spiropoulou, C.F. Nipah Virus Outbreaks: Still Small but Extremely Lethal. J. Infect. Dis. 2019, 219, 1855–1857. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Update: Outbreak of Nipah Virus—Malaysia and Singapore, 1999. MMWR Morb. Mortal. Wkly. Rep. 1999, 48, 335–337. [Google Scholar]
- Paton, N.I.; Leo, Y.S.; Zaki, S.R.; Auchus, A.P.; Lee, K.E.; Ling, A.E.; Chew, S.K.; Ang, B.; Rollin, P.E.; Umapathi, T.; et al. Outbreak of Nipah-Virus Infection among Abattoir Workers in Singapore. Lancet 1999, 354, 1253–1256. [Google Scholar] [CrossRef]
- Chadha, M.S.; Comer, J.A.; Lowe, L.; Rota, P.A.; Rollin, P.E.; Bellini, W.J.; Ksiazek, T.G.; Mishra, A.C. Nipah Virus-Associated Encephalitis Outbreak, Siliguri, India. Emerg. Infect. Dis. 2006, 12, 235–240. [Google Scholar] [CrossRef]
- Ching, P.K.G.; de Los Reyes, V.C.; Sucaldito, M.N.; Tayag, E.; Columna-Vingno, A.B.; Malbas, F.F.; Bolo, G.C.; Sejvar, J.J.; Eagles, D.; Playford, G.; et al. Outbreak of Henipavirus Infection, Philippines, 2014. Emerg. Infect. Dis. 2015, 21, 328–331. [Google Scholar] [CrossRef]
- Gurley, E.S.; Montgomery, J.M.; Hossain, M.J.; Bell, M.; Azad, A.K.; Ra, M.; Abdur, M.; Molla, R.; Carroll, D.S.; Ksiazek, T.G.; et al. Person-to-Person Transmission of Nipah Virus in a Bangladeshi Community. Emerg. Infect. Dis. 2007, 13, 1031–1037. [Google Scholar] [CrossRef]
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Sharker, M.A.Y.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of Humans, Bats, Trees, and Culture in Nipah Virus. Emerg. Infect. Dis. 2017, 23, 1446. [Google Scholar] [CrossRef] [Green Version]
- Hsu, V.P.; Hossain, M.J.; Parashar, U.D.; Ali, M.M.; Ksiazek, T.G.; Kuzmin, I.; Niezgoda, M.; Rupprecht, C.; Bresee, J.; Breiman, R.F. Nipah Virus Encephalitis Reemergence, Bangladesh. Emerg. Infect. Dis. 2004, 10, 2082–2087. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.U.; Gurley, E.S.; Hossain, M.J.; Nahar, N.; Sharker, M.A.Y.; Luby, S.P. A Randomized Controlled Trial of Interventions to Impede Date Palm Sap Contamination by Bats to Prevent Nipah Virus Transmission in Bangladesh. PLoS ONE 2012, 7, 0042689. [Google Scholar] [CrossRef]
- Luby, S.P.; Rahman, M.; Hossain, M.J.; Blum, L.S.; Husain, M.M.; Gurley, E.; Khan, R.; Ahmed, B.N.; Rahman, S.; Nahar, N.; et al. Foodborne Transmission of Nipah Virus, Bangladesh. Emerg. Infect. Dis. 2006, 12, 1888–1894. [Google Scholar] [CrossRef] [PubMed]
- Luby, S.P.; Hossain, M.J.; Gurley, E.S.; Ahmed, B.; Banu, S.; Khan, S.U.; Homaira, N.; Rota, P.A.; Rollin, P.E.; Comer, J.A.; et al. Recurrent Zoonotic Transmission of Nipah Virus into Humans. Emerg. Infect. Dis. 2009, 15, 2001–2007. [Google Scholar] [CrossRef]
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Halpin, K.; Hyatt, A.D.; Fogarty, R.; Middleton, D.; Bingham, J.; Epstein, J.H.; Rahman, S.A.; Hughes, T.; Smith, C.; Field, H.E.; et al. Pteropid Bats Are Confirmed as the Reservoir Hosts of Henipaviruses: A Comprehensive Experimental Study of Virus Transmission. Am. J. Trop. Med. Hyg. 2011, 85, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Chow, V.T.K.; Tambyah, P.A.; Yeo, W.M.; Phoon, M.C.; Howe, J. Diagnosis of Nipah Virus Encephalitis by Electron Microscopy of Cerebrospinal Fluid. J. Clin. Virol. 2000, 19, 143–147. [Google Scholar] [CrossRef]
- Goh, K.J.; Tan, C.T.; Chew, N.K.; Tan, P.S.K.; Kamarulzaman, A.; Sarji, S.A.; Wong, K.T.; Abdullah, B.J.J.; Chua, K.B.; Lam, S.K. Clinical Features of Nipah Virus Encephalitis among Pig Farmers in Malaysia. N. Engl. J. Med. 2000, 342, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Niyas, V.K.M.; Soneja, M.; Shibeesh, A.P.; Basheer, M.; Sadanandan, R.; Wig, N.; Biswas, A. First Experience of Ribavirin Postexposure Prophylaxis for Nipah Virus, Tried during the 2018 Outbreak in Kerala, India. J. Infect. 2019, 78, 497–499. [Google Scholar] [CrossRef]
- Arunkumar, G.; Chandni, R.; Mourya, D.T.; Singh, S.K.; Sadanandan, R.; Sudan, P.; Bhargava, B. Outbreak Investigation of Nipah Virus Disease in Kerala, India, 2018. J. Infect. Dis. 2019, 219, 1867–1878. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.E.; Islam, A.; Crameri, G.; Todd, S.; Islam, A.; Khan, S.U.; Foord, A.; Rahman, M.Z.; Mendenhall, I.H.; Luby, S.P.; et al. Isolation and Full-Genome Characterization of Nipah Viruses from Bats, Bangladesh. Emerg. Infect. Dis. 2019, 25, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Yob, J.M.; Field, H.; Rashdi, A.M.; Morrissy, C.; van der Heide, B.; Rota, P.; bin Adzhar, A.; White, J.; Daniels, P.; Jamaluddin, A.; et al. Nipah Virus Infection in Bats (Order Chiroptera) in Peninsular Malaysia. Emerg. Infect. Dis. 2001, 7, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.D.; Raut, C.G.; Shete, A.M.; Mishra, A.C.; Towner, J.S.; Nichol, S.T.; Mourya, D.T. Short Report: Detection of Nipah Virus RNA in Fruit Bat (Pteropus Giganteus) from India. Am. J. Trop. Med. Hyg. 2012, 87, 576–578. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.S.; Sazzad, H.M.S.; Satter, S.M.; Sultana, S.; Hossain, M.J.; Hasan, M.; Rahman, M.; Campbell, S.; Cannon, D.L.; Ströher, U.; et al. Nipah Virus Transmission from Bats to Humans Associated with Drinking Traditional Liquor Made from Date Palm Sap, Bangladesh, 2011-2014. Emerg. Infect. Dis. 2016, 22, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, J.M.; Hossain, M.J.; Gurley, E.; Carroll, D.S.; Croisier, A.; Bertherat, E.; Asgari, N.; Formenty, P.; Keeler, N.; Comer, J.; et al. Risk Factors for Nipah Virus Encephalitis in Bangladesh. Emerg. Infect. Dis. 2008, 14, 1526–1532. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, V.; Aslan, H.; Ainouze, M.; Guerbois, M.; Fabian Wild, T.; Buckland, R.; Langedijk, J.P.M. Evidence of a Potential Receptor-Binding Site on the Nipah Virus G Protein (NiV-G): Identification of Globular Head Residues with a Role in Fusion Promotion and Their Localization on an NiV-G Structural Model. J. Virol. 2006, 80, 7546–7554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaparte, M.I.; Dimitrov, A.S.; Bossart, K.N.; Crameri, G.; Mungall, B.A.; Bishop, K.A.; Choudhry, V.; Dimitrov, D.S.; Wang, L.F.; Eaton, B.T.; et al. Ephrin-B2 Ligand Is a Functional Receptor for Hendra Virus and Nipah Virus. Proc. Natl. Acad. Sci. USA 2005, 102, 10652–10657. [Google Scholar] [CrossRef] [Green Version]
- Negrete, O.A.; Levroney, E.L.; Aguilar, H.C.; Bertolotti-Ciarlet, A.; Nazarian, R.; Tajyar, S.; Lee, B. EphrinB2 Is the Entry Receptor for Nipah Virus, an Emergent Deadly Paramyxovirus. Nature 2005, 436, 401–405. [Google Scholar] [CrossRef]
- Aguilar, H.C.; Matreyek, K.; Filone, C.; Hashimi, S.; Levroney, E.; Negrete, O.; ABertolotti-Ciarlet, A.; Choi, D.; McHardy, I.; Fulcher, J.; et al. N-Glycans on Nipah Virus Fusion Protein Protect against Neutralization but Reduce Membrane Fusion and Viral Entry. J. Virol. 2006, 80, 4878–4889. [Google Scholar] [CrossRef] [Green Version]
- Patch, J.R.; Crameri, G.; Wang, L.F.; Eaton, B.T.; Broder, C.C. Quantitative Analysis of Nipah Virus Proteins Released as Virus-like Particles Reveals Central Role for the Matrix Protein. Virol. J. 2007, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Uchida, S.; Horie, R.; Sato, H.; Kai, C.; Yoneda, M. Possible Role of the Nipah Virus V Protein in the Regulation of the Interferon Beta Induction by Interacting with UBX Domain-Containing Protein1. Sci. Rep. 2018, 8, 7682. [Google Scholar] [CrossRef] [Green Version]
- Harcourt, B.H.; Lowe, L.; Tamin, A.; Liu, X.; Bankamp, B.; Bowden, N.; Rollin, P.E.; Comer, J.A.; Ksiazek, T.G.; Hossain, M.J.; et al. Genetic Characterization of Nipah Virus, Bangladesh, 2004. Emerg. Infect. Dis. 2005, 11, 1594–1597. [Google Scholar] [CrossRef]
- Wong, K.T.; Shieh, W.J.; Kumar, S.; Norain, K.; Abdullah, W.; Guarner, J.; Goldsmith, C.S.; Chua, K.B.; Lam, S.K.; Tan, C.T.; et al. Nipah Virus Infection: Pathology and Pathogenesis of an Emerging Paramyxoviral Zoonosis. Am. J. Pathol. 2002, 161, 2153–2167. [Google Scholar] [CrossRef]
- Mire, C.E.; Satterfield, B.A.; Geisbert, J.B.; Agans, K.N.; Borise, V.; Yan, L.; Chan, Y.; Cross, R.W.; Fenton, K.A.; Broder, C.C.; et al. Pathogenic Differences between Nipah Virus Bangladesh and Malaysia Strains in Primates: Implications for Antibody Therapy. Sci. Rep. 2016, 6, 30616. [Google Scholar] [CrossRef]
- Chong, H.T.; Kamarulzaman, A.; Tan, C.T.; Goh, K.J.; Thayaparan, T.; Kunjapan, S.R.; Chew, N.K.; Chua, K.B.; Lam, S.K. Treatment of Acute Nipah Encephalitis with Ribavirin. Ann. Neurol. 2001, 49, 810–813. [Google Scholar] [CrossRef]
- Freiberg, A.N.; Worthy, M.N.; Lee, B.; Holbrook, M.R. Combined Chloroquine and Ribavirin Treatment Does Not Prevent Death in a Hamster Model of Nipah and Hendra Virus Infection. J. Gen. Virol. 2010, 91, 765–772. [Google Scholar] [CrossRef]
- Georges-Courbot, M.C.; Contamin, H.; Faure, C.; Loth, P.; Baize, S.; Leyssen, P.; Neyts, J.; Deubel, V. Poly(I)-Poly(C12U) but Not Ribavirin Prevents Death in a Hamster Model of Nipah Virus Infection. Antimicrob. Agents Chemother. 2006, 50, 1768–1772. [Google Scholar] [CrossRef] [Green Version]
- Dawes, B.E.; Kalveram, B.; Ikegami, T.; Juelich, T.; Smith, J.K.; Zhang, L.; Park, A.; Lee, B.; Komeno, T.; Furuta, Y.; et al. Favipiravir (T-705) Protects against Nipah Virus Infection in the Hamster Model. Sci. Rep. 2018, 8, 7604. [Google Scholar] [CrossRef] [Green Version]
- Lo, M.K.; Jordan, R.; Arvey, A.; Sudhamsu, J.; Shrivastava-Ranjan, P.; Hotard, A.L.; Flint, M.; McMullan, L.K.; Siegel, D.; Clarke, M.O.; et al. GS-5734 and Its Parent Nucleoside Analog Inhibit Filo-, Pneumo-, and Paramyxoviruses. Sci. Rep. 2017, 7, 43395. [Google Scholar] [CrossRef]
- Lo, M.K.; Feldmann, F.; Gary, J.M.; Jordan, R.; Bannister, R.; Cronin, J.; Patel, N.R.; Klena, J.D.; Nichol, S.T.; Cihlar, T.; et al. Remdesivir (GS-5734) Protects African Green Monkeys from Nipah Virus Challenge. Sci. Transl. Med. 2019, 9242, 1–7. [Google Scholar] [CrossRef]
- Hotard, A.L.; He, B.; Nichol, S.T.; Spiropoulou, C.F.; Lo, M.K. 4′-Azidocytidine (R1479) Inhibits Henipaviruses and Other Paramyxoviruses with High Potency. Antivir. Res. 2017, 144, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Bossart, K.N.; Zhu, Z.; Middleton, D.; Klippel, J.; Crameri, G.; Bingham, J.; McEachern, J.A.; Green, D.; Hancock, T.J.; Chan, Y.P.; et al. A Neutralizing Human Monoclonal Antibody Protects against Lethal Disease in a New Ferret Model of Acute Nipah Virus Infection. PLoS Pathog. 2009, 5, e1000642. [Google Scholar] [CrossRef] [PubMed]
- Geisbert, T.W.; Mire, C.E.; Geisbert, J.B.; Chan, Y.P.; Agans, K.N.; Feldmann, F.; Fenton, K.A.; Zhu, Z.; Dimitrov, D.S.; Scott, D.P.; et al. Therapeutic Treatment of Nipah Virus Infection in Nonhuman Primates with a Neutralizing Human Monoclonal Antibody. Sci. Transl. Med. 2014, 6, 242ra82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mire, C.E.; Chan, Y.P.; Borisevich, V.; Cross, R.W.; Yan, L.; Agans, K.N.; Dang, H.V.; Veesler, D.; Fenton, K.A.; Geisbert, T.W.; et al. A Cross-Reactive Humanized Monoclonal Antibody Targeting Fusion Glycoprotein Function Protects Ferrets Against Lethal Nipah Virus and Hendra Virus Infection. J. Infect. Dis. 2020, 221, S471–S479. [Google Scholar] [CrossRef] [PubMed]
- Tamin, A.; Harcourt, B.H.; Ksiazek, T.G.; Rollin, P.E.; Bellini, W.J.; Rota, P.A. Functional Properties of the Fusion and Attachment Glycoproteins of Nipah Virus. Virology 2002, 296, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Pallister, J.A.; Klein, R.; Arkinstall, R.; Haining, J.; Long, F.; White, J.R.; Payne, J.; Feng, Y.R.; Wang, L.F.; Broder, C.C.; et al. Vaccination of Ferrets with a Recombinant G Glycoprotein Subunit Vaccine Provides Protection against Nipah Virus Disease for over 12 Months. Virol. J. 2013, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Van Doremalen, N.; Lambe, T.; Sebastian, S.; Bushmaker, T.; Fischer, R.; Feldmann, F.; Haddock, E.; Letko, M.; Avanzato, V.A.; Rissanen, I.; et al. A Single-Dose ChAdOx1-Vectored Vaccine Provides Complete Protection against Nipah Bangladesh and Malaysia in Syrian Golden Hamsters. PLoS Negl. Trop. Dis. 2019, 13, e0007462. [Google Scholar] [CrossRef]
- Mire, C.E.; Geisbert, J.B.; Agans, K.N.; Versteeg, K.M.; Deer, D.J.; Satterfield, B.A.; Fenton, K.A.; Geisbert, T.W. Use of Single-Injection Recombinant Vesicular Stomatitis Virus Vaccine to Protect Nonhuman Primates against Lethal Nipah Virus Disease. Emerg. Infect. Dis. 2019, 25, 1144–1152. [Google Scholar] [CrossRef] [Green Version]
- Keshwara, R.; Shiels, T.; Postnikova, E.; Kurup, D.; Wirblich, C.; Johnson, R.F.; Schnell, M.J. Rabies-Based Vaccine Induces Potent Immune Responses against Nipah Virus. NPJ Vaccines 2019, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Prescott, J.; DeBuysscher, B.L.; Feldmann, F.; Gardner, D.J.; Haddock, E.; Martellaro, C.; Scott, D.; Feldmann, H. Single-Dose Live-Attenuated Vesicular Stomatitis Virus-Based Vaccine Protects African Green Monkeys from Nipah Virus Disease. Vaccine 2015, 33, 2823–2829. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, M.; Georges-Courbot, M.C.; Ikeda, F.; Ishii, M.; Nagata, N.; Jacquot, F.; Raoul, H.; Sato, H.; Kai, C. Recombinant Measles Virus Vaccine Expressing the Nipah Virus Glycoprotein Protects against Lethal Nipah Virus Challenge. PLoS ONE 2013, 8, e0058414. [Google Scholar] [CrossRef]
- Pedrera, M.; Macchi, F.; McLean, R.K.; Franceschi, V.; Thakur, N.; Russo, L.; Medfai, L.; Todd, S.; Tchilian, E.Z.; Audonnet, J.-C.; et al. Bovine Herpesvirus-4-Vectored Delivery of Nipah Virus Glycoproteins Enhances T Cell Immunogenicity in Pigs. Vaccines 2020, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Walpita, P.; Cong, Y.; Jahrling, P.B.; Rojas, O.; Postnikova, E.; Yu, S.; Johns, L.; Holbrook, M.R. A VLP-Based Vaccine Provides Complete Protection against Nipah Virus Challenge Following Multiple-Dose or Single-Dose Vaccination Schedules in a Hamster Model. NPJ Vaccines 2017, 2, 21. [Google Scholar] [CrossRef]
- Lo, M.K.; Spengler, J.R.; Welch, S.R.; Harmon, J.R.; Coleman-McCray, J.D.; Scholte, F.E.M.; Shrivastava-Ranjan, P.; Montgomery, J.M.; Nichol, S.T.; Weissman, D.; et al. Evaluation of a Single-Dose Nucleoside-Modified Messenger RNA Vaccine Encoding Hendra Virus-Soluble Glycoprotein Against Lethal Nipah Virus Challenge in Syrian Hamsters. J. Infect. Dis. 2020, 221, S493–S498. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). Outbreak of Hendra-like Virus—Malaysia and Singapore, 1998–1999. MMWR Morb. Mortal. Wkly. Rep. 1999, 48, 265–269. [Google Scholar]
- Luby, S.P.; Gurley, E.S.; Hossain, M.J. Transmission of Human Infection with Nipah Virus. Clin. Infect. Dis. 2009, 49, 1743–1748. [Google Scholar] [CrossRef] [Green Version]


| Drug | Description | Experimental Model | Reference |
|---|---|---|---|
| Chloroquine | 4-aminoquinoline | In vitro | Freiberg et al., 2010 [36] |
| Ribavirin | Guanosine analogue | In vitro | Freiberg et al., 2010 [36] |
| Acyclovir | Guanosine analogue | Historical review | Paton et al., 1999 [6] |
| Favipiravir | Purine analogue | In vitro Syrian hamster | Dawes et al., 2018 [38] |
| Remdesivir (GS-5734) | Adenosine analogue | In vitro African green monkey | Lo et al., 2017 [39] Lo et al., 2019 [40] |
| Balapiravir (R1479) | Cytidine analogue | In vitro | Hotard et al., 2017 [41] |
| Poly(I)-poly(C12U) | Interferon inducer | In vitro Hamster | Georges-Courbot et al., 2006 [37] |
| EphrinB2 | G glycoprotein fusion inhibitor | In vitro | Negrete et al., 2005 [28] |
| Human mAb m102.4 | G glycoprotein fusion inhibitor | Ferrets African green monkey | Bossart et al., 2009 [42] Geisbert et al., 2014 [43] |
| Human mAb h5B3.1 | F glycoprotein fusion inhibitor | Ferrets | Mire et al., 2020 [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hauser, N.; Gushiken, A.C.; Narayanan, S.; Kottilil, S.; Chua, J.V. Evolution of Nipah Virus Infection: Past, Present, and Future Considerations. Trop. Med. Infect. Dis. 2021, 6, 24. https://doi.org/10.3390/tropicalmed6010024
Hauser N, Gushiken AC, Narayanan S, Kottilil S, Chua JV. Evolution of Nipah Virus Infection: Past, Present, and Future Considerations. Tropical Medicine and Infectious Disease. 2021; 6(1):24. https://doi.org/10.3390/tropicalmed6010024
Chicago/Turabian StyleHauser, Naomi, Alexis C. Gushiken, Shivakumar Narayanan, Shyam Kottilil, and Joel V. Chua. 2021. "Evolution of Nipah Virus Infection: Past, Present, and Future Considerations" Tropical Medicine and Infectious Disease 6, no. 1: 24. https://doi.org/10.3390/tropicalmed6010024
APA StyleHauser, N., Gushiken, A. C., Narayanan, S., Kottilil, S., & Chua, J. V. (2021). Evolution of Nipah Virus Infection: Past, Present, and Future Considerations. Tropical Medicine and Infectious Disease, 6(1), 24. https://doi.org/10.3390/tropicalmed6010024