
Intestinal Parasitic Infection and Nutritional Status in Children under Five Years Old: A Systematic Review



Abstract
:1. Introduction
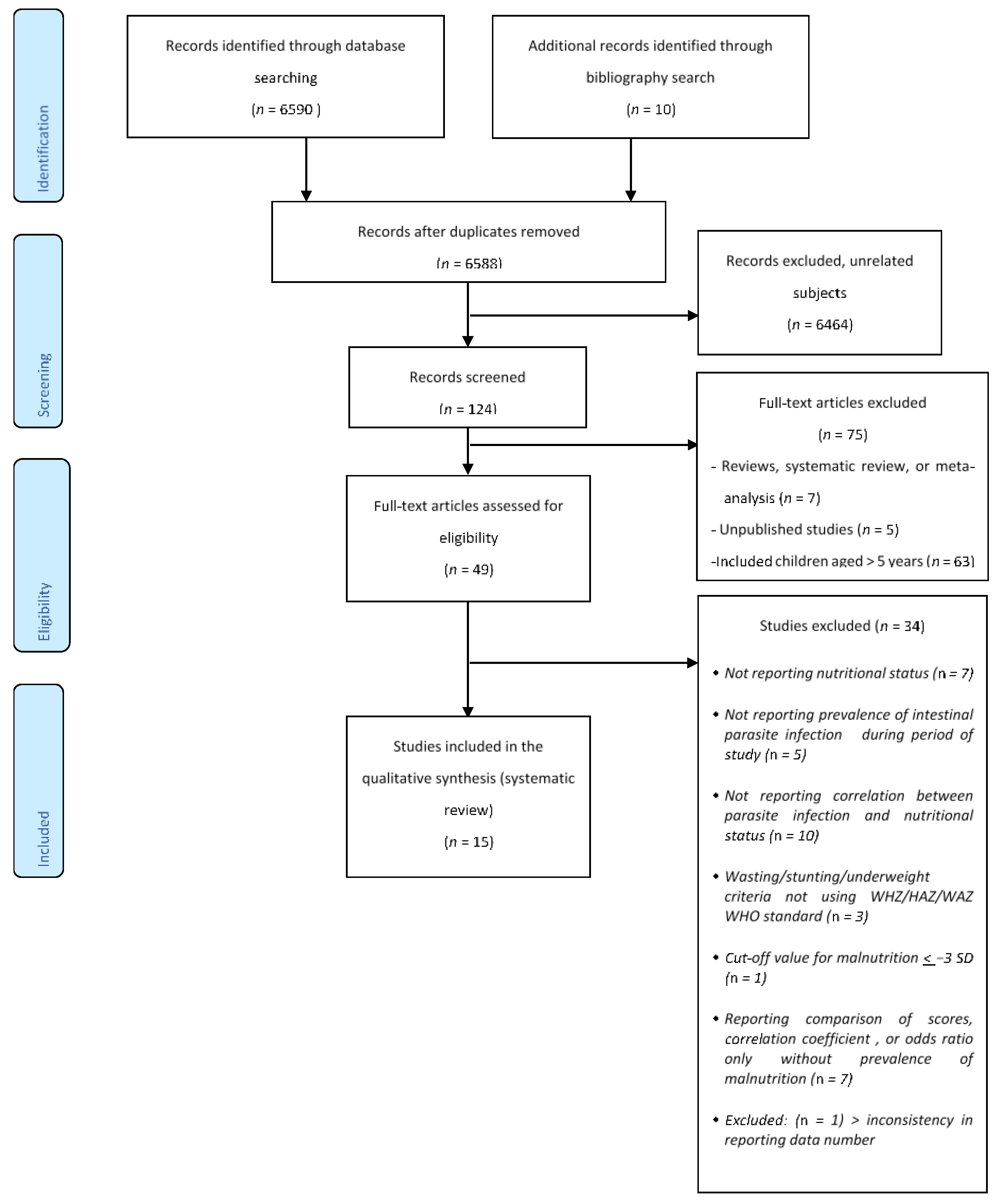
2. Materials and Methods
2.1. Literature Search and Identification
2.2. Inclusion and Exclusion Criteria
2.3. Data Collection and Analysis
2.4. Outcome Variables
2.5. Statistical Analysis
3. Results
3.1. Type of Parasite and Geographical Distribution
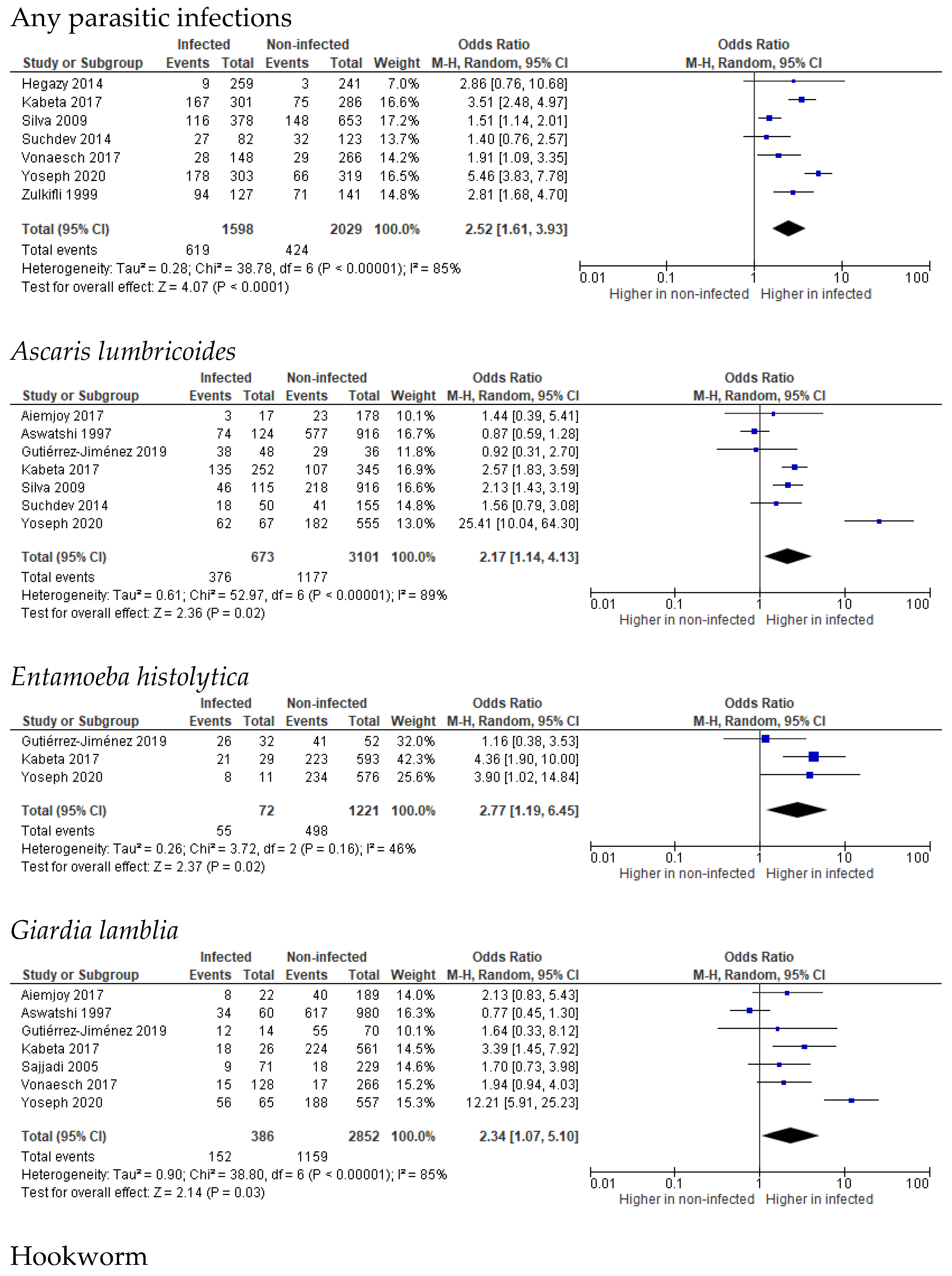
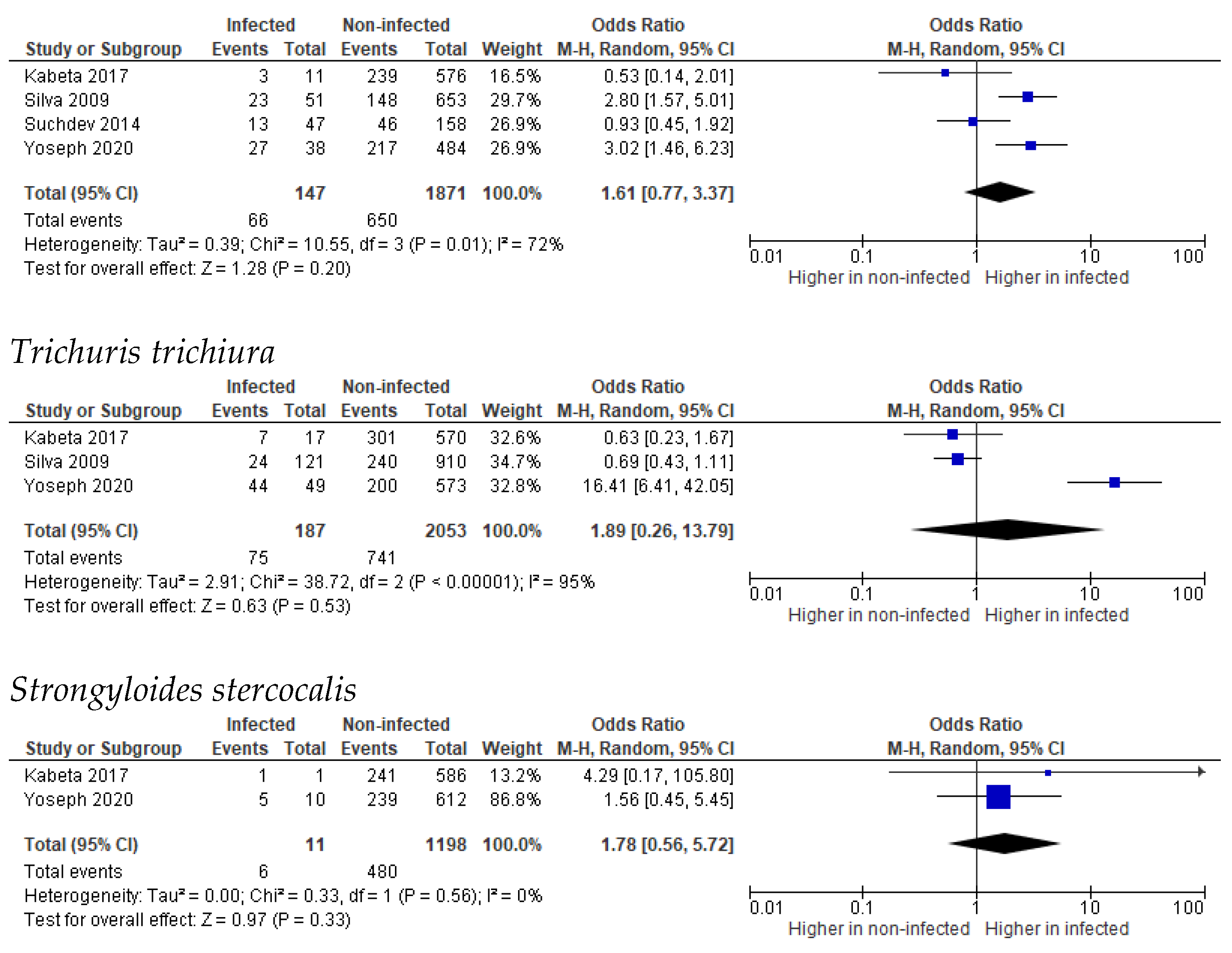
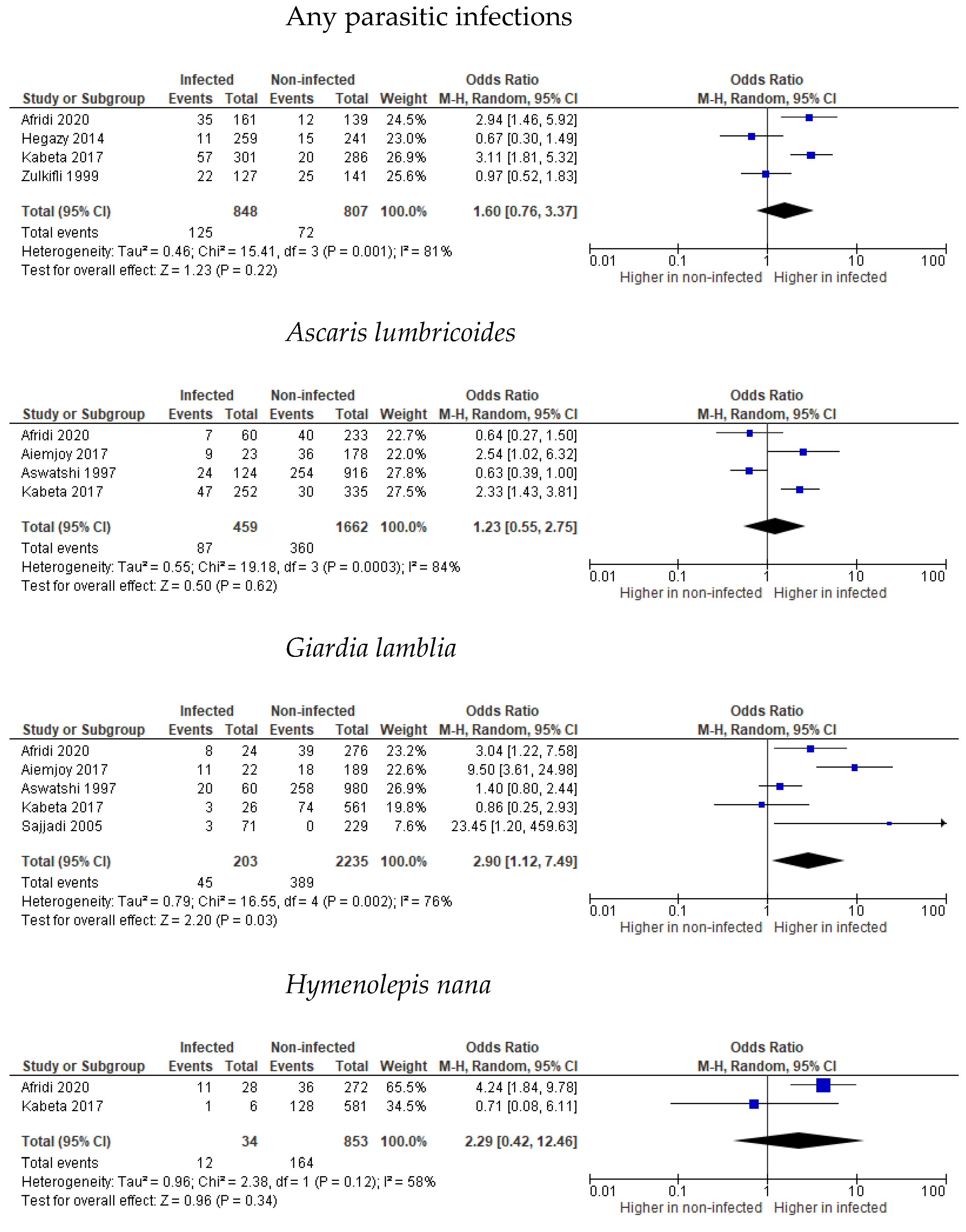
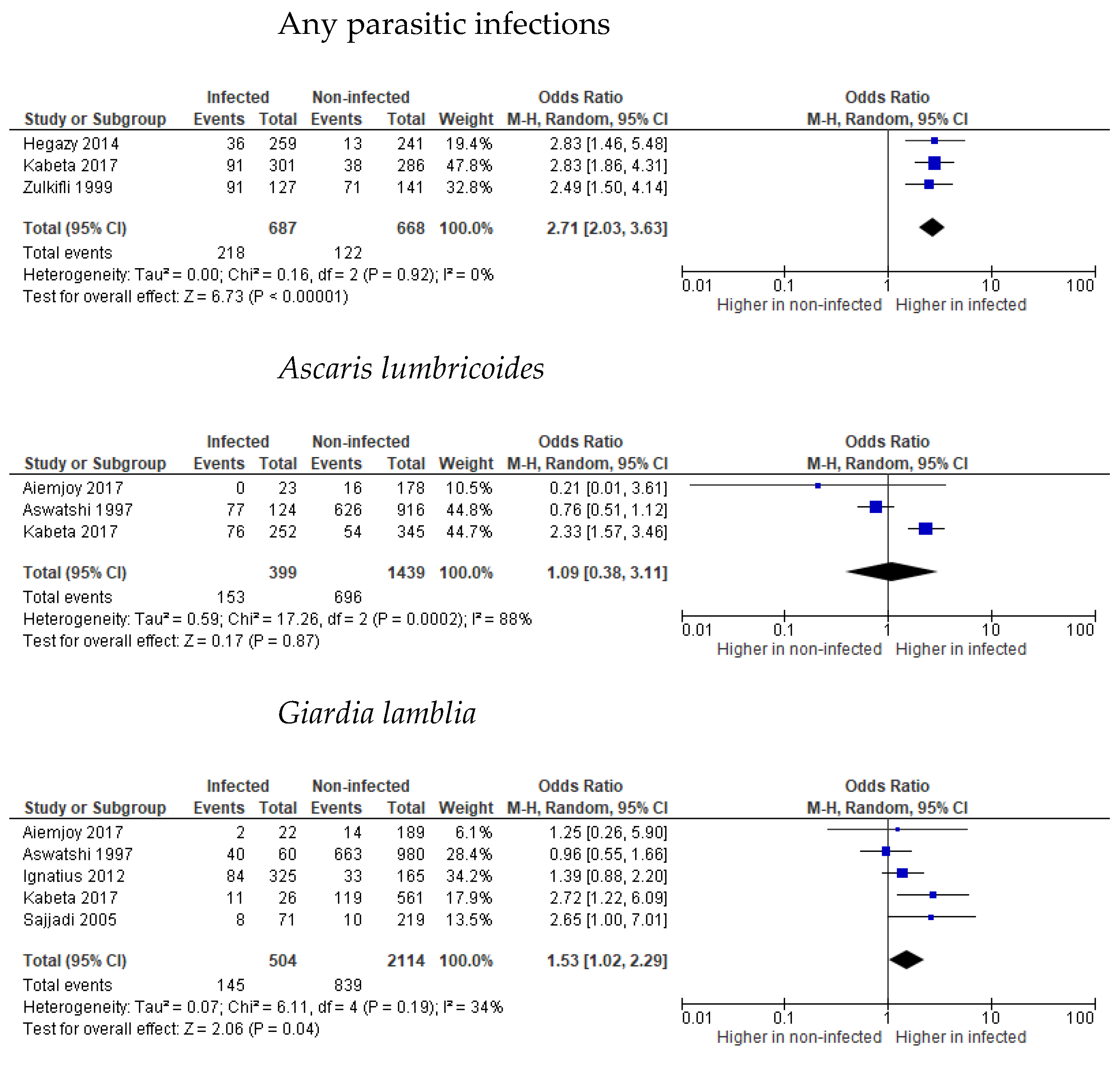
3.2. Stunting
3.3. Wasting
3.4. Underweight
3.5. Undernutrition
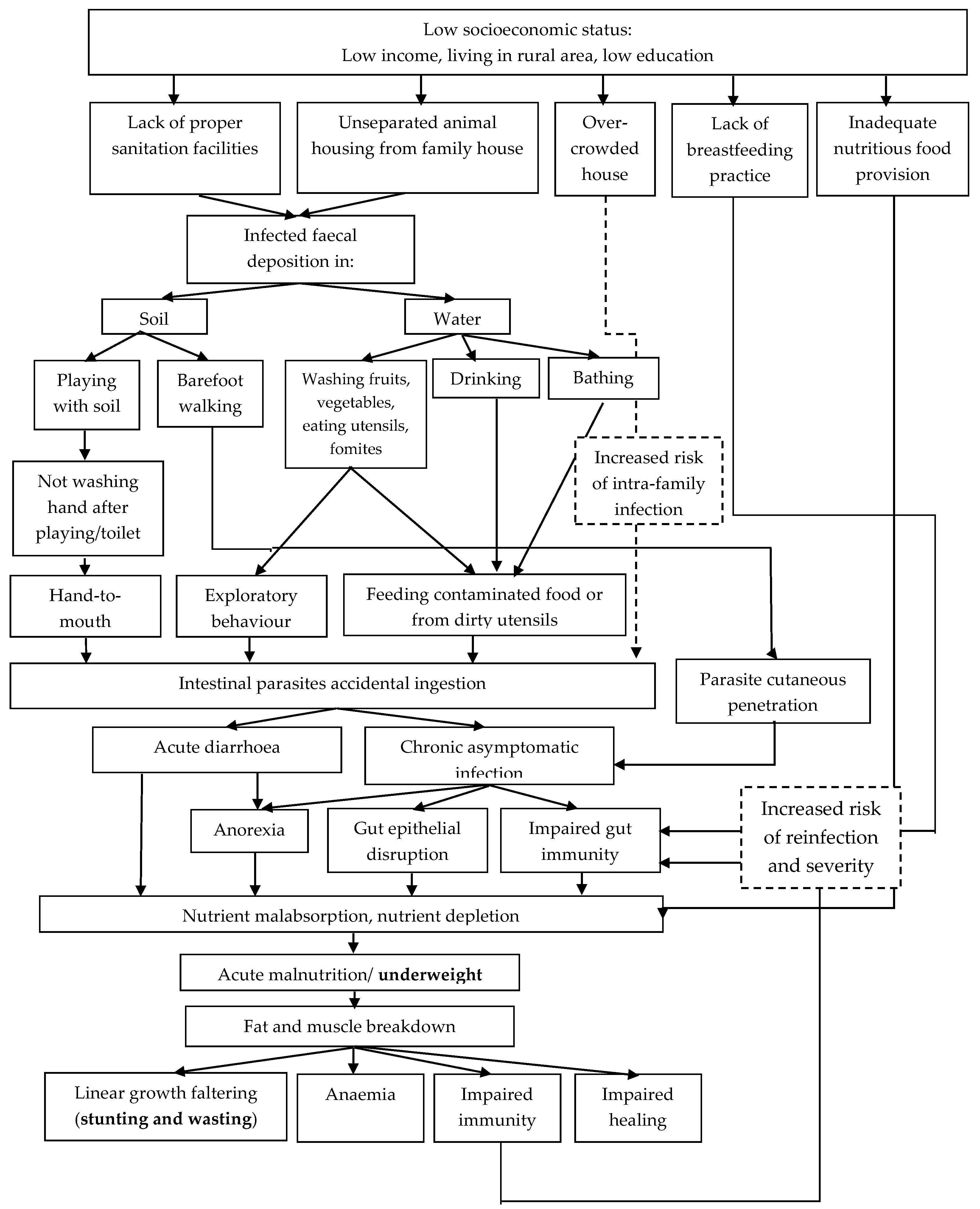
3.6. Risk Factors Related to Parasitic Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tigabu, A.; Taye, S.; Aynalem, M.; Adane, K. Prevalence and associated factors of intestinal parasitic infections among patients attending Shahura Health Center, Northwest Ethiopia. BMC Res. Notes 2019, 12, 333. [Google Scholar] [CrossRef] [PubMed]
- Belete, Y.A.; Kassa, T.Y.; Baye, M.F. Prevalence of intestinal parasite infections and associated risk factors among patients of Jimma health center requested for stool examination, Jimma, Ethiopia. PLoS ONE 2021, 16, e0247063. [Google Scholar] [CrossRef] [PubMed]
- Getachew, T.; Argaw, A. Intestinal helminth infections and dietary diversity score predict nutritional status of urban schoolchildren from souther Ethiopia. BMC Nutr. 2017, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Zemene, T.; Shiferaw, M.B. Prevalence of intestinal parasitic infections in children under the age of 5 years attending the Debre Birhan referral hospital, North Shoa, Ethiopia. BMC Res. Notes 2018, 11, 58. [Google Scholar] [CrossRef]
- Mardu, F.; Yohannes, M.; Tadesse, D. Prevalence of intestinal parasites and associated risk factors among inmates of Mekelle prison, Tigrai Region, Northern Ethiopia, 2017. BMC Infect. Dis. 2019, 19, 406. [Google Scholar] [CrossRef]
- World Health Organization. Global Distribution and Prevalence of Soil Transmitted Helminth Infections; World Health Organization Key Fact Sheet; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Jourdan, P.M.; Lamberton, P.H.L.; Fenwick, A.; Addiss, D.G. Soil-transmitted helminth infections. Lancet 2018, 391, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Montresor, A.; Mupfasoni, D.; Mikhailov, A.; Mwinzi, P.; Lucianez, A.; Jamsheed, M.; Gasimov, E.; Warusavithana, S.; Yajima, A.; Bisoffi, Z.; et al. The global progress of soil-transmitted helminthiases control in 2020 and World Health Organization targets for 2030. PLoS Negl. Trop. Dis. 2020, 14, e0008505. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.-N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 11 Foodborne Parasitic Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, Z.; Karim, R.; Zhang, L. Detection of human intestinal protozoan parasites in vegetables and fruits: A review. Parasites Vectors 2020, 13, 380. [Google Scholar] [CrossRef]
- Keiser, J.; Utzinger, J. Food-Borne Trematodiases. Clin. Microbiol. Rev. 2009, 22, 466–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, G.L. Food-borne protozoa. Br. Med. Bull. 2000, 56, 209–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantzanou, M.; Karalexi, M.A.; Vrioni, G.; Tsakris, A. Prevalence of Intestinal Parasitic Infections among Children in Europe over the Last Five Years. Trop. Med. Infect. Dis. 2021, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Curico, G.; García-Bardales, P.; Pinedo, T.; Shapiama, W.; Moncada-Yaicate, M.; Romaina, L.; Yori, P.P.; Paredes-Olortegui, M.; Meza-Sánchez, G.; Lescano, A.G.; et al. Resistance to single dose albendazole and reinfection with intestinal helminths among children ages 2 to 11 years from the Peruvian Amazon region: A study protocol. BMC Infect. Dis. 2022, 22, 528. [Google Scholar] [CrossRef] [PubMed]
- Woldegerima, E.; Bayih, A.G.; Tegegne, Y.; Aemero, M.; Zeleke, A.J. Prevalence and Reinfection Rates of Schistosoma mansoni and Praziquantel Efficacy against the Parasite among Primary School Children in Sanja Town, Northwest Ethiopia. J. Parasitol. Res. 2019, 2019, 3697216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gungoren, B.; Latipov, R.; Regallet, G.; Musabaev, E. Effect of hygiene promotion on the risk of reinfection rate of intestinal parasites in children in rural Uzbekistan. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 564–569. [Google Scholar] [CrossRef]
- World Health Organization. Malnutrition. 9 June 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/malnutrition (accessed on 10 November 2021).
- DiPasquale, V.; Cucinotta, U.; Romano, C. Acute Malnutrition in Children: Pathophysiology, Clinical Effects and Treatment. Nutrients 2020, 12, 2413. [Google Scholar] [CrossRef]
- Bouma, S. Diagnosing Pediatric Malnutrition. Nutr. Clin. Pract. 2016, 32, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Elia, M. Defining, Recognizing, and Reporting Malnutrition. Int. J. Low. Extremity Wounds 2017, 16, 230–237. [Google Scholar] [CrossRef]
- Cederholm, T.; Barazzoni, R.; Austin, P.; Ballmer, P.; Biolo, G.; Bischoff, S.C.; Compher, C.; Correia, I.; Higashiguchi, T.; Holst, M.; et al. ESPEN guidelines on definitions and terminology of clinical nutrition. Clin. Nutr. 2017, 36, 49–64. [Google Scholar] [CrossRef]
- Ereinhardt, K.; Fanzo, J. Addressing Chronic Malnutrition through Multi-Sectoral, Sustainable Approaches: A Review of the Causes and Consequences. Front. Nutr. 2014, 1, 13. [Google Scholar] [CrossRef]
- United Nations International Children’s Emergency Fund. Undernutrition. UNICEF Data: Monitoring the situation of children and women. April 2021. Available online: https://data.unicef.org/topic/nutrition/malnutrition/ (accessed on 10 November 2021).
- Dobner, J.; Kaser, S. Body mass index and the risk of infection-from underweight to obesity. Clin. Microbiol. Infect. 2017, 24, 24–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomons, N.W. Malnutrition and Infection: An Update. Br. J. Nutr. 2007, 98 (Suppl. S1), S5–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katona, P.; Katona-Apte, J. The Interaction between Nutrition and Infection. Clin. Infect. Dis. 2008, 46, 1582–1588. [Google Scholar] [CrossRef]
- Simon, A.K.; Hollander, G.A.; McMichael, A. Evolution of the immune system in humans from infancy to old age. Proc. R. Soc. B Biol. Sci. 2015, 282, 20143085. [Google Scholar] [CrossRef] [Green Version]
- Schaible, U.E.; Kaufmann, S.H. Malnutrition and Infection: Complex Mechanisms and Global Impacts. PLoS Med. 2007, 4, e115. [Google Scholar] [CrossRef] [Green Version]
- Ygberg, S.; Nilsson, A. The developing immune system-from foetus to toddler. Acta Paediatr. 2011, 101, 120–127. [Google Scholar] [CrossRef]
- Wood, A.C.; Blissett, J.M.; Burnstorm, J.M.; Carnell, S.; Faith, M.S.; Fisher, J.O.; Hayman, L.L.; Khalsa, A.S.; Hughes, S.O.; Miller, A.L.; et al. Caregiver influences on eating behaviours in young children. J. Am. Heart Assoc. 2020, 9, e014520. [Google Scholar] [CrossRef]
- White, E.J.; Hutka, S.A.; Williams, L.J.; Emoreno, S. Learning, neural plasticity and sensitive periods: Implications for language acquisition, music training and transfer across the lifespan. Front. Syst. Neurosci. 2013, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Sustainable Development Solutions Network. Indicators and a Monitoring Framework. 2012. Available online: https://indicators.report/targets/2-2/ (accessed on 14 November 2021).
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gotzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statementfor reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef]
- Moola, S.; Munn, Z.; Tufanaru, C.; Aromataris, E.; Sears, K.; Sfetcu, R.; Currie, M.; Qureshi, R.; Mattis, P.; Lisy, K.; et al. Chapter 7: Systematic reviews of etiology and risk. In JBI Manual for Evidence Synthesis. JBI; Aromataris, E., Munn, Z., Eds.; JBI Global: Adelaide, Australia, 2020; Available online: https://synthesismanual.jbi.global (accessed on 14 November 2021).
- Aiemjoy, K.; Gebresillasie, S.; Stoller, N.E.; Shiferaw, A.; Tadesse, Z.; Chanyalew, M.; Aragie, S.; Callahan, K.; Keenan, J.D. Epidemiology of soil-transmitted helminth and intestinal protozonal infections in preschool-aged children in the Amhara region of Ethiopia. Am. J. Trop. Med. Hyg. 2017, 96, 866–872. [Google Scholar] [PubMed] [Green Version]
- Gutiérrez-Jiménez, J.; Luna-Cázares, L.M.; La Cruz, L.M.-D.; De Aquino-López, J.A.; Sandoval-Gómez, D.; León-Ortiz, A.T.; Hernández-Shilón, J.A.; Constantino-Jonapa, L.A.; Matamoros, W.A.; Vidal, J.E. Children from a rural region in The Chiapas Highlands, Mexico, show an increased risk of stunting and intestinal parasitoses when compared with urban children. Bol. Med. Hosp. Infant Mex. 2019, 76. [Google Scholar] [CrossRef]
- Haratipour, H.; Sohrabi, M.B.; Zolfaghari, P.; Nezakati, E.; Yahyaei, E.; Rezvani, S. The Relationship between Malnutrition and Intestinal Parasitic Infections among Preschool Children in East Area of Iran. Int. J. Pediatr. 2016, 4, 2011–2018. [Google Scholar] [CrossRef]
- Hegazy, A.M.; Tawakol, N.; Aminou, H.; Badr, A.M. Prevalence of intestinal parasites and its impact on nutritional status among preschool children living in Damanhur City El-Behera Governorate, Egypt. J. Egypt Soc. Parasitol. 2014, 44, 517–524. [Google Scholar]
- Vonaesch, P.; Tondeur, L.; Breurec, S.; Bata, P.; Nguyen, L.B.L.; Frank, T.; Ferra, A.; Rafaï, C.; Giles-Vernick, T.; Gody, J.C.; et al. Factors associated with stunting in healthy children aged 5 years and less living in Bangui (RCA). PLoS ONE 2017, 12, e0182363. [Google Scholar] [CrossRef] [Green Version]
- Yoseph, A.; Beyene, H. The high prevalence of intestinal parasitic infections in associated with stunting among children aged 6-59 months in Boricha Woreda, Southern Ethiopia: A cross-sectional study. BMC Public Health 2020, 20, 1270. [Google Scholar] [CrossRef]
- Afridi, M.F.; Farhat, K.; Ali, S.; Qaisrani, M.N.; Ahmed, Z. Prevalence of intestinal parasitic infestations in relation to wasting among children under 5 years of age in Skardu, Pakistan: A cross sectional observational study. Isra Med. J. 2021, 13, 130–133. [Google Scholar]
- Awasthi, S.; Pande, V.K. Prevalence of malnutrition and intestinal parasites in preschool slum children in Lucknow. Indian Pediatr. 1997, 34, 599–605. [Google Scholar]
- Ignatius, R.; Gahutu, J.B.; Klotz, C.; Steininger, C.; Shyirambere, C.; Lyng, M.; Musemakweri, A.; Aebischer, T.; Martus, P.; Harms, G.; et al. High Prevalence of Giardia duodenalis Assemblage B Infection and Association with Underweight in Rwandan Children. PLoS Negl. Trop. Dis. 2012, 6, e1677. [Google Scholar] [CrossRef]
- Kabeta, A.; Assefa, S.; Hailu, D.; Berhanu, G. Intestinal parasitic infections and nutritional status of pre-school children in Hawassa Zuria Distrtic, South Ethiopia. Afr. J. Microbiol. Res. 2017, 11, 1243–1251. [Google Scholar]
- Sajadi, S.M.; Tanideh, M. Nutritional status of preschool children infected with Giardia intestinalis. Iran. J Public Health 2005, 34, 51–57. [Google Scholar]
- Silva, R.R.; Da Silva, C.A.M.; Pereira, C.A.; Nicolato, R.L.D.C.; Negrão-Corrêa, D.; Lamounier, J.A.; Carneiro, M. Association between nutritional status, environmental and socio-economic factors and Giardia lamblia infections among children aged 6–71 months in Brazil. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Suchdev, P.S.; Davis, S.M.; Bartoces, M.; Ruth, L.J.; Worrell, C.M.; Kanyi, H.; Odero, K.; Wiegand, R.E.; Njenga, S.M.; Montgomery, J.M.; et al. Soil-transmitted helminth infection and nutritional stats among urban slum children in Kenya. Am. J. Trop Med. Hyg. 2014, 90, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulkifli, A.; Khairul, A.A.; Atiya, A.S.; Yano, A. Malnutrition and helminth infections among pre-school children in orang asli resettlement villages in Kelantan. JUMMEC 1999, 2, 99–103. [Google Scholar]
- Doni, N.Y.; Zeyrek, F.Y.; Simsek, Z.; Gurges, G.; Sahin, I. Growth retardation and psychomotor development delays of children in Sanhurfa, Turkey. Turk. Parazitol. Derg. 2015, 39, 270–276. [Google Scholar] [CrossRef]
- Omitola, O.; Mogaji, H.O.; Oluwole, A.; Adeniran, A.A.; Alabi, O.M.; Ekpo, U. Geohelminth Infections and Nutritional Status of Preschool Aged Children in a Periurban Settlement of Ogun State. Scientifica 2016, 2016, 7897351. [Google Scholar] [CrossRef]
- Waterlow, J.C. Summary of causes and mechanisms of linear growth retardation. Eur. J. Clin. Nutr. 1994, 48 (Suppl. S1), S210–S211. [Google Scholar]
- LaBeaud, A.D.; Singer, M.N.; McKibben, M.; Mungai, P.; Muchiri, E.M.; McKibben, E.; Gildengorin, G.; Sutherland, L.J.; King, C.H.; King, C.L.; et al. Parasitism in Children Aged Three Years and Under: Relationship between Infection and Growth in Rural Coastal Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003721. [Google Scholar] [CrossRef]
- Fauziah, N.; Ar-Rizqi, M.A.; Hana, S.; Patahuddin, N.M.; Diptyanusa, A. Stunting as a Risk Factor of Soil-Transmitted Helminthiasis in Children: A Literature Review. Interdiscip. Perspect. Infect. Dis. 2022, 2022, 8929025. [Google Scholar] [CrossRef]
- Dold, C.; Holland, C.V. Ascaris and ascariasis. Microbes Infect. 2011, 13, 632–637. [Google Scholar] [CrossRef]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyemo, F.E.; Singh, G.; Reddy, P.; Bux, F.; Stenström, T.A. Efficiency of chlorine and UV in the inactivation of Cryptosporidium and Giardia in wastewater. PLoS ONE. 2019, 14, e0216040. [Google Scholar] [CrossRef] [PubMed]
- Loukas, A.; Hotes, P.J.; Diemert, D.; Yazdanbaksh, M.; Mccarty, J.S.; Correa-Oliveira, R.; Croese, J.; Bethony, J.M. Affiliations expand. Hookworm infection. Nat. Rev. 2016, 2, 16088. [Google Scholar]
- Jiraanankul, V.; Aphijirawat, W.; Piyaraj, P.; Leelayoova, S.; Khositnithikul, R.; Taamasri, P.; Traub, R.J.; Mungthin, M.; Naaglor, T.; Rangsin, R. Incidence and Risk Factors of Hookworm Infection in a Rural Community of Central Thailand. Am. J. Trop. Med. Hyg. 2011, 84, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Else, K.J.; Keiser, J.; Holland, C.V.; Grencis, R.K.; Sattelle, D.B.; Fujiwara, R.T.; Bueno, L.L.; Asaolu, S.O.; Sowemimo, O.A.; Cooper, P.J. Whipworm and roundworm infections. Nat. Rev. Dis. Prim. 2020, 6, 44. [Google Scholar] [CrossRef]
- Kantor, M.; Abrantes, A.; Estevez, A.; Schiller, A.; Torrent, J.; Gascon, J.; Hernandez, R.; Ochner, C. Entamoeba Histolytica: Updates in Clinical Manifestation, Pathogenesis, and Vaccine Development. Can. J. Gastroenterol. Hepatol. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Hymenolepiasis. DPDx-Laboratory Identificatin of Parasites of Public Health Concern. 2017. Available online: https://www.cdc.gov/dpdx/hymenolepiasis/index.html (accessed on 25 October 2022).
- Cook, G.C. Enterobius vermicularis infection. Gut 1994, 35, 1159–1162. [Google Scholar] [CrossRef] [Green Version]
- Hossain, S.; Das, S.; Gazi, A.; Mahfuz, M.; Ahmed, T. Ascaris lumbricoides infection: Still a threat for iron deficiency anaemia in 2-year-old Bangladeshi slum-dwelling children. J. Infect. Dev. Ctries. 2019, 13, 933–938. [Google Scholar] [CrossRef]
- Ngui, R.; Lim, Y.; Kin, L.C.; Chuen, C.S.; Jaffar, S. Association between Anaemia, Iron Deficiency Anaemia, Neglected Parasitic Infections and Socioeconomic Factors in Rural Children of West Malaysia. PLoS Negl. Trop. Dis. 2012, 6, e1550. [Google Scholar] [CrossRef]
- Schwarz, A.; Gauly, M.; Abel, H.; Daş, G.; Humburg, J.; Rohn, K.; Breves, G.; Rautenschlein, S. Immunopathogenesis of Ascaridia galli infection in layer chicken. Dev. Comp. Immunol. 2011, 35, 774–784. [Google Scholar] [CrossRef]
- Koehler, S.; Springer, A.; Issel, N.; Klinger, S.; Wendt, M.; Breves, G.; Strube, C. Ascaris suum Nutrient Uptake and Metabolic Release, and Modulation of Host Intestinal Nutrient Transport by Excretory-Secretory and Cuticle Antigens In Vitro. Pathogens 2021, 10, 1419. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, L.; Latham, M.; Ottesen, E. Malnutrition and parasitic helminth infections. Parasitology 2000, 121, S23–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelt, L.A.; Sartor, R.B. Advances in understanding Giardia: Determinants and mechanisms of chronic sequelae. F1000Prime Rep. 2015, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burret, A.G.; Cotton, J. Pathophysiological processes and clinical manifestations of giardiasis. In Giardia: A Model Organism; Lujan, H.D., Svard, S., Eds.; Springer: Wien, Austria, 2011; p. 309. [Google Scholar]
- Gyorkos, T.W.; Gilbert, N.L.; Larocque, R.; Casapía, M.; Montresor, A. Re-Visiting Trichuris trichiura Intensity Thresholds Based on Anemia during Pregnancy. PLoS Negl. Trop. Dis. 2012, 6, e1783. [Google Scholar] [CrossRef]
- Celiksoz, A.; Acioz, M.; Degerli, S.; Oztop, A.Y.; Alim, A. Effect of Enterobiasis on primary school children. Afr. J. Microbiol. Res. 2010, 4, 634–639. [Google Scholar]
- Al-Daoody, A.A.K.; Al-Bazzaz, E.N.H. Impact of Enterobius vermicularis infection on biochemical parameters in the blood of children in Erbil Province, Iraq. BMC Infect Dis. 2020, 20, 336. [Google Scholar] [CrossRef]
- Tsuyuoka, R.; Bailey, J.W.; Guimaraes, A.M.D.N.; Gurgel, R.Q.; Cuevas, L.E. Anemia and intestinal parasitic infections in primary school students in Aracaju, Sergipe, Brazil. Cad. Saúde Pública 1999, 15, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Al Qadhi, B.N.; Al-Warid, H.S.J. Anemia and enterobiasis among Iraqi children. J. Coll. B Educ. 2012, 76, 57–62. [Google Scholar]
- Budge, S.; Parker, A.H.; Hutchings, P.T.; Garbutt, C. Environmental enteric dysfunction and child stunting. Nutr. Rev. 2019, 77, 240–253. [Google Scholar] [CrossRef] [Green Version]
- Alberda, C.; Graf, A.; McCargar, L. Malnutrition: Etiology, consequences, and assessment of a patient at risk. Best Clin. Pract. Res. Clin. Gastroenterol. 2006, 20, 419–439. [Google Scholar] [CrossRef]
- Thurstans, S.; Sessions, N.; Dolan, C.; Sadler, K.; Cichon, B.; Isanaka, S.; Roberfroid, D.; Stobaugh, H.; Webb, P.; Khara, T. The relationship between wasting and stunting in young children: A systematic review. Matern. Child Nutr. 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Briend, A.; Akomo, P.; Bahwere, P.; De Pee, S.; DiBari, F.; Golden, M.H.; Manary, M.; Ryan, K. Developing Food Supplements for Moderately Malnourished Children: Lessons Learned from Ready-to-Use Therapeutic Foods. Food Nutr. Bull. 2015, 36 (Suppl. S1), S53–S58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forson, A.O.; Arthur, I.; Ayeh-Kumi, P.F. The role of family size, employment and education of parents in the prevalence of intestinal parasitic infections in school children in Accra. PLoS ONE 2018, 13, e0192303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutty, P.K. Breastfeeding and risk of parasitic infection-a review. Asian Pac. J. Trop. Biomed. 2014, 4, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Nhambirre, O.L.; Cossa-Moiane, I.; Bauhofer, A.F.L.; Chissaque, A.; Lobo, M.L.; Matos, O.; de Deus, N. Intestinal Parasites in Children up to 14 Years Old Hospitalized with Diarrhea in Mozambique, 2014–2019. Pathogens 2022, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Conterno, L.O.; Turchi, M.D.; Corrêa, I.; Almeida, R.A.M.D.B. Anthelmintic drugs for treating ascariasis. Cochrane Database Syst. Rev. 2020, 2020, CD010599. [Google Scholar] [CrossRef] [PubMed]
- Welch, V.A.; Ghogomu, E.; Hossain, A.; Awasthi, S.; Bhutta, Z.; Cumberbatch, C.; Fletcher, R.; McGowan, J.; Krishnaratne, S.; Kristjansson, E.; et al. Deworming and adjuvant interventions for improving the developmental health and well-being of children in low- and middle-income countries: A systematic review and network meta-analysis. Campbell Syst. Rev. 2016, 12, 1–383. [Google Scholar] [CrossRef]
- Betancourt, W.Q.; Rose, J.B. Drinking water treatment processes for removal of Cryptosporidium and Giardia. Veter Parasitol. 2004, 126, 219–234. [Google Scholar] [CrossRef]
- Gera, T.; Shah, D.; Sachdev, H.S. Impact of Water, Sanitation and Hygiene Interventions on Growth, Non-diarrheal Morbidity and Mortality in Children Residing in Low- and Middle-income Countries: A Systematic Review. Indian Pediatr. 2018, 55, 381–393. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite | Study (Year) Country | Stunting (HAZ < −2SD) | Wasting (WHZ < −2SD) | Underweight (WAZ < −2SD) | Undernutrition (HAZ, WHZ, WAZ < −2SD) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Infected | Non-Infected | Crude OR (95% CI), p-Value | Adjusted OR (95% CI), p-Value | Infected | Non-Infected | Crude OR (95% CI), p-Value | Adjusted OR (95% CI), p-Value | Infected | Non-Infected | Crude OR (95% CI), p-Value | Adjusted OR (95% CI), p-Value | Infected | Non-Infected | Crude OR (95% CI), p-Value | Adjusted OR (95% CI), p-Value | ||
| All (not specified) | Doni (2015) [50] Şanlıurfa, Turkey | 42/58 | 19/50 | 4.28 (1.90, 0.0004 | N/A | ||||||||||||
| Haratipour (2016) [38] Shahroud, Iran | 87/649 | 68/1201 | 2.58 (1.85, 3.60), <0.00001 | N/A | |||||||||||||
| Hegazy (2014) [39] Damanhur City El-Behera Governorate, Egypt | 9/259 | 3/241 | 2.98 (0.80, 11.12), 0.100 | N/A | 11/259 | 15/241 | 0.70 (0.31, 1.55), 0.38 | N/A | 36/259 | 13/241 | 2.96 (1.53, 5.72), 0.001 | N/A | |||||
| Vonaesch (2017) [40] Bangui, Central African Republic | 28/148 | 29/266 | 1.91 (1.09, 3.35), 0.025 | 0.370 * | |||||||||||||
| Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 178/303 | 66/319 | 5.46 (3.83, 7.78), <0.00001 | N/A | |||||||||||||
| Afridi (2020) [42] Skardu, Pakistan | 35/161 | 12/139 | 2.94 (1.46, 5.92), 0.003 | N/A | |||||||||||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 167/301 | 75/286 | 3.51 (2.48, 4.97), <0.00001 | N/A | 57/301 | 20/286 | 3.11 (1.81, 5.32), <0.0001 | N/A | 91/301 | 38/286 | 2.83 (1.86, 4.31), <0.00001 | N/A | |||||
| Silva (2009) [47] Itinga, Vale of Jequitinhonha, Brazil | 116/378 | 148/653 | 1.51 (1.14, 2.01), 0.0005 | N/A | |||||||||||||
| Suchdev (2014) [48] Kibera, Nairobi, Kenya | 27/82 | 32/123 | 1.40 (0.76, 2.57), 0.290 | N/A | |||||||||||||
| Zulkifli (1999) [49] Kelantan, Malaysia | 94/127 | 71/141 | 2.81 (1.68, 4.70), <0.00001 | N/A | 22/127 | 25/141 | 0.97 (0.52, 1.83), 0.93 | N/A | 91/127 | 71/141 | 2.49 (1.50, 4.14), 0.0004 | N/A | |||||
| Ascaris lumbricoides | Aiemjoy (2017) [36] Amhara, Ethiopia | 3/23 | 22/178 | 1.06 (0.29, 3.88), 0.93 | 1.44 (0.39, 5.41), 0.59 + | 3/23 | 17/178 | 1.42 (0.38, 5.28), 0.60 | 2.54 (1.02, 6.32), 0.045 + | 0/23 | 16/178 | 0.21 (0.01, 3.61), 0.28 | N/A | ||||
| Gutiérrez-Jiménez (2019) [37] Chiapas Highlands, Mexico | 38/48 | 29/36 | 0.92 (0.31, 2.70), 0.88 | N/A | |||||||||||||
| Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 62/67 | 182/555 | 25.41 (10.04, 64.30), 0.00001 | N/A | |||||||||||||
| Afridi (2020) [42] Skardu, Pakistan | 7/60 | 40/233 | 0.64 (0.27, 1.50), 0.30 | N/A | |||||||||||||
| Aswatshi (1997) [43] Lucknow, India | 74/124 | 577/916 | 0.87 (0.59, 1.28), 0.47 | N/A | 24/124 | 254/916 | 0.63 (0.39, 1.00) 0.05 | N/A | 77/124 | 626/916 | 0.76 (0.51, 1.12), 0.160 | N/A | |||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 135/252 | 107/345 | 2.46 (1.75, 3.45), <0.00001 | N/A | 47/252 | 30/335 | 2.33 (1.43, 3.81), 0.0007 | N/A | 76/252 | 54/345 | 2.25 (1.51, 3.34), <0.00001 | N/A | |||||
| Silva (2009) [47] Itinga, Vale of Jequitinhonha, Brazil | 46/115 | 218/916 | 2.13 (1.43, 3.19), 0.0002 | N/A | |||||||||||||
| Suchdev (2014) [48] Kibera, Nairobi, Kenya | 18/50 | 41/155 | 1.56 (0.79, 3.08), 0.20 | N/A | |||||||||||||
| Zulkifli (1999) [49] Kelantan, Malaysia | 34/48 | 161/220 | 0.89 (0.45, 1.77), 0.74 | N/A | |||||||||||||
| Cryptosporidium sp. | Afridi (2020) [42] Skardu, Pakistan | 9/42 | 38/258 | 1.58 (0.70, 3.56), 0.27 | N/A | ||||||||||||
| Entamoeba histolytica | Gutiérrez-Jiménez (2019) [47] Chiapas highland, Mexico | 26/32 | 41/52 | 1.16 (0.38, 3.53), 0.79 | N/A | ||||||||||||
| Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 21/29 | 223/593 | 4.36 (1.90, 10.00), 0.0005 | N/A | |||||||||||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 8/11 | 234/576 | 3.90 (1.02, 14.84), 0.05 | N/A | 1/11 | 76/576 | 0.66 (0.08, 5.21), 0.69 | N/A | 5/11 | 125/576 | 3.01 (0.90, 10.01), 0.07 | N/A | |||||
| Enterobius vermicularis | Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 2/6 | 240/581 | 0.71 (0.13, 3.91), 0.69 | N/A | 1/6 | 76/581 | 1.33 (0.15, 11.53), 0.80 | N/A | 1/6 | 129/581 | 0.70 (0.08, 6.05), 0.75 | N/A | ||||
| Haratipour (2016) [38] Shahroud, Iran | 69/230 | 86/1620 | 7.64 (5.36, 10.91), <0.00001 | N/A | |||||||||||||
| Giardia lamblia | Aiemjoy (2017) [36] Amhara, Ethiopia | 4/22 | 21/189 | 1.78 (0.55, 5.75), 0.34 | 2.13 (0.83, 5.43) 0.11+ | 5/22 | 15/189 | 3.41 (1.10, 10.54), 0.03 | 9.50 (3.61, 24.98), <0.0001 + | 2/22 | 14/189 | 1.25 (0.26, 5.90), 0.78 | 1.26 (0.34, 4.62), 0.76 + | ||||
| Vonaesch (2017) [40] Bangui, Central African Republic | 15/148 | 17/266 | 1.65 (0.80, 3.41), 0.175 | 0.801 * | |||||||||||||
| Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 56/65 | 188/557 | 12.21 (5.91, 25.23), <0.00001 | N/A | |||||||||||||
| Afridi (2020) [42] Skardu, Pakistan | 8/24 | 39/276 | 3.04 (1.22, 7.58), 0.02 | N/A | |||||||||||||
| Aswatshi (1997) [43] Lucknow, India | 34/60 | 617/980 | 0.77 (0.45, 1.30), 0.33 | N/A | 20/60 | 258/980 | 1.40 (0.80, 2.44), 0.240 | N/A | 40/60 | 663/980 | 0.96 (0.55, 1.66), 0.870 | N/A | |||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 18/26 | 224/561 | 3.39 (1.45, 7.92), 0.005 | N/A | 3/26 | 74/561 | 0.86 (0.25, 2.93), 0.81 | N/A | 11/26 | 119/561 | 2.72 (1.22, 6.09), 0.010 | N/A | |||||
| Gutiérrez-Jiménez (2019) [38] Chiapas Highlands, Mexico | 12/14 | 55/70 | 1.64 (0.33, 8.12), 0.55 | N/A | |||||||||||||
| Sajjadi (2005) [46] Marvdhast, Iran | 9/71 | 18/229 | 1.70 (0.73, 3.98), 0.22 | N/A | 3/71 | 0/229 | 23.45 (1.20, 459.63), 0.04 | N/A | 8/71 | 10/219 | 2.78 (1.05, 7.34), 0.04 | N/A | |||||
| Ignatius (2012) [44] Butare and Huye, Rwanda | 84/325 | 33/165 | 1.39 (0.88, 2.20), 0.15 | N/A | |||||||||||||
| Hookworm | Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 44/49 | 200/573 | 16.41 (6.41, 42.05) <0.00001 | N/A | ||||||||||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 7/17 | 301/570 | 0.63 (0.23, 1.67), 0.350 | N/A | 5/17 | 72/570 | 2.88 (0.99, 8.42), 0.045 | N/A | 7/17 | 301/570 | 0.63 (0.23, 1.67), 0.350 | N/A | |||||
| Silva (2009) [47] Itinga, Vale of Jequitinhonha, Brazil | 24/121 | 240/910 | 0.69 (0.43, 1.11), 0.12 | N/A | |||||||||||||
| Zulkifli (1999) [49] Kelantan, Malaysia | 8/10 | 187/258 | 1.52 (0.31, 7.32), 0.60 | ||||||||||||||
| Hymenolepsis nana | Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 4/6 | 238/581 | 2.88 (0.52, 15.86), 0.220 | N/A | 1/6 | 128/581 | 0.71 (0.08, 6.11), 0.750 | N/A | 2/6 | 128/581 | 1.77 (0.32, 9.77), 0.510 | N/A | ||||
| Afridi (2020) [42] Skardu, Pakistan | 11/28 | 36/272 | 4.24 (1.84, 9.78), 0.0007 | N/A | |||||||||||||
| Strongyloides sterocalis | Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 5/10 | 239/612 | 1.56 (0.45, 5.45), 0.490 | N/A | ||||||||||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 1/1 | 241/586 | 4.29 (0.17, 105.80), 0.37 | N/A | 0/1 | 77/586 | 2.19 (0.09, 54.27), 0.63 | N/A | 0/1 | 128/586 | 1.19 (0.05, 29.37), 0.92 | N/A | |||||
| Taenia spp. | Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 4/8 | 240/614 | 1.56 (0.39, 6.29), 0.530 | N/A | ||||||||||||
| Trichuris trichiura | Yoseph (2020) [41] Boricha Woreda, Southern Ethiopia | 27/38 | 217/484 | 3.02 (1.46, 6.23) 0.003 | N/A | ||||||||||||
| Kabeta (2017) [45] Hawasa Zuria, South Ethiopia | 3/11 | 239/576 | 0.53 (0.14, 2.01), 0.350 | N/A | 4/11 | 73/576 | 3.94 (1.12, 13.78), 0.030 | N/A | 2/11 | 128/576 | 0.78 (0.17, 3.65), 0.75 | N/A | |||||
| Silva (2009) [47] Itinga, Vale of Jequitinhonha, Brazil | 23/51 | 148/653 | 2.52 (1.42, 4.46), 0.002 | N/A | |||||||||||||
| Suchdev (2014) [48] Kibera, Nairobi, Kenya | 13/47 | 46/158 | 0.93 (0.45, 1.92), 0.85 | ||||||||||||||
| Zulkifli (1999) [49] Kelantan, Malaysia | 25/37 | 170/231 | 0.75 (0.35, 1.58), 0.45 | N/A | |||||||||||||
| Parasites | Country | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Africa | Asia | America | ||||||||||
| Ethiopia (Tropical) | Kenya (Tropical) | Central African Republic (Tropical) | Rwanda (Tropical) | Egypt (Sub-Tropical) | Pakistan (Temperate) | Iran (Temperate) | Turkey (Temperate) | India (Sub-Tropical) | Malaysia (Tropical) | Brazil (Sub-Tropical) | Mexico (Tropical) | |
| Not specified | 303/622 (48.71%) [41] 301/587 (51.28%) [45] | 82/205 (40.0%) [48] | 148/414 (35.77%) [40] | 259/500 (51.78%) [39] | 161/300 (53.67) [42] | 649/1850 (35.08%) [38] | 58/108 (53.71%) [50] | 127/268 (47.39%) [49] | 378/1031 (36.67%) [47] | |||
| A. lumbricoides | 23/201 (11.44%) [36] 67/622 (10.77%) [41] 252/597 (42.41%) [45] | 50/205 (24.39%) [48] | 60/293 (20.48%) [42] | 124/1040 (11.92%) [43] | 48/268 (17.91%) [49] | 115/1031 (11.15%) [47] | 48/84 (57.14%) [37] | |||||
| Cryptosporidium | 42/300 (14.00%) [42] | |||||||||||
| E. histolytica | 29/622 (4.66%) [41] 11/587 (1.87%) [45] | 32/84 (38.10%) [37] | ||||||||||
| E. vermicularis | 6/587 (1.02%) [45] | 230/1850 (12.42%) [38] | ||||||||||
| G. lamblia | 22/210 (10.48%) [36] 65/622 (12.06%) [41] 26/587 (4.43%) [45] | 148/414 (35.75%) [40] | 325/490 (66.33%) [44] | 24/300 (8.00%) [42] | 71/300 (23.67%) [46] | 60/1040 (5.77%) [43] | 14/84 (16.67%) [37] | |||||
| Hookworm | 49/622 (7.88%) [41] 17/587 (2.89%) [45] | 10/268 (3.73%) [49] | 121/1031 (11.73%) [47] | |||||||||
| H. nana | 6/587 (1.02%) [45] | 11/300 (3.67%) [42] | ||||||||||
| S. sterocalis | 10/622 (1.61%) [41] 1/587 (1.70%) [45] | |||||||||||
| Taenia spp. | 8/622 (1.29%) [41] | |||||||||||
| T. trichiura | 38/622 (6.11%) [41] 11/587 (1.87%) [45] | 50/205 (24.39%) [48] | 37/268 (13.81%) [49] | 51/1031 (4.97%) [47] | ||||||||
| Studies | Risk Factors | Summary of Findings |
|---|---|---|
| Afridi (2020) [42] | Socioeconomic background | Most children with parasitic infections belonged to low or low-middle SE groups, with 165/300 (55%) children from low SE, 90/300 (30%) from low-middle SE, 30/300 (10%) from middle SE and 15/300 (5%) from upper middle SE groups, respectively. |
| Aiemjoy (2017) [36] | Household income (socioeconomic background) | Higher prevalence of Ascaris lumbricoides infections was found in children from families with a household income of <1USD/day with a 6.68 (95% CI 1.01, 44.34, p = 0.042) prevalence ratio. |
| Gutierrez-Jimenez (2019) [38] | Rural vs urban municipalities (socioeconomic background) | Intestinal parasitic infections were only found in children that lived in rural areas. No infections were found in urban children. There were contrasting conditions of rural houses compared to urban houses, where the former had earthen floor and no access to potable water. |
| Silva (2009) [47] | Family members (socioeconomic background) | Children that lived with more than 5 family members were more likely to be infected with G. lamblia (OR 1.8 (95% CI 1.2–2.8), p = 0.011). |
| Number of bedrooms (socioeconomic background) | Children that lived in a house with less than 2 bedrooms were more likely to be infected with G. lamblia (OR 1.5 CI (95% 1.1–3.0), p = 0.012). | |
| Have brothers/sisters <5 years old | G. lamblia infection was noted to be higher in those with brothers/sisters <5 years old (OR 2.7 (95% CI 1.5–4.5), p = 0.000). | |
| Tap water (sanitation) | Children without access to tap water experienced a 2.1-fold (1.3–3.3, p = 0.001) increased risk of acquiring Gl.lamblia infection. | |
| Sewage system (sanitation) | G. lamblia infection was more prevalent in children that lived in a house without a sewage system with an odds ratio of 1.8 (1.1–3.1, p = 0.030). | |
| Doni (2015) [50] | Maternal education (socioeconomic background) | Higher risk of infection in children with illiterate mother (OR 3.019 (95% CI 1.279–7.130, p = 0.012)). |
| Paternal education (socioeconomic background) | Higher risk of infection in children with illiterate father (OR 4.954 (95% CI 1.819–13.492, p = 0.012)). | |
| Poor economic situation (socioeconomic background) | Higher odds of infection found in children born into a financially poor family (OR 2.945 (95% CI 1.292–6.717, p = 0.010). | |
| Number of households (6 or above) | Higher risk of infections was found in children living with more than 6 family members (OR 2.865 (95% CI 1.233–6.667), p = 0.014). | |
| Children aged 36 months and below | Increased risk of infection in children younger than 36 months (OR 1.93 (95% CI 1.14–3.29), p = 0.036). | |
| Playing with soil (hygiene) | Higher risk of infections in children playing with soil (OR 4.956 (95% CI 1.856–13.528), p = 0.000). | |
| Not washing hands after toilet (hygiene) | Higher risk of infections in children not washing hands after the toilet (OR 6.369 (95%CI 1.361–2.941), p = 0.019). | |
| Wrong technique of washing hands (hygiene) | Higher infections in children not washing hands after the toilet (OR 6.369 (95%CI 1.361–7.299), p = 0.007). | |
| Ignatius (2017) [44] | Age of more than 1 year | Increased risk of G. lamblia infections in children aged >1 year with the following odds ratios: 1–2 years: 1.97 (1.08–3.59, p = 0.03) 2–3 years: 3.82 (2.06–7.07, p <0.0001) 3–4 years: 4.52 (2.65–7.72, p <0.0001) 4–5 years: 5.52 (3.15–9.70, p < 0.0001). |
| Breastfed | Lower risk of infections in breastfed children (OR 0.40 (95% CI 0.32–0.51), p <0.0004). | |
| Number of siblings | Higher risk of infections in children that had more than 4 siblings (OR 3.14 (95% CI 1.87–5.27), p = 0.0001). | |
| Yoseph (2020) [41] | Consuming raw vegetables and fruits | Higher risk of infections in children that consumed uncooked vegetables and fruits (OR 2.65 (95% CI 1.6–4.7). |
| Absence of sanitation facility | Higher risk of infections in children living without sanitation facilities (OR 2.9 (95% CI 1.6–5.3)). | |
| Wearing shoes | Higher risk of infections in children not wearing shoes (OR 3.5 (95% CI 2.–5.7). | |
| Family size | Higher risk of infections in children living with a large family (OR 2.7 (95% CI 1.5–5.0). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fauziah, N.; Aviani, J.K.; Agrianfanny, Y.N.; Fatimah, S.N. Intestinal Parasitic Infection and Nutritional Status in Children under Five Years Old: A Systematic Review. Trop. Med. Infect. Dis. 2022, 7, 371. https://doi.org/10.3390/tropicalmed7110371
Fauziah N, Aviani JK, Agrianfanny YN, Fatimah SN. Intestinal Parasitic Infection and Nutritional Status in Children under Five Years Old: A Systematic Review. Tropical Medicine and Infectious Disease. 2022; 7(11):371. https://doi.org/10.3390/tropicalmed7110371
Chicago/Turabian StyleFauziah, Nisa, Jenifer Kiem Aviani, Yukan Niko Agrianfanny, and Siti Nur Fatimah. 2022. "Intestinal Parasitic Infection and Nutritional Status in Children under Five Years Old: A Systematic Review" Tropical Medicine and Infectious Disease 7, no. 11: 371. https://doi.org/10.3390/tropicalmed7110371
APA StyleFauziah, N., Aviani, J. K., Agrianfanny, Y. N., & Fatimah, S. N. (2022). Intestinal Parasitic Infection and Nutritional Status in Children under Five Years Old: A Systematic Review. Tropical Medicine and Infectious Disease, 7(11), 371. https://doi.org/10.3390/tropicalmed7110371