
Pathogenesis and Manifestations of Zika Virus-Associated Ocular Diseases

Abstract
:1. Introduction
2. Virology
3. Host–Virus Interaction
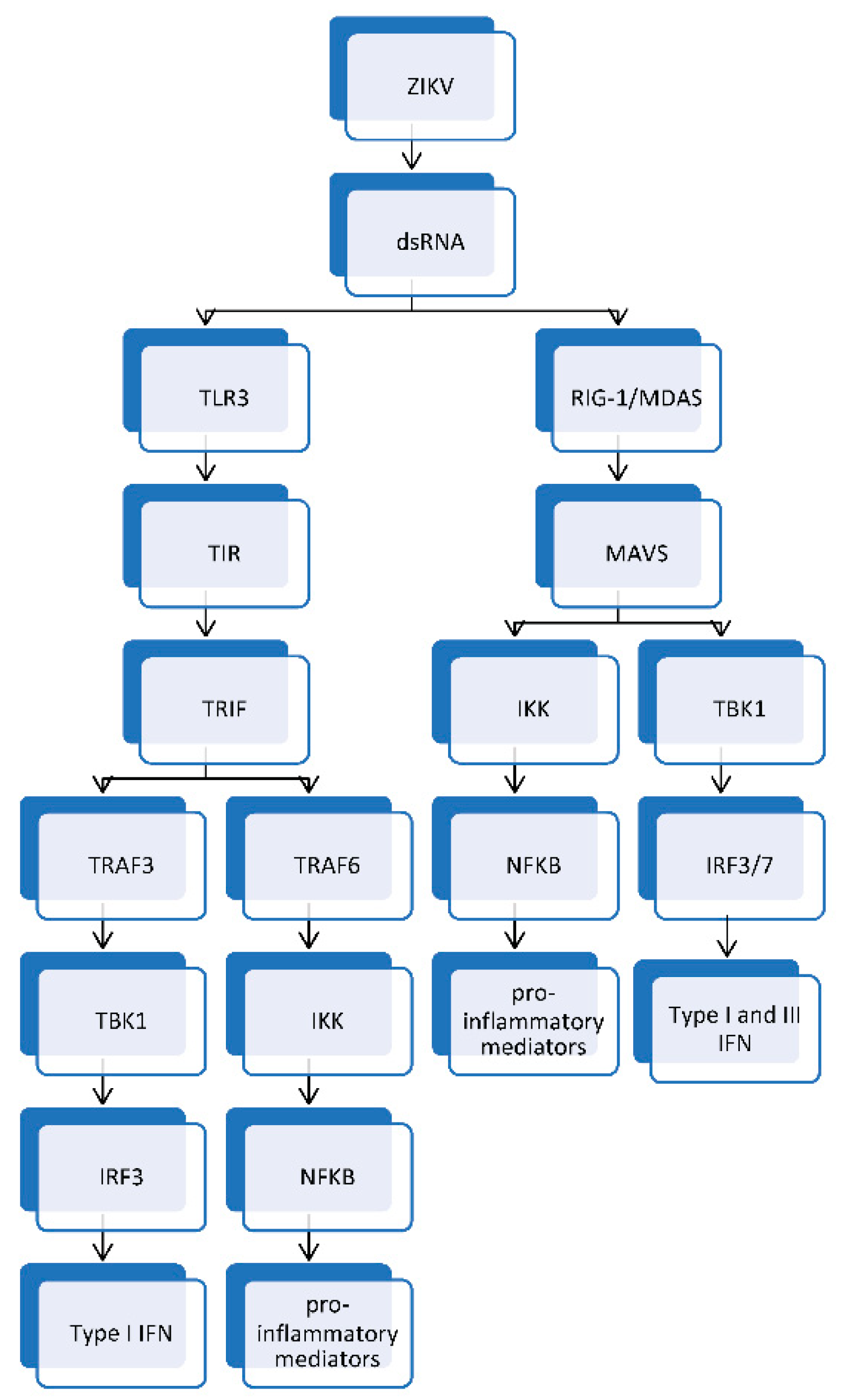
3.1. Innate Immune Response to ZIKV Infection
3.2. T Cell-Mediated Immune Response to ZIKV Infection
3.3. Antibody-Mediated Immune Response to ZIKV Infection
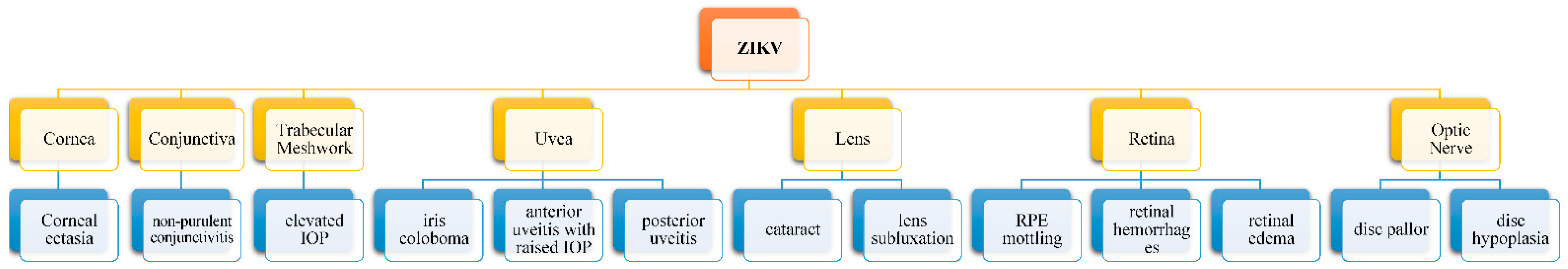
4. Ocular Manifestations
5. Pathological Mechanisms
5.1. Blood–Tissue Barriers
5.2. ZIKV-Infected Myeloid Cells
5.3. Microcephaly
5.4. Placental Dysfunction
5.5. Congenital Zika Syndrome
5.6. Acute ZIKV Infection
5.6.1. Retina
5.6.2. Cornea
5.6.3. Trabecular Meshwork
5.6.4. Conjunctiva
5.6.5. Anterior Uvea
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Song, B.H.; Yun, S.I.; Woolley, M.; Lee, Y.M. Zika virus: History, epidemiology, transmission, and clinical presentation. J. Neuroimmunol. 2017, 308, 50–64. [Google Scholar] [CrossRef] [Green Version]
- Dick, G.W. Zika virus. II. Pathogenicity and physical properties. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 521–534. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Culshaw, A.; Mongkolsapaya, J.; Screaton, G. The immunology of Zika Virus. F1000Research 2018, 7, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquezan, M.C.; Ventura, C.V.; Sheffield, J.S.; Golden, W.C.; Omiadze, R.; Belfort, R., Jr.; May, W. Ocular effects of Zika virus-a review. Surv. Ophthalmol. 2018, 63, 166–173. [Google Scholar] [CrossRef]
- Merle, H.; Donnio, A.; Jean-Charles, A.; Guyomarch, J.; Hage, R.; Najioullah, F.; Cesaire, R.; Cabie, A. Ocular manifestations of emerging arboviruses: Dengue fever, Chikungunya, Zika virus, West Nile virus, and yellow fever. J. Français Ophtalmol. 2018, 41, e235–e243. [Google Scholar] [CrossRef]
- Gregory, C.J.; Oduyebo, T.; Brault, A.C.; Brooks, J.T.; Chung, K.W.; Hills, S.; Kuehnert, M.J.; Mead, P.; Meaney-Delman, D.; Rabe, I.; et al. Modes of Transmission of Zika Virus. J. Infect. Dis. 2017, 216, S875–S883. [Google Scholar] [CrossRef] [Green Version]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Younger, D.S. Epidemiology of Zika Virus. Neurol. Clin. 2016, 34, 1049–1056. [Google Scholar] [CrossRef]
- Beaver, J.T.; Lelutiu, N.; Habib, R.; Skountzou, I. Evolution of Two Major Zika Virus Lineages: Implications for Pathology, Immune Response, and Vaccine Development. Front. Immunol. 2018, 9, 1640. [Google Scholar] [CrossRef]
- Tan, J.J.L.; Balne, P.K.; Leo, Y.S.; Tong, L.; Ng, L.F.P.; Agrawal, R. Persistence of Zika virus in conjunctival fluid of convalescence patients. Sci. Rep. 2017, 7, 11194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, S.; Schlaberg, R.; Lewis, J.; Hanson, K.E.; Couturier, M.R. Fatal Zika Virus Infection with Secondary Nonsexual Transmission. N. Engl. J. Med. 2016, 375, 1907–1909. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Vieira, D.F.; Jacome, F.C.; da Silva, M.A.N.; Caldas, G.C.; de Filippis, A.M.B.; de Sequeira, P.C.; de Souza, E.M.; Andrade, A.A.; Manso, P.P.A.; Trindade, G.F.; et al. Structural investigation of C6/36 and Vero cell cultures infected with a Brazilian Zika virus. PLoS ONE 2017, 12, e0184397. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Fernandez, E.; Dowd, K.A.; Speer, S.D.; Platt, D.J.; Gorman, M.J.; Govero, J.; Nelson, C.A.; Pierson, T.C.; Diamond, M.S.; et al. Structural Basis of Zika Virus-Specific Antibody Protection. Cell 2016, 166, 1016–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, A.; Patil, A.A.; Skariyachan, S. Recent Perspectives on Genome, Transmission, Clinical Manifestation, Diagnosis, Therapeutic Strategies, Vaccine Developments, and Challenges of Zika Virus Research. Front. Microbiol. 2017, 8, 1761. [Google Scholar] [CrossRef] [PubMed]
- Atif, M.; Azeem, M.; Sarwar, M.R.; Bashir, A. Zika virus disease: A current review of the literature. Infection 2016, 44, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Sironi, M.; Forni, D.; Clerici, M.; Cagliani, R. Nonstructural Proteins Are Preferential Positive Selection Targets in Zika Virus and Related Flaviviruses. PLoS Negl. Trop. Dis. 2016, 10, e0004978. [Google Scholar] [CrossRef] [Green Version]
- Kostyuchenko, V.A.; Lim, E.X.; Zhang, S.; Fibriansah, G.; Ng, T.S.; Ooi, J.S.; Shi, J.; Lok, S.M. Structure of the thermally stable Zika virus. Nature 2016, 533, 425–428. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Seong, R.K.; Son, S.W.; Shin, O.S. Insights into ZIKV-Mediated Innate Immune Responses in Human Dermal Fibroblasts and Epidermal Keratinocytes. J. Investig. Dermatol. 2019, 139, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Bos, S.; Li, G.; Wang, S.; Gadea, G.; Despres, P.; Zhao, R.Y. Probing Molecular Insights into Zika Virus–Host Interactions. Viruses 2018, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Robbiani, D.F.; Bozzacco, L.; Keeffe, J.R.; Khouri, R.; Olsen, P.C.; Gazumyan, A.; Schaefer-Babajew, D.; Avila-Rios, S.; Nogueira, L.; Patel, R.; et al. Recurrent Potent Human Neutralizing Antibodies to Zika Virus in Brazil and Mexico. Cell 2017, 169, 597–609.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, A.P.; Moraes, A.H. Zika virus proteins at an atomic scale: How does structural biology help us to understand and develop vaccines and drugs against Zika virus infection? J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e20190013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Vasudevan, S.G.; Lescar, J. The flavivirus NS2B-NS3 protease-helicase as a target for antiviral drug development. Antivir. Res. 2015, 118, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Shin, O.S. Advances in Zika Virus(-)Host Cell Interaction: Current Knowledge and Future Perspectives. Int. J. Mol. Sci. 2019, 20, 1101. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.H.M.; Moises, R.N.C.; Alves, B.E.B.; Pereira, H.W.B.; de Paiva, A.A.P.; Morais, I.C.; Nascimento, Y.M.; Monteiro, J.D.; de Souto, J.T.; Nascimento, M.S.L.; et al. Innate immune response in patients with acute Zika virus infection. Med. Microbiol. Immunol. 2019, 208, 703–714. [Google Scholar] [CrossRef]
- Wen, Z.; Song, H.; Ming, G.L. How does Zika virus cause microcephaly? Genes Dev. 2017, 31, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Routhu, N.K.; Byrareddy, S.N. Host-Virus Interaction of ZIKA Virus in Modulating Disease Pathogenesis. J. Neuroimmune Pharmacol. 2017, 12, 219–232. [Google Scholar] [CrossRef]
- Sirohi, D.; Kuhn, R.J. Zika Virus Structure, Maturation, and Receptors. J. Infect. Dis. 2017, 216, S935–S944. [Google Scholar] [CrossRef] [Green Version]
- Chigbu, D.I.; Jain, P.; Crumley, B.L.; Patel, D.; Khan, Z.K. Human T cell leukemia virus type 1 and Zika virus: Tale of two reemerging viruses with neuropathological sequelae of public health concern. J. Neurovirol. 2019, 25, 289–300. [Google Scholar] [CrossRef]
- Beys-da-Silva, W.O.; Rosa, R.L.; Santi, L.; Berger, M.; Park, S.K.; Campos, A.R.; Terraciano, P.; Varela, A.P.M.; Teixeira, T.F.; Roehe, P.M.; et al. Zika Virus Infection of Human Mesenchymal Stem Cells Promotes Differential Expression of Proteins Linked to Several Neurological Diseases. Mol. Neurobiol. 2019, 56, 4708–4717. [Google Scholar] [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, A.B.; Talarico, L.B. Role of the complement system in antibody-dependent enhancement of flavivirus infections. Int. J. Infect. Dis. 2021, 103, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Merle, N.S.; Noe, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part II: Role in Immunity. Front. Immunol. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, K.M.; Liszewski, M.K.; Nybakken, G.; Davis, A.E.; Townsend, R.R.; Fremont, D.H.; Atkinson, J.P.; Diamond, M.S. West Nile virus nonstructural protein NS1 inhibits complement activation by binding the regulatory protein factor H. Proc. Natl. Acad. Sci. USA 2006, 103, 19111–19116. [Google Scholar] [CrossRef] [Green Version]
- Malekshahi, Z.; Schiela, B.; Bernklau, S.; Banki, Z.; Wurzner, R.; Stoiber, H. Interference of the Zika Virus E-Protein With the Membrane Attack Complex of the Complement System. Front. Immunol. 2020, 11, 569549. [Google Scholar] [CrossRef]
- Ma, J.; Ketkar, H.; Geng, T.; Lo, E.; Wang, L.; Xi, J.; Sun, Q.; Zhu, Z.; Cui, Y.; Yang, L.; et al. Zika Virus Non-structural Protein 4A Blocks the RLR-MAVS Signaling. Front. Microbiol. 2018, 9, 1350. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Cai, X.; Wu, J.; Cong, Q.; Chen, X.; Li, T.; Du, F.; Ren, J.; Wu, Y.T.; Grishin, N.V.; et al. Phosphorylation of innate immune adaptor proteins MAVS, STING, and TRIF induces IRF3 activation. Science 2015, 347, aaa2630. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liu, S.; Chen, Z.J. SnapShot: Pathways of antiviral innate immunity. Cell 2010, 140, 436.e2. [Google Scholar] [CrossRef] [Green Version]
- Seth, R.B.; Sun, L.; Chen, Z.J. Antiviral innate immunity pathways. Cell Res. 2006, 16, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Vercammen, E.; Staal, J.; Beyaert, R. Sensing of viral infection and activation of innate immunity by toll-like receptor 3. Clin. Microbiol. Rev. 2008, 21, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Hacker, H.; Redecke, V.; Blagoev, B.; Kratchmarova, I.; Hsu, L.C.; Wang, G.G.; Kamps, M.P.; Raz, E.; Wagner, H.; Hacker, G.; et al. Specificity in Toll-like receptor signalling through distinct effector functions of TRAF3 and TRAF6. Nature 2006, 439, 204–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorkstrom, N.K.; Strunz, B.; Ljunggren, H.G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, L.; Valache, M.; Facchini, F.A.; Granucci, F. How dendritic cells sense and respond to viral infections. Clin. Sci. 2021, 135, 2217–2242. [Google Scholar] [CrossRef]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4(+) T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef]
- Hassert, M.; Wolf, K.J.; Schwetye, K.E.; DiPaolo, R.J.; Brien, J.D.; Pinto, A.K. CD4+T cells mediate protection against Zika associated severe disease in a mouse model of infection. PLoS Pathog. 2018, 14, e1007237. [Google Scholar] [CrossRef]
- Tonnerre, P.; Melgaco, J.G.; Torres-Cornejo, A.; Pinto, M.A.; Yue, C.; Blumel, J.; de Sousa, P.S.F.; de Mello, V.D.M.; Moran, J.; de Filippis, A.M.B.; et al. Evolution of the innate and adaptive immune response in women with acute Zika virus infection. Nat. Microbiol. 2020, 5, 76–83. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Craft, J.E.; Kaech, S.M. The multifaceted role of CD4(+) T cells in CD8(+) T cell memory. Nat. Rev. Immunol. 2016, 16, 102–111. [Google Scholar] [CrossRef]
- Elong Ngono, A.; Vizcarra, E.A.; Tang, W.W.; Sheets, N.; Joo, Y.; Kim, K.; Gorman, M.J.; Diamond, M.S.; Shresta, S. Mapping and Role of the CD8(+) T Cell Response During Primary Zika Virus Infection in Mice. Cell Host Microbe 2017, 21, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Priyamvada, L.; Suthar, M.S.; Ahmed, R.; Wrammert, J. Humoral Immune Responses Against Zika Virus Infection and the Importance of Preexisting Flavivirus Immunity. J. Infect. Dis. 2017, 216, S906–S911. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Rouphael, N.; Xu, Y.; Natrajan, M.S.; Beck, A.; Hart, M.; Feldhammer, M.; Feldpausch, A.; Hill, C.; Wu, H.; et al. Innate, T-, and B-Cell Responses in Acute Human Zika Patients. Clin. Infect. Dis. 2018, 66, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Screaton, G.; Mongkolsapaya, J.; Yacoub, S.; Roberts, C. New insights into the immunopathology and control of dengue virus infection. Nat. Rev. Immunol. 2015, 15, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Miranda, G.; Lim, S.M.; Mohsen, M.O.; Pobelov, I.V.; Roesti, E.S.; Heath, M.D.; Skinner, M.A.; Kramer, M.F.; Martina, B.E.E.; Bachmann, M.F. Zika Virus-Derived E-DIII Protein Displayed on Immunologically Optimized VLPs Induces Neutralizing Antibodies without Causing Enhancement of Dengue Virus Infection. Vaccines 2019, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Stettler, K.; Beltramello, M.; Espinosa, D.A.; Graham, V.; Cassotta, A.; Bianchi, S.; Vanzetta, F.; Minola, A.; Jaconi, S.; Mele, F.; et al. Specificity, cross-reactivity, and function of antibodies elicited by Zika virus infection. Science 2016, 353, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, R.; Oo, H.H.; Balne, P.K.; Ng, L.; Tong, L.; Leo, Y.S. Zika Virus and the Eye. Ocul. Immunol. Inflamm. 2018, 26, 654–659. [Google Scholar] [CrossRef]
- Furtado, J.M.; Esposito, D.L.; Klein, T.M.; Teixeira-Pinto, T.; da Fonseca, B.A. Uveitis Associated with Zika Virus Infection. N. Engl. J. Med. 2016, 375, 394–396. [Google Scholar] [CrossRef]
- Troumani, Y.; Touhami, S.; Jackson, T.L.; Ventura, C.V.; Stanescu-Segall, D.M.; Errera, M.H.; Rousset, D.; Bodaghi, B.; Cartry, G.; David, T.; et al. Association of Anterior Uveitis With Acute Zika Virus Infection in Adults. JAMA Ophthalmol. 2021, 139, 95–102. [Google Scholar] [CrossRef]
- Kodati, S.; Palmore, T.N.; Spellman, F.A.; Cunningham, D.; Weistrop, B.; Sen, H.N. Bilateral posterior uveitis associated with Zika virus infection. Lancet 2017, 389, 125–126. [Google Scholar] [CrossRef] [Green Version]
- Parke, D.W., 3rd; Almeida, D.R.; Albini, T.A.; Ventura, C.V.; Berrocal, A.M.; Mittra, R.A. Serologically Confirmed Zika-Related Unilateral Acute Maculopathy in an Adult. Ophthalmology 2016, 123, 2432–2433. [Google Scholar] [CrossRef]
- De Paula Freitas, B.; Ventura, C.V.; Maia, M.; Belfort, R., Jr. Zika virus and the eye. Curr. Opin. Ophthalmol. 2017, 28, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Buathong, R.; Hermann, L.; Thaisomboonsuk, B.; Rutvisuttinunt, W.; Klungthong, C.; Chinnawirotpisan, P.; Manasatienkij, W.; Nisalak, A.; Fernandez, S.; Yoon, I.K.; et al. Detection of Zika Virus Infection in Thailand, 2012–2014. Am. J. Trop. Med. Hyg. 2015, 93, 380–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paula Freitas, B.; de Oliveira Dias, J.R.; Prazeres, J.; Sacramento, G.A.; Ko, A.I.; Maia, M.; Belfort, R., Jr. Ocular Findings in Infants With Microcephaly Associated With Presumed Zika Virus Congenital Infection in Salvador, Brazil. JAMA Ophthalmol. 2016, 134, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, B.M. Zika virus-related hypertensive iridocyclitis. Arq. Bras. Oftalmol. 2016, 79, 63. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, P.; Kestelman, E.; Kestelman, B.; Vizzoni, A.G.; Cerbino-Neto, J.; Curi, A.L.L. Multifocal Choroiditis Secondary to Acute Zika Virus Infection. Ocul. Immunol. Inflamm. 2020, 28, 952–955. [Google Scholar] [CrossRef]
- Man, B.L. Total internal and external ophthalmoplegia as presenting symptoms of Miller Fisher syndrome. BMJ Case Rep. 2014, 2014, bcr2014205554. [Google Scholar] [CrossRef] [Green Version]
- Karam, E.; Giraldo, J.; Rodriguez, F.; Hernandez-Pereira, C.E.; Rodriguez-Morales, A.J.; Blohm, G.M.; Paniz-Mondolfi, A.E. Ocular flutter following Zika virus infection. J. Neurovirol. 2017, 23, 932–934. [Google Scholar] [CrossRef]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Parra, B.; Lizarazo, J.; Jimenez-Arango, J.A.; Zea-Vera, A.F.; Gonzalez-Manrique, G.; Vargas, J.; Angarita, J.A.; Zuniga, G.; Lopez-Gonzalez, R.; Beltran, C.L.; et al. Guillain-Barre Syndrome Associated with Zika Virus Infection in Colombia. N. Engl. J. Med. 2016, 375, 1513–1523. [Google Scholar] [CrossRef]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Castillo-Medina, L.F.; Rodriguez, Y.; Pacheco, Y.; Halstead, S.; Willison, H.J.; Anaya, J.M.; Ramirez-Santana, C. Autoimmune Neurological Conditions Associated With Zika Virus Infection. Front. Mol. Neurosci. 2018, 11, 116. [Google Scholar] [CrossRef]
- White, M.K.; Wollebo, H.S.; David Beckham, J.; Tyler, K.L.; Khalili, K. Zika virus: An emergent neuropathological agent. Ann. Neurol. 2016, 80, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Lemos, J.; Eggenberger, E. Saccadic intrusions: Review and update. Curr. Opin. Neurol. 2013, 26, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Wiest, G.; Safoschnik, G.; Schnaberth, G.; Mueller, C. Ocular flutter and truncal ataxia may be associated with enterovirus infection. J. Neurol. 1997, 244, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Do Rosario, M.S.; Giovanetti, M.; de Jesus, P.A.P.; Farias, D.S.; Faria, N.R.; de Lima, C.P.S.; da Silva, S.P.; Nunes, M.R.; Alcantara, L.C.J.; de Siqueira, I.C. Opsoclonus-myoclonus-ataxia syndrome associated with chikungunya and dengue virus co-infection. Int. J. Infect. Dis. 2018, 75, 11–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, C.V.; Maia, M.; Travassos, S.B.; Martins, T.T.; Patriota, F.; Nunes, M.E.; Agra, C.; Torres, V.L.; van der Linden, V.; Ramos, R.C.; et al. Risk Factors Associated With the Ophthalmoscopic Findings Identified in Infants With Presumed Zika Virus Congenital Infection. JAMA Ophthalmol. 2016, 134, 912–918. [Google Scholar] [CrossRef]
- Yasri, S.; Wiwanitkit, V. Glaucoma in Congenital Zika Syndrome. J. Glaucoma 2019, 28, e131. [Google Scholar] [CrossRef] [PubMed]
- Leyser, M.; Nascimento, O.J.M. Congenital Zika Virus Infection: Beyond Neonatal Microcephaly. JAMA Neurol. 2017, 74, 610. [Google Scholar] [CrossRef] [PubMed]
- Cofre, F. Zika virus intrauterine infection causes fetal brain abnormality and microcephaly: Tip of the iceberg? Rev. Chilena. Infectol. 2016, 33, 96. [Google Scholar] [CrossRef] [Green Version]
- Ventura, C.V.; Maia, M.; Bravo-Filho, V.; Gois, A.L.; Belfort, R., Jr. Zika virus in Brazil and macular atrophy in a child with microcephaly. Lancet 2016, 387, 228. [Google Scholar] [CrossRef] [Green Version]
- Yepez, J.B.; Murati, F.A.; Pettito, M.; Penaranda, C.F.; de Yepez, J.; Maestre, G.; Arevalo, J.F.; Johns Hopkins Zika, C. Ophthalmic Manifestations of Congenital Zika Syndrome in Colombia and Venezuela. JAMA Ophthalmol. 2017, 135, 440–445. [Google Scholar] [CrossRef]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.; Horovitz, D.D.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.; Neri, J.I.; Neto, J.M.; Wanderley, H.Y.; et al. Possible Association Between Zika Virus Infection and Microcephaly—Brazil, 2015. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Sahiner, F.; Sig, A.K.; Savasci, U.; Tekin, K.; Akay, F. Zika Virus-associated Ocular and Neurologic Disorders: The Emergence of New Evidence. Pediatr. Infect. Dis. J. 2017, 36, e341–e346. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, G.C.; Macedo Pereira, C.M.; Toledo de Paula, C.H.; de Souza Haueisen Barbosa, P.; Machado de Souza, D.; Coelho, L.M. Corneal ectasia and high ametropia in an infant with microcephaly associated with presumed Zika virus congenital infection: New ocular findings. J. AAPOS 2019, 23, 354–356. [Google Scholar] [CrossRef] [PubMed]
- De Paula Freitas, B.; Zin, A.; Ko, A.; Maia, M.; Ventura, C.V.; Belfort, R., Jr. Anterior-Segment Ocular Findings and Microphthalmia in Congenital Zika Syndrome. Ophthalmology 2017, 124, 1876–1878. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.V.; Maia, M.; Ventura, B.V.; Linden, V.V.; Araujo, E.B.; Ramos, R.C.; Rocha, M.A.; Carvalho, M.D.; Belfort, R., Jr.; Ventura, L.O. Ophthalmological findings in infants with microcephaly and presumable intra-uterus Zika virus infection. Arq. Bras. Oftalmol. 2016, 79, 1–3. [Google Scholar] [CrossRef]
- Zin, A.A.; Tsui, I.; Rossetto, J.; Vasconcelos, Z.; Adachi, K.; Valderramos, S.; Halai, U.A.; Pone, M.; Pone, S.M.; Silveira Filho, J.C.B.; et al. Screening Criteria for Ophthalmic Manifestations of Congenital Zika Virus Infection. JAMA Pediatr. 2017, 171, 847–854. [Google Scholar] [CrossRef]
- Ventura, C.V.; Ventura, L.O.; Bravo-Filho, V.; Martins, T.T.; Berrocal, A.M.; Gois, A.L.; de Oliveira Dias, J.R.; Araujo, L.; Escariao, P.; van der Linden, V.; et al. Optical Coherence Tomography of Retinal Lesions in Infants With Congenital Zika Syndrome. JAMA Ophthalmol. 2016, 134, 1420–1427. [Google Scholar] [CrossRef]
- De Oliveira Dias, J.R.; Ventura, C.V.; de Paula Freitas, B.; Prazeres, J.; Ventura, L.O.; Bravo-Filho, V.; Aleman, T.; Ko, A.I.; Zin, A.; Belfort, R., Jr.; et al. Zika and the Eye: Pieces of a Puzzle. Prog. Retin. Eye Res. 2018, 66, 85–106. [Google Scholar] [CrossRef]
- Miranda, H.A., 2nd; Costa, M.C.; Frazao, M.A.M.; Simao, N.; Franchischini, S.; Moshfeghi, D.M. Expanded Spectrum of Congenital Ocular Findings in Microcephaly with Presumed Zika Infection. Ophthalmology 2016, 123, 1788–1794. [Google Scholar] [CrossRef]
- Ventura, C.V.; Zin, A.; Paula Freitas, B.; Ventura, L.O.; Rocha, C.; Costa, F.; Nery, N., Jr.; De Senna, T.C.R.; Lopes Moreira, M.E.; Maia, M.; et al. Ophthalmological manifestations in congenital Zika syndrome in 469 Brazilian children. J. AAPOS 2021, 25, 158.e1–158.e8. [Google Scholar] [CrossRef]
- Vercosa, I.; Carneiro, P.; Vercosa, R.; Girao, R.; Ribeiro, E.M.; Pessoa, A.; Almeida, N.G.; Vercosa, P.; Tartarella, M.B. The visual system in infants with microcephaly related to presumed congenital Zika syndrome. J. AAPOS 2017, 21, 300–304.e1. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Dias, J.R.; Ventura, C.V.; Borba, P.D.; de Paula Freitas, B.; Pierroti, L.C.; do Nascimento, A.P.; de Moraes, N.S.B.; Maia, M.; Belfort, R., Jr. Infants with Congenital Zika Syndrome and Ocular Findings from Sao Paulo, Brazil: Spread of Infection. Retin. Cases Brief Rep. 2018, 12, 382–386. [Google Scholar] [CrossRef] [PubMed]
- De Paula Freitas, B.; Ko, A.I.; Khouri, R.; Mayoral, M.; Henriques, D.F.; Maia, M.; Belfort, R., Jr. Glaucoma and Congenital Zika Syndrome. Ophthalmology 2017, 124, 407–408. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.M.; Medronho, R.A.; Cunha, A. Zika virus in Brazil and worldwide: A narrative review. Paediatr. Int. Child Health 2021, 41, 28–35. [Google Scholar] [CrossRef]
- Maharajan, M.K.; Ranjan, A.; Chu, J.F.; Foo, W.L.; Chai, Z.X.; Lau, E.Y.; Ye, H.M.; Theam, X.J.; Lok, Y.L. Zika Virus Infection: Current Concerns and Perspectives. Clin. Rev. Allergy Immunol. 2016, 51, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Hua, S.; Chen, H.R.; Ouyang, Z.; Einkauf, K.; Tse, S.; Ard, K.; Ciaranello, A.; Yawetz, S.; Sax, P.; et al. Transcriptional Changes during Naturally Acquired Zika Virus Infection Render Dendritic Cells Highly Conducive to Viral Replication. Cell Rep. 2017, 21, 3471–3482. [Google Scholar] [CrossRef] [Green Version]
- Lugo-Villarino, G.; Troegeler, A.; Balboa, L.; Lastrucci, C.; Duval, C.; Mercier, I.; Benard, A.; Capilla, F.; Al Saati, T.; Poincloux, R.; et al. The C-Type Lectin Receptor DC-SIGN Has an Anti-Inflammatory Role in Human M(IL-4) Macrophages in Response to Mycobacterium tuberculosis. Front. Immunol. 2018, 9, 1123. [Google Scholar] [CrossRef] [Green Version]
- Tsou, W.I.; Nguyen, K.Q.; Calarese, D.A.; Garforth, S.J.; Antes, A.L.; Smirnov, S.V.; Almo, S.C.; Birge, R.B.; Kotenko, S.V. Receptor tyrosine kinases, TYRO3, AXL, and MER, demonstrate distinct patterns and complex regulation of ligand-induced activation. J. Biol. Chem 2014, 289, 25750–25763. [Google Scholar] [CrossRef] [Green Version]
- Nowakowski, T.J.; Pollen, A.A.; Di Lullo, E.; Sandoval-Espinosa, C.; Bershteyn, M.; Kriegstein, A.R. Expression Analysis Highlights AXL as a Candidate Zika Virus Entry Receptor in Neural Stem Cells. Cell Stem Cell 2016, 18, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Bayless, N.L.; Greenberg, R.S.; Swigut, T.; Wysocka, J.; Blish, C.A. Zika Virus Infection Induces Cranial Neural Crest Cells to Produce Cytokines at Levels Detrimental for Neurogenesis. Cell Host Microbe 2016, 20, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Ma, Z.; Hu, W.; Wang, D.; Gong, B.; Fan, C.; Jiang, S.; Li, T.; Gao, J.; Yang, Y. Molecular insights of Gas6/TAM in cancer development and therapy. Cell Death Dis. 2017, 8, e2700. [Google Scholar] [CrossRef]
- Roach, T.; Alcendor, D.J. Zika virus infection of cellular components of the blood-retinal barriers: Implications for viral associated congenital ocular disease. J. Neuroinflamm. 2017, 14, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Yang, M.; Azar, S.R.; Soong, L.; Weaver, S.C.; Sun, J.; Chen, Y.; Rossi, S.L.; Cai, J. Viral Retinopathy in Experimental Models of Zika Infection. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4355–4365. [Google Scholar] [CrossRef] [PubMed]
- Vielle, N.J.; Zumkehr, B.; Garcia-Nicolas, O.; Blank, F.; Stojanov, M.; Musso, D.; Baud, D.; Summerfield, A.; Alves, M.P. Silent infection of human dendritic cells by African and Asian strains of Zika virus. Sci. Rep. 2018, 8, 5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonin, Y.; Erkilic, N.; Damodar, K.; Cle, M.; Desmetz, C.; Bollore, K.; Taleb, M.; Torriano, S.; Barthelemy, J.; Dubois, G.; et al. Zika virus induces strong inflammatory responses and impairs homeostasis and function of the human retinal pigment epithelium. eBioMedicine 2019, 39, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.R.; Roby, J.A.; Dobyns, W.B.; Rajagopal, L.; Gale, M., Jr.; Adams Waldorf, K.M. Immune Evasion Strategies Used by Zika Virus to Infect the Fetal Eye and Brain. Viral Immunol. 2020, 33, 22–37. [Google Scholar] [CrossRef]
- Beaver, J.T.; Mills, L.K.; Swieboda, D.; Lelutiu, N.; Esser, E.S.; Antao, O.Q.; Scountzou, E.; Williams, D.T.; Papaioannou, N.; Littauer, E.Q.; et al. Zika virus-induced neuro-ocular pathology in immunocompetent mice correlates with anti-ganglioside autoantibodies. Hum. Vaccines Immunother. 2020, 16, 2092–2108. [Google Scholar] [CrossRef]
- Hoffmann, S.; He, S.; Ehren, M.; Ryan, S.J.; Wiedemann, P.; Hinton, D.R. MMP-2 and MMP-9 secretion by rpe is stimulated by angiogenic molecules found in choroidal neovascular membranes. Retina 2006, 26, 454–461. [Google Scholar] [CrossRef]
- Young, A.T.L.; Moore, R.B.; Murray, A.G.; Mullen, J.C.; Lakey, J.R.T. Assessment of Different Transfection Parameters in Efficiency Optimization. Cell Transpl. 2004, 13, 179–185. [Google Scholar] [CrossRef] [Green Version]
- McDonald, E.M.; Anderson, J.; Wilusz, J.; Ebel, G.D.; Brault, A.C. Zika Virus Replication in Myeloid Cells during Acute Infection Is Vital to Viral Dissemination and Pathogenesis in a Mouse Model. J. Virol. 2020, 94, e00838-20. [Google Scholar] [CrossRef]
- Wen, C.; Yu, Y.; Gao, C.; Qi, X.; Cardona, C.J.; Xing, Z. Concomitant pyroptotic and apoptotic cell death triggered in macrophages infected by Zika virus. PLoS ONE 2022, 17, e0257408. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, E.; Orovou, E.; Sarella, A.; Iliadou, M.; Rigas, N.; Palaska, E.; Iatrakis, G.; Dagla, M. Zika Virus and the Risk of Developing Microcephaly in Infants: A Systematic Review. Int. J. Environ. Res. Public Health 2020, 17, 3806. [Google Scholar] [CrossRef] [PubMed]
- King, E.L.; Irigoyen, N. Zika Virus and Neuropathogenesis: The Unanswered Question of Which Strain Is More Prone to Causing Microcephaly and Other Neurological Defects. Front. Cell. Neurosci. 2021, 15, 695106. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Gorshkov, K.; Lee, E.M.; Xu, M.; Cheng, Y.S.; Sun, N.; Soheilian, F.; de Val, N.; Ming, G.; Song, H.; et al. Zika Virus-Induced Neuronal Apoptosis via Increased Mitochondrial Fragmentation. Front. Microbiol. 2020, 11, 598203. [Google Scholar] [CrossRef]
- Kuadkitkan, A.; Wikan, N.; Sornjai, W.; Smith, D.R. Zika virus and microcephaly in Southeast Asia: A cause for concern? J. Infect. Public Health 2020, 13, 11–15. [Google Scholar] [CrossRef]
- Gabriel, E.; Ramani, A.; Karow, U.; Gottardo, M.; Natarajan, K.; Gooi, L.M.; Goranci-Buzhala, G.; Krut, O.; Peters, F.; Nikolic, M.; et al. Recent Zika Virus Isolates Induce Premature Differentiation of Neural Progenitors in Human Brain Organoids. Cell Stem Cell 2017, 20, 397–406.e5. [Google Scholar] [CrossRef] [Green Version]
- Rosa-Fernandes, L.; Cugola, F.R.; Russo, F.B.; Kawahara, R.; de Melo Freire, C.C.; Leite, P.E.C.; Bassi Stern, A.C.; Angeli, C.B.; de Oliveira, D.B.L.; Melo, S.R.; et al. Zika Virus Impairs Neurogenesis and Synaptogenesis Pathways in Human Neural Stem Cells and Neurons. Front. Cell. Neurosci. 2019, 13, 64. [Google Scholar] [CrossRef]
- Li, C.; Xu, D.; Ye, Q.; Hong, S.; Jiang, Y.; Liu, X.; Zhang, N.; Shi, L.; Qin, C.F.; Xu, Z. Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 2016, 19, 672. [Google Scholar] [CrossRef] [Green Version]
- Komarasamy, T.V.; Adnan, N.A.A.; James, W.; Balasubramaniam, V. Zika Virus Neuropathogenesis: The Different Brain Cells, Host Factors and Mechanisms Involved. Front. Immunol. 2022, 13, 773191. [Google Scholar] [CrossRef]
- Williams, A.L.; Bohnsack, B.L. Neural crest derivatives in ocular development: Discerning the eye of the storm. Birth Defects Res. C Embryo Today 2015, 105, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Akula, M.; Park, J.W.; West-Mays, J.A. Relationship between neural crest cell specification and rare ocular diseases. J. Neurosci. Res. 2019, 97, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Tamm, E.R. Development of the iridocorneal angle and congenital glaucoma. Ophthalmologe 2011, 108, 610–614, 616–617. [Google Scholar] [CrossRef] [PubMed]
- Tawara, A.; Inomata, H. Developmental immaturity of the trabecular meshwork in juvenile glaucoma. Am. J. Ophthalmol. 1984, 98, 82–97. [Google Scholar] [CrossRef]
- Alkatan, H.M.; Bedaiwi, K.M.; Al-Faky, Y.H.; Maktabi, A.M.Y. Demographics and histopathological characteristics of enucleated microphthalmic globes. Sci. Rep. 2022, 12, 5283. [Google Scholar] [CrossRef]
- Verma, A.S.; Fitzpatrick, D.R. Anophthalmia and microphthalmia. Orphanet J. Rare Dis. 2007, 2, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.L.; Bohnsack, B.L. The Ocular Neural Crest: Specification, Migration, and Then What? Front. Cell Dev. Biol. 2020, 8, 595896. [Google Scholar] [CrossRef]
- Medina-Martinez, O.; Brownell, I.; Amaya-Manzanares, F.; Hu, Q.; Behringer, R.R.; Jamrich, M. Severe defects in proliferation and differentiation of lens cells in Foxe3 null mice. Mol. Cell. Biol. 2005, 25, 8854–8863. [Google Scholar] [CrossRef] [Green Version]
- El Costa, H.; Gouilly, J.; Mansuy, J.M.; Chen, Q.; Levy, C.; Cartron, G.; Veas, F.; Al-Daccak, R.; Izopet, J.; Jabrane-Ferrat, N. ZIKA virus reveals broad tissue and cell tropism during the first trimester of pregnancy. Sci. Rep. 2016, 6, 35296. [Google Scholar] [CrossRef]
- Hirsch, A.J.; Roberts, V.H.J.; Grigsby, P.L.; Haese, N.; Schabel, M.C.; Wang, X.; Lo, J.O.; Liu, Z.; Kroenke, C.D.; Smith, J.L.; et al. Zika virus infection in pregnant rhesus macaques causes placental dysfunction and immunopathology. Nat. Commun. 2018, 9, 263. [Google Scholar] [CrossRef]
- Rabelo, K.; de Souza, L.J.; Salomao, N.G.; Machado, L.N.; Pereira, P.G.; Portari, E.A.; Basilio-de-Oliveira, R.; Dos Santos, F.B.; Neves, L.D.; Morgade, L.F.; et al. Zika Induces Human Placental Damage and Inflammation. Front. Immunol. 2020, 11, 2146. [Google Scholar] [CrossRef]
- Singh, P.K.; Guest, J.M.; Kanwar, M.; Boss, J.; Gao, N.; Juzych, M.S.; Abrams, G.W.; Yu, F.S.; Kumar, A. Zika virus infects cells lining the blood-retinal barrier and causes chorioretinal atrophy in mouse eyes. JCI Insight 2017, 2, e92340. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Khatri, I.; Jha, A.; Pretto, C.D.; Spindler, K.R.; Arumugaswami, V.; Giri, S.; Kumar, A.; Bhasin, M.K. Determination of system level alterations in host transcriptome due to Zika virus (ZIKV) Infection in retinal pigment epithelium. Sci. Rep. 2018, 8, 11209. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Singh, S.; Farr, D.; Kumar, A. Interferon-stimulated gene 15 (ISG15) restricts Zika virus replication in primary human corneal epithelial cells. Ocul. Surf. 2019, 17, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Kasetti, R.B.; Zode, G.S.; Goyal, A.; Juzych, M.S.; Kumar, A. Zika Virus Infects Trabecular Meshwork and Causes Trabeculitis and Glaucomatous Pathology in Mouse Eyes. mSphere 2019, 4, e00173-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueta, M.; Kawai, T.; Yokoi, N.; Akira, S.; Kinoshita, S. Contribution of IPS-1 to polyI:C-induced cytokine production in conjunctival epithelial cells. Biochem. Biophys. Res. Commun. 2011, 404, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Mai, K.; Chui, J.J.; Di Girolamo, N.; McCluskey, P.J.; Wakefield, D. Role of toll-like receptors in human iris pigment epithelial cells and their response to pathogen-associated molecular patterns. J. Inflamm. 2014, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Ryan, F.J.; Carr, J.M.; Furtado, J.M.; Ma, Y.; Ashander, L.M.; Simoes, M.; Oliver, G.F.; Granado, G.B.; Dawson, A.C.; Michael, M.Z.; et al. Zika Virus Infection of Human Iris Pigment Epithelial Cells. Front. Immunol. 2021, 12, 644153. [Google Scholar] [CrossRef]
- De Groot-Mijnes, J.D.F.; Chan, A.S.Y.; Chee, S.P.; Verjans, G. Immunopathology of Virus-Induced Anterior Uveitis. Ocul. Immunol. Inflamm. 2018, 26, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Kalogeropoulos, D.; Sung, V.C. Pathogenesis of Uveitic Glaucoma. J. Curr. Glaucoma Pract. 2018, 12, 125–138. [Google Scholar] [CrossRef]
- Ventura, C.V.; Ventura, L.O. Ophthalmologic Manifestations Associated With Zika Virus Infection. Pediatrics 2018, 141, S161–S166. [Google Scholar] [CrossRef] [Green Version]
- Sherman, K.E.; Rouster, S.D.; Kong, L.X.; Aliota, M.T.; Blackard, J.T.; Dean, G.E. Zika virus replication and cytopathic effects in liver cells. PLoS ONE 2019, 14, e0214016. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.P.; Tsai, K.N.; Luo, Y.C.; Chung, P.J.; Su, Y.W.; Teng, Y.; Wu, M.S.; Lin, Y.F.; Lai, C.Y.; Chuang, T.H.; et al. Establishment of a mouse model for the complete mosquito-mediated transmission cycle of Zika virus. PLoS Negl. Trop. Dis. 2018, 12, e0006417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manangeeswaran, M.; Kielczewski, J.L.; Sen, H.N.; Xu, B.C.; Ireland, D.D.C.; McWilliams, I.L.; Chan, C.C.; Caspi, R.R.; Verthelyi, D. ZIKA virus infection causes persistent chorioretinal lesions. Emerg. Microbes Infect. 2018, 7, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamer, W.D.; Clark, A.F. The many faces of the trabecular meshwork cell. Exp. Eye Res. 2017, 158, 112–123. [Google Scholar] [CrossRef] [Green Version]
- McArthur, M.A. Zika Virus: Recent Advances towards the Development of Vaccines and Therapeutics. Viruses 2017, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, M.; Chigbu, D.I.; Crumley, B.L.; Sharma, R.; Pustylnikov, S.; Crilley, T.; Ginwala, R.; Loonawat, R.; Joseph, J.; Sales, D.; et al. Human Acute and Chronic Viruses: Host-Pathogen Interactions and Therapeutics. In Advanced Concepts in Human Immunology: Prospects for Disease Control; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–120. [Google Scholar]
- Bodh, S.A.; Kumar, V.; Raina, U.K.; Ghosh, B.; Thakar, M. Inflammatory glaucoma. Oman J. Ophthalmol. 2011, 4, 3–9. [Google Scholar] [CrossRef]
- Badawi, A.H.; Al-Muhaylib, A.A.; Al Owaifeer, A.M.; Al-Essa, R.S.; Al-Shahwan, S.A. Primary congenital glaucoma: An updated review. Saudi J. Ophthalmol. 2019, 33, 382–388. [Google Scholar] [CrossRef]
- Tavallali, A.; Yannuzzi, L.A. Idiopathic Multifocal Choroiditis. J. Ophthalmic Vis. Res. 2016, 11, 429–432. [Google Scholar] [CrossRef]
- Baz, M.; Boivin, G. Antiviral Agents in Development for Zika Virus Infections. Pharmaceuticals 2019, 12, 101. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Anterior Segment | Neuro-Ophthalmic | Posterior Segment |
|---|---|---|
| Nonpurulent conjunctivitis [62]. Anterior uveitis (bilateral, nongranulomatous iridocyclitis with or without elevated intraocular pressure [57,64]. | Papilledema [66]. Ophthalmoplegia [5,66]. Ocular flutter [67]. | Maculopathy with outer retinal layer and RPE disruption [57,61]. Multifocal choroiditis [65]. |
| Anterior Segment | Neuro-Ophthalmic | Posterior Segment |
|---|---|---|
| Lens subluxation [61,63]. Cataract [63,76,78]. Intraocular calcifications [63,76,78]. Congenital glaucoma [80] Microphthalmia [81,82]. Corneal ectasia [83]. Iris coloboma [63,84]. | Strabismus [79,85]. Horizontal nystagmus [79,85]. Exophoria/esophoria [79,85]. Loss of pupillary response [79,85]. Disc hypoplasia [79,85]. Disc pallor [79,85]. Enlarged cup-to-disc ratio [79,85]. | Macular pigment mottling [5,79]. Chorioretinal atrophy either macular, paramacular, or peripheral [5,79]. Retinal hemorrhage [86]. Vascular tortuosity [85]. Early termination of retinal vasculature [85]. Washed out peripheral retina with hypoluscent spot [85]. Photoreceptor, RPE thinning with pigment loss, and choroidal thinning [87,88]. |
| Primary Targets | Receptors | Mechanisms | Outcomes |
|---|---|---|---|
| Neural progenitor cells | AXL [99]. | Apoptosis of ZIKV-infected NPCs. Reduced proliferation of NPCs. Premature differentiation of NPCs [113,115]. | Microcephaly [115]. |
| Neural crest cells | AXL [100]. | ZIKV infection of NCCs during the developmental stages of the fetus causes abnormal migration, proliferation, and differentiation of NCCs [100,120]. The loss of NCCs associated with ZIKV infection can contribute to the disruption of the formation of the optic fissure, resulting in failure to close the optic fissure [86,120]. ZIKV infection of NCCs will result in abnormal proliferation and differentiation of cells required for the normal development of the cornea [121]. ZIKV infection of NCCs during the developmental stages of the fetus can lead to abnormal morphogenesis of the trabecular meshwork [100,122,123]. | Microphthalmia [106,120,124,125]. Iris coloboma [7,84,120,126]. Corneal ectasia [83]. Congenital glaucoma [93,100,122,123]. |
| Mesenchymal stem cells | AXL [101]. | ZIKV infects mesenchymal stem cells, resulting in impaired proliferation and differentiation of cells required for the development of the crystalline lens [30,100,121,127,128]. | Congenital cataract [30,100,121,127,128]. |
| Placental endothelial cells and trophoblasts | TIM-1, AXL, TYRO3 [27]. | Destruction of ZIKV-infected placenta [129,130]. Compromised maternal-fetal interface [129,130]. | Placental insufficiency causes restricted growth of the fetus and disruption of neurodevelopment of the fetus [129]. Facilitate access of ZIKV to the fetus [129,130]. |
| Blood retinal barrier cells (Retinal vascular endothelial cells and retinal pigment epithelial cells) | AXL, TYRO3, TIM-1, RIG-I/MDA5, TLR3 [20]. | ZIKV infected BRB cells induce inflammation that damages the BRB and facilitate influx of effector immune cells into the retina [131,132]. | Chorioretinitis, macular pigment mottling, chorioretinal atrophy, and maculopathy [131,132]. |
| Cornea epithelial cells | TLR3, RIG-I, MDA5 [133]. | ZIKV-infected corneal epithelium induces antiviral response and immune-mediated inflammation [133]. | Keratitis [133]. |
| Trabecular meshwork cells | RIG-I, TLR3 [134]. | ZIKV-infected trabecular meshwork cells secrete cytokines and chemokines that promote inflammation via the recruitment of effector immune cells such as Th1 cells to the trabecular meshwork [134]. | Trabeculitis [134]. |
| Conjunctival epithelial cells | RIG-I/MDA5, TLR3 [135]. | ZIKV-infected conjunctival epithelial cells induce an immune-mediated inflammatory response [135]. | Nonpurulent conjunctivitis [135]. |
| Iris pigment epithelium | TLR3 [136]. | Immune-mediated inflammation triggered in response to ZIKV-infected iris pigment epithelium. ZIKV-infected blood aqueous barrier (BAB) cells induce inflammation that damages the BAB and facilitate influx of effector immune cells into the anterior uvea. Influx of ZIKV-infected monocytes acting as Trojan horses [137,138,139]. | Anterior uveitis with or without raised intraocular pressure [137,138,139]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labib, B.A.; Chigbu, D.I. Pathogenesis and Manifestations of Zika Virus-Associated Ocular Diseases. Trop. Med. Infect. Dis. 2022, 7, 106. https://doi.org/10.3390/tropicalmed7060106
Labib BA, Chigbu DI. Pathogenesis and Manifestations of Zika Virus-Associated Ocular Diseases. Tropical Medicine and Infectious Disease. 2022; 7(6):106. https://doi.org/10.3390/tropicalmed7060106
Chicago/Turabian StyleLabib, Bisant A., and DeGaulle I. Chigbu. 2022. "Pathogenesis and Manifestations of Zika Virus-Associated Ocular Diseases" Tropical Medicine and Infectious Disease 7, no. 6: 106. https://doi.org/10.3390/tropicalmed7060106
APA StyleLabib, B. A., & Chigbu, D. I. (2022). Pathogenesis and Manifestations of Zika Virus-Associated Ocular Diseases. Tropical Medicine and Infectious Disease, 7(6), 106. https://doi.org/10.3390/tropicalmed7060106