
The Efficacy of Vaccination in the Prevention of Renal and Genital Leptospirosis in Experimentally Infected Sheep

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Animals
2.3. Experimental Groups
2.4. Vaccines
2.5. Bacterial Strain
2.6. Sampling
2.7. Laboratory Procedures
2.7.1. Serology (MAT)
2.7.2. Polymerase Chain Reaction (PCR)
2.8. Data Analysis
3. Results
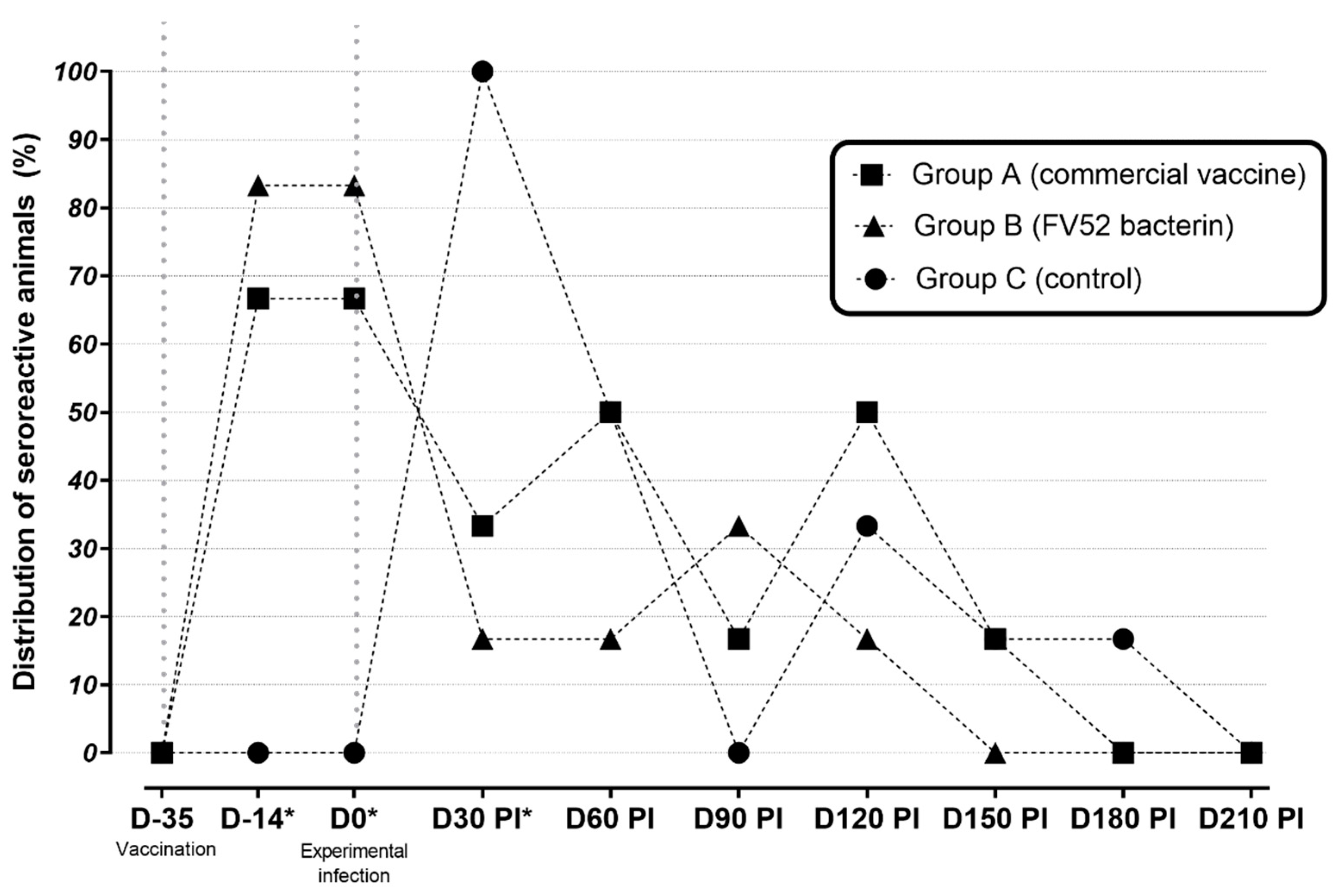
3.1. Serological Results
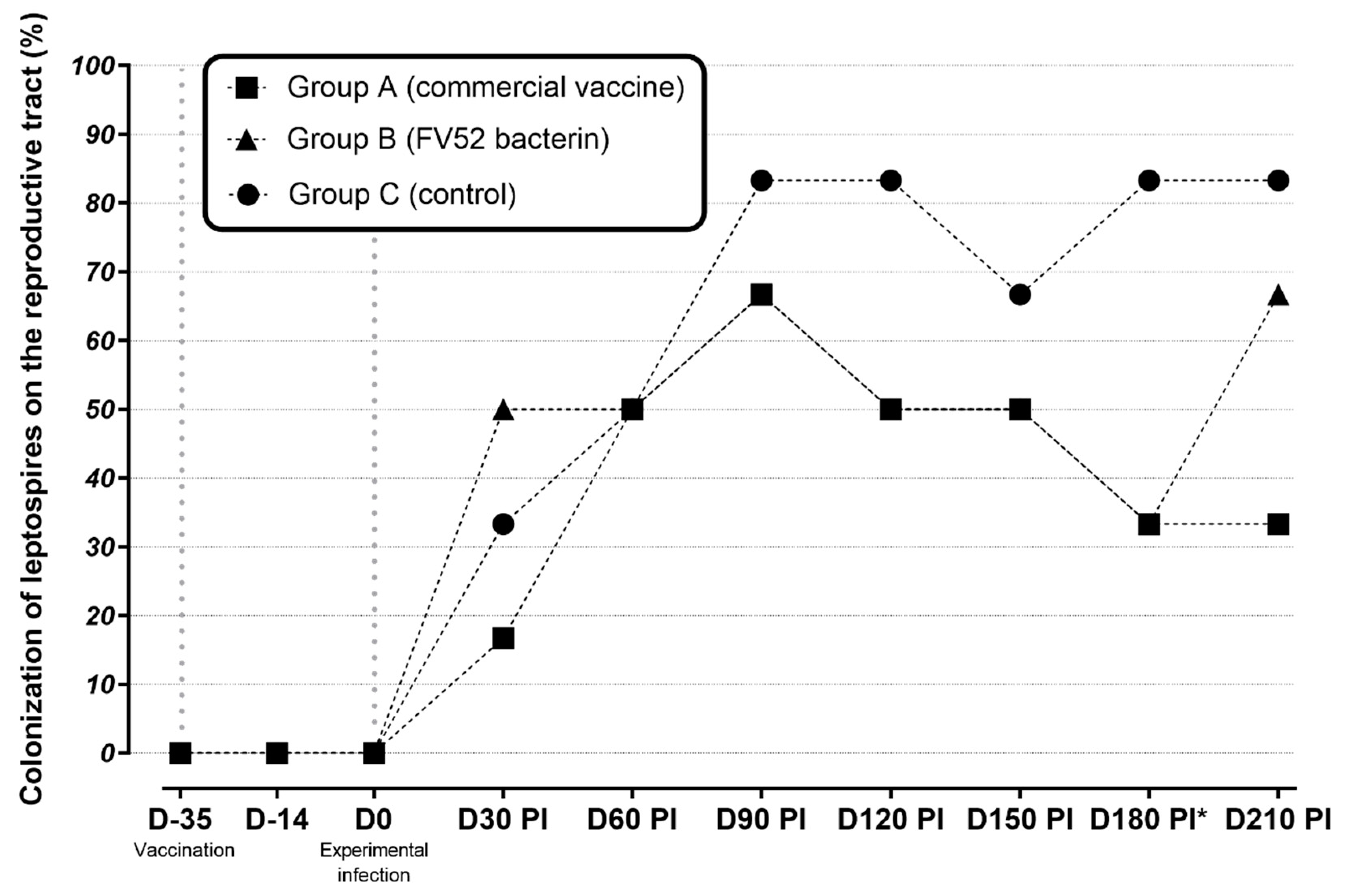
3.2. Urinary PCR Results
3.3. Genital (CVM and/or Uterine Fragment) PCR Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lata, K.S.; Vaghasia, V.; Bhairappanvar, S.; Patel, S.; Das, J. Vaccine Design Against Leptospirosis Using an Immunoinformatic Approach. In Immunoinformatics. Methods in Molecular Biology; Humana: New York, NY, USA, 2020; Volume 2131, pp. 173–184. [Google Scholar] [CrossRef]
- Ellis, W.A. Animal Leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 99–137. [Google Scholar] [CrossRef]
- Ellis, W.A.; Thiermann, A.B. Isolation of Leptospira interrogans Serovar Bratislava from Sows in Iowa. Am. J. Vet. Res. 1986, 47, 1458–1460. [Google Scholar]
- Dhaliwal, G.S.; Murray, R.D.; Ellis, W.A. Reproductive Performance of Dairy Herds Infected with Leptospira interrogans Serovar Hardjo Relative to the Year of Diagnosis. Vet. Rec. 1996, 138, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, A.P.; Pestana, C.; Medeiros, M.A.; Lilenbaum, W. High Frequency of Leptospiral Vaginal Carriers among Slaughtered Cows. Anim. Reprod. Sci. 2017, 178, 50–54. [Google Scholar] [CrossRef]
- Bielanski, A.; Surujballi, O.; Golsteyn Thomas, E.; Tanaka, E. Sanitary Status of Oocytes and Embryos Collected from Heifers Experimentally Exposed to Leptospira borgpetersenii Serovar Hardjobovis. Anim. Reprod. Sci. 1998, 54, 65–73. [Google Scholar] [CrossRef]
- Cabral Pires, B.; Berzin Grapiglia, J.; Moreira, L.; Jaeger, L.H.; Carvalho-Costa, F.A.; Lilenbaum, W. Occurrence of Uterine Carriers for Leptospira interrogans on Slaughtered Cows. Microb. Pathog. 2018, 114, 163–165. [Google Scholar] [CrossRef] [PubMed]
- Di Azevedo, M.I.N.; Pires, B.C.; Libonati, H.; Pinto, P.S.; Cardoso Barbosa, L.F.; Carvalho-Costa, F.A.; Lilenbaum, W. Extra-Renal Bovine Leptospirosis: Molecular Characterization of the Leptospira interrogans Sejroe Serogroup on the Uterus of Non-Pregnant Cows. Vet. Microbiol. 2020, 250, 108869. [Google Scholar] [CrossRef] [PubMed]
- Di Azevedo, M.I.N.; Pires, B.C.; Barbosa, L.F.C.; Carvalho-Costa, F.A.; Lilenbaum, W. Characterization of Leptospiral DNA in the Follicular Fluid of Non-Pregnant Cows. Vet. Rec. 2021, 188, e143. [Google Scholar] [CrossRef]
- Dos Santos Pereira, P.V.; Di Azevedo, M.I.N.; Dos Santos Baptista Borges, A.L.; Loureiro, A.P.; Martins, G.; Carvalho-Costa, F.A.; Souza-Fabjan, J.M.G.; Lilenbaum, W. Bovine Genital Leptospirosis: Evidence of Ovarian Infection by Leptospira interrogans. Vet. Microbiol. 2022, 271, 109489. [Google Scholar] [CrossRef]
- Loureiro, A.P.; Hamond, C.; Pinto, P.; Bremont, S.; Bourhy, P.; Lilenbaum, W. Molecular Analysis of Leptospires from Serogroup Sejroe Obtained from Asymptomatic Cattle in Rio de Janeiro—Brazil Reveals Genetic Proximity to Serovar Guaricura. Res. Vet. Sci. 2016, 105, 249–253. [Google Scholar] [CrossRef]
- Aymée, L.; Nogueira Di Azevedo, M.I.; de Souza Pedrosa, J.; de Melo, J.D.S.L.; Carvalho-Costa, F.A.; Lilenbaum, W. The Role of Leptospira santarosai Serovar Guaricura as Agent of Bovine Genital Leptospirosis. Vet. Microbiol. 2022, 268, 109413. [Google Scholar] [CrossRef]
- Pinto, P.S.; Barbosa, C.; Ferreira, A.M.R.; Lilenbaum, W. Short Communication: Uterine Leptospiral Infection Is Strongly Associated to Strains of Serogroup Sejroe on Experimentally Infected Hamsters. Microb. Pathog. 2020, 142, 104030. [Google Scholar] [CrossRef] [PubMed]
- Rocha, B.R.; Balaro, M.; Pereira, P.V.; Martins, G.; Lilenbaum, W. Chronic Experimental Genital Leptospirosis with Autochthonous Leptospira santarosai Strains of Serogroup Sejroe. Small Rumin. Res. 2018, 164, 28–31. [Google Scholar] [CrossRef]
- da Rosa, M.C.; Martins, G.; Rocha, B.R.; Correia, L.; Ferronato, G.; Lilenbaum, W.; Dellagostin, O.A. Assessment of the Immunogenicity of the Leptospiral LipL32, LigAni, and LigBrep Recombinant Proteins in the Sheep Model. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Rocha, B.R.; Martins, G.; Lilenbaum, W. An Historical View of the Experimental Leptospiral Infection in Ruminants. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101532. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.F.; Farias, P.J.A.; Silva, M.L.C.R.; Araújo Júnior, J.P.; Malossi, C.D.; Ullmann, L.S.; Costa, D.F.; Higino, S.S.S.; Azevedo, S.S.; Alves, C.J. High Frequency of Genital Carriers of Leptospira sp. in Sheep Slaughtered in the Semi-Arid Region of Northeastern Brazil. Trop. Anim. Health Prod. 2019, 51, 43–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, W.A.; Bryson, D.G.; Neill, S.D.; McParland, P.J.; Malone, F.E. Possible Involvement of Leptospires in Abortion, Stillbirths and Neonatal Deaths in Sheep. Vet. Rec. 1983, 112, 291–293. [Google Scholar] [CrossRef]
- Leon-Vizcaino, L.; Hermoso de Mendoza, M.; Garrido, F. Incidence of Abortions Caused by Leptospirosis in Sheep and Goats in Spain. Comp. Immunol. Microbiol. Infect. Dis. 1987, 10, 149–153. [Google Scholar] [CrossRef]
- Pereira, M.H.C.; Cooke, R.F.; Alfieri, A.A.; Vasconcelos, J.L.M. Effects of Vaccination against Reproductive Diseases on Reproductive Performance of Lactating Dairy Cows Submitted to AI. Anim. Reprod. Sci. 2013, 137, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Libonati, H.A.; Santos, G.B.; Souza, G.N.; Brandão, F.Z.; Lilenbaum, W. Leptospirosis Is Strongly Associated to Estrus Repetition on Cattle. Trop. Anim. Health Prod. 2018, 50, 1625–1629. [Google Scholar] [CrossRef]
- Loureiro, A.P.; Lilenbaum, W. Genital Bovine Leptospirosis: A New Look for an Old Disease. Theriogenology 2020, 141, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Lilenbaum, W. Control of Bovine Leptospirosis: Aspects for Consideration in a Tropical Environment. Res. Vet. Sci. 2017, 112, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, R.; Whittier, W.D.; Collins, J.C.; Currin, J.F.; Inman, B.; Hall, J.B.; Pelzer, K.D. A Field Study of the Effects of a Monovalent Leptospira borgpetersenii Serovar Hardjo Strain Hardjobovis Vaccine Administered with Oxytetracycline on Reproductive Performance in Beef Cattle. J. Am. Vet. Med. Assoc. 2007, 231, 1709–1714. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, A.H.; Graham, T.W.; Famula, T.R.; Oberbauer, A.M. Effect of a Monovalent Vaccine against Leptospira borgpetersenii Serovar Hardjo Strain Hardjobovis on Fertility in Holstein Dairy Cattle. J. Am. Vet. Med. Assoc. 2013, 242, 1564–1572. [Google Scholar] [CrossRef] [Green Version]
- Vallée, E.; Heuer, C.; Collins-Emerson, J.M.; Benschop, J.; Ridler, A.L.; Wilson, P.R. Effects of Natural Infection by L. borgpetersenii Serovar Hardjo Type Hardjo-Bovis and L. interrogans Serovar Pomona, and Leptospiral Vaccination, on Sheep Growth. Prev. Vet. Med. 2018, 159, 196–202. [Google Scholar] [CrossRef]
- Vallée, E.; Heuer, C.; Collins-Emerson, J.M.; Benschop, J.; Ridler, A.L.; Wilson, P.R. Effects of Natural Infection by L. borgpetersenii Serovar Hardjo Type Hardjo-Bovis, L. interrogans Serovar Pomona and Leptospiral Vaccination on Sheep Reproduction. Theriogenology 2018, 114, 126–135. [Google Scholar] [CrossRef]
- Bolin, C.A.; Alt, D.P. Use of a Monovalent Leptospiral Vaccine to Prevent Renal Colonization and Urinary Shedding in Cattle Exposed to Leptospira borgpetersenii Serovar Hardjo. Am. J. Vet. Res. 2001, 62, 995–1000. [Google Scholar] [CrossRef]
- 9 CFR 113.105—Leptospira Hardjo Bacterin. Available online: https://www.ecfr.gov/current/title-9/chapter-I/subchapter-E/part-113/subject-group-ECFR275e24cb93d1fab/section-113.105 (accessed on 11 August 2022).
- World Organisation for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 6th ed.; World Organisation for Animal Health: Paris, France, 2012. [Google Scholar]
- Vallée, E.; Heuer, C.; Collins-Emerson, J.M.; Benschop, J.; Wilson, P.R. Serological Patterns, Antibody Half-Life and Shedding in Urine of Leptospira Spp. in Naturally Exposed Sheep. N. Z. Vet. J. 2015, 63, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Mughini-Gras, L.; Bonfanti, L.; Natale, A.; Comin, A.; Ferronato, A.; La Greca, E.; Patregnani, T.; Lucchese, L.; Marangon, S. Application of an Integrated Outbreak Management Plan for the Control of Leptospirosis in Dairy Cattle Herds. Epidemiol. Infect. 2014, 142, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Loureiro, A.P.; Libonati, H.; Lilenbaum, W. Humoral Response in Naturally Exposed Horses After Leptospiral Vaccination. J. Equine Vet. Sci. 2017, 57, 24–28. [Google Scholar] [CrossRef]
- Martins, G.; Oliveira, C.S.; Lilenbaum, W. Dynamics of Humoral Response in Naturally-Infected Cattle after Vaccination against Leptospirosis. Acta Trop. 2018, 187, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Dorjee, S.; Heuer, C.; Jackson, R.; West, D.M.; Collins-Emerson, J.M.; Midwinter, A.C.; Ridler, A.L. Are White-Spot Lesions in Kidneys in Sheep Associated with Leptospirosis? N. Z. Vet. J. 2009, 57, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Sanhueza, J.M.; Wilson, P.R.; Benschop, J.; Collins-Emerson, J.M.; Heuer, C. Meta-Analysis of the Efficacy of Leptospira Serovar Hardjo Vaccines to Prevent Urinary Shedding in Cattle. Prev. Vet. Med. 2018, 153, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, M.J.; Koopmans, M.J.; Olyhoek, T. Effect of Streptomycin Treatment on the Shedding of and the Serologic Responses to Leptospira interrogans Serovar Hardjo Subtype Hardjobovis in Experimentally Infected Cows. Vet. Microbiol. 1993, 38, 129–135. [Google Scholar] [CrossRef]
- Bolin, C.A.; Thiermann, A.B.; Handsaker, A.L.; Foley, J.W. Effect of Vaccination with a Pentavalent Leptospiral Vaccine on Leptospira interrogans Serovar Hardjo Type Hardjo-Bovis Infection of Pregnant Cattle. Am. J. Vet. Res. 1989, 50, 161–165. [Google Scholar]
- Putz, E.J.; Nally, J.E. Investigating the Immunological and Biological Equilibrium of Reservoir Hosts and Pathogenic Leptospira: Balancing the Solution to an Acute Problem? Front. Microbiol. 2020, 11, 2005. [Google Scholar] [CrossRef]
- Bolin, C.A.; Cassells, J.A.; Zuerner, R.L.; Trueba, G. Effect of Vaccination with a Monovalent Leptospira interrogans Serovar Hardjo Type Hardjo-Bovis Vaccine on Type Hardjo-Bovis Infection of Cattle. Am. J. Vet. Res. 1991, 52, 1639–1643. [Google Scholar]



{kind=link}
{kind=link}
| DAY | GROUP | SEROGROUPS | ||||
|---|---|---|---|---|---|---|
| Australis | Canicola | Icterohaemorrhagiae | Pomona | Sejroe | ||
| D-35 | A (commercial vaccine) | 0% | 0% | 0% | 0% | 0% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 0% | |
| C (control) | 0% | 0% | 0% | 0% | 0% | |
| D-14 * | A (commercial vaccine) | 0% | 0% | 0% | 16.7% | 66.7% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 83.3% | |
| C (control) | 0% | 0% | 0% | 0% | 0% | |
| D0 * | A (commercial vaccine) | 0% | 0% | 0% | 16.7% | 66.7% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 83.3% | |
| C (control) | 0% | 0% | 0% | 0% | 0% | |
| D30 | A (commercial vaccine) | 0% | 0% | 0% | 0% | 33.3% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 16.7% | |
| C (control) | 0% | 0% | 0% | 0% | 100% | |
| D60 * | A (commercial vaccine) | 0% | 0% | 0% | 0% | 50% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 16.7% | |
| C (control) | 16.7% | 0% | 0% | 0% | 33.3% | |
| D90 | A (commercial vaccine) | 0% | 0% | 0% | 0% | 16.7% |
| B (FV52 bacterin) | 16.7% | 0% | 0% | 0% | 16.7% | |
| C (control) | 0% | 0% | 0% | 0% | 0% | |
| D120 | A (commercial vaccine) | 16.7% | 0% | 0% | 0% | 33.3% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 16.7% | |
| C (control) | 0% | 0% | 0% | 0% | 33.3% | |
| D150 | A (commercial vaccine) | 0% | 0% | 0% | 0% | 16.7% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 0% | |
| C (control) | 0% | 0% | 0% | 0% | 16.7% | |
| D180 | A (commercial vaccine) | 0% | 0% | 0% | 0% | 0% |
| B (FV52 bacterin) | 0% | 0% | 0% | 0% | 0% | |
| C (control) | 0% | 0% | 0% | 0% | 16.7% | |
| Group A (Commercial Vaccine) | Group B (FV52 Bacterin) | Group C (Control) | |
|---|---|---|---|
| D-35 | 0% | 0% | 0% |
| D-14 | 0% | 0% | 0% |
| D0 | 0% | 0% | 0% |
| D30 PI | 66.7% | 66.7% | 50% |
| D60 PI | 66.7% | 33.3% | 0% |
| D90 PI * | 0% | 66.7% | 83.3% |
| D120 PI | 0% | 0% | 0% |
| D150 PI | 0% | 0% | 0% |
| D180 PI | 0% | 0% | 0% |
| D210 PI | 0% | 0% | 0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, G.; Guadelupe, B.; Aymée, L.; Balaro, M.F.A.; Pinto, P.H.; Di Azevedo, M.I.N.; Brandão, F.Z.; Lilenbaum, W. The Efficacy of Vaccination in the Prevention of Renal and Genital Leptospirosis in Experimentally Infected Sheep. Trop. Med. Infect. Dis. 2022, 7, 321. https://doi.org/10.3390/tropicalmed7100321
Martins G, Guadelupe B, Aymée L, Balaro MFA, Pinto PH, Di Azevedo MIN, Brandão FZ, Lilenbaum W. The Efficacy of Vaccination in the Prevention of Renal and Genital Leptospirosis in Experimentally Infected Sheep. Tropical Medicine and Infectious Disease. 2022; 7(10):321. https://doi.org/10.3390/tropicalmed7100321
Chicago/Turabian StyleMartins, Gabriel, Bruna Guadelupe, Luiza Aymée, Mario Felipe Alvarez Balaro, Pedro Henrique Pinto, Maria Isabel Nogueira Di Azevedo, Felipe Zandonadi Brandão, and Walter Lilenbaum. 2022. "The Efficacy of Vaccination in the Prevention of Renal and Genital Leptospirosis in Experimentally Infected Sheep" Tropical Medicine and Infectious Disease 7, no. 10: 321. https://doi.org/10.3390/tropicalmed7100321
APA StyleMartins, G., Guadelupe, B., Aymée, L., Balaro, M. F. A., Pinto, P. H., Di Azevedo, M. I. N., Brandão, F. Z., & Lilenbaum, W. (2022). The Efficacy of Vaccination in the Prevention of Renal and Genital Leptospirosis in Experimentally Infected Sheep. Tropical Medicine and Infectious Disease, 7(10), 321. https://doi.org/10.3390/tropicalmed7100321