
SARS-CoV-2 Post-Infection and Sepsis by Saccharomyces cerevisiae: A Fatal Case Report—Focus on Fungal Susceptibility and Potential Virulence Attributes

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Report
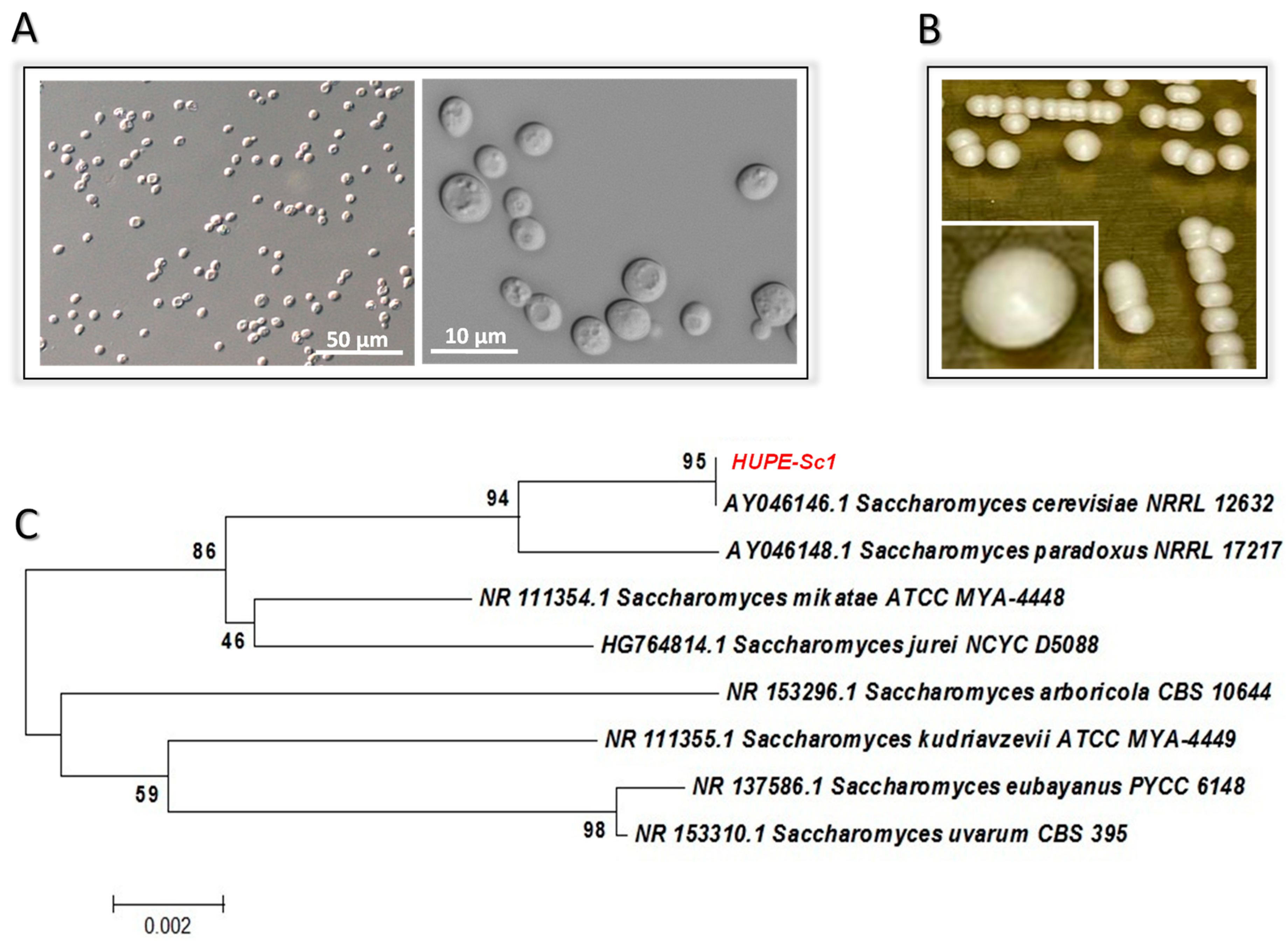
2.2. Yeast-like Identification
2.3. Antifungal Susceptibility Assay
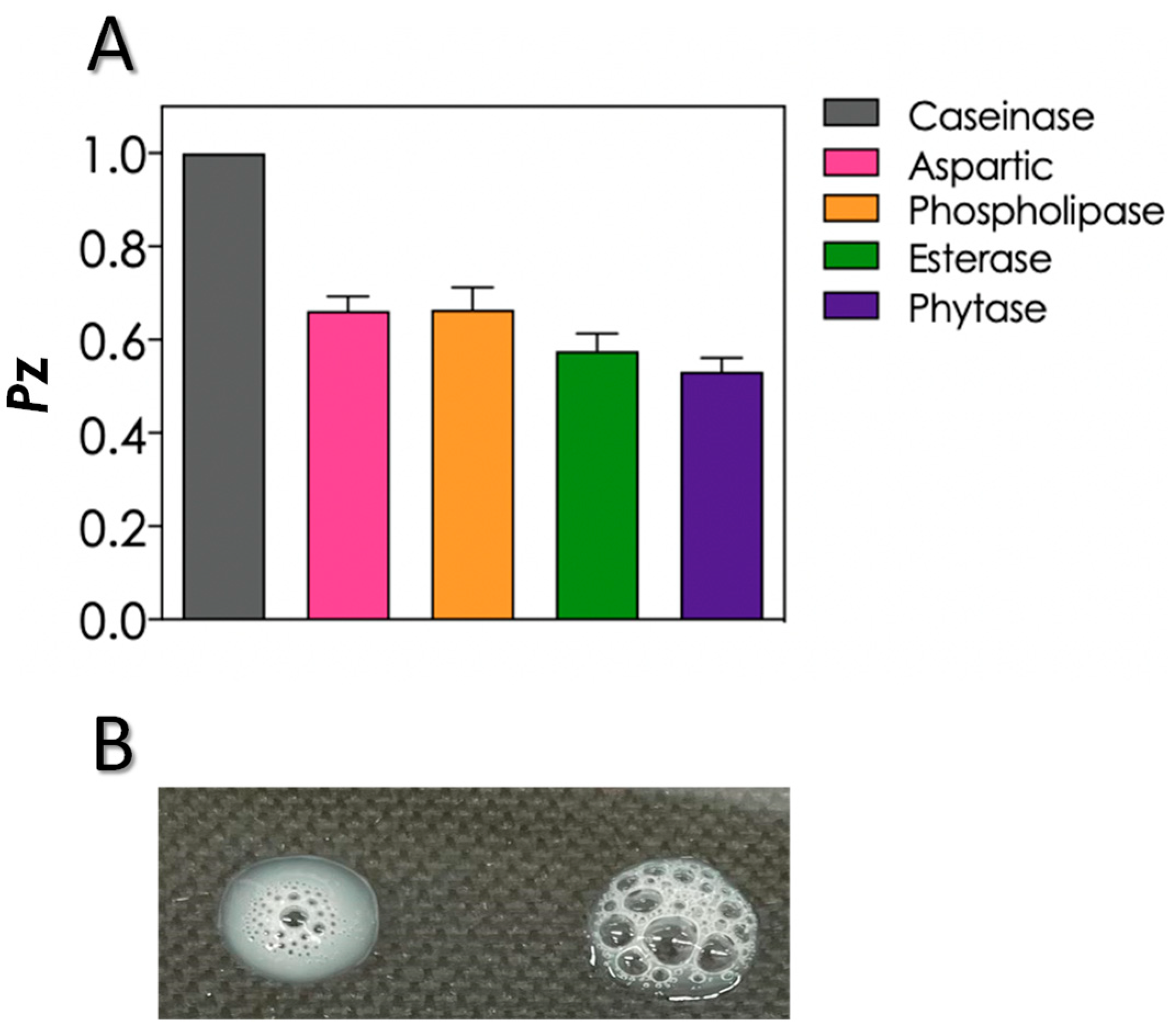
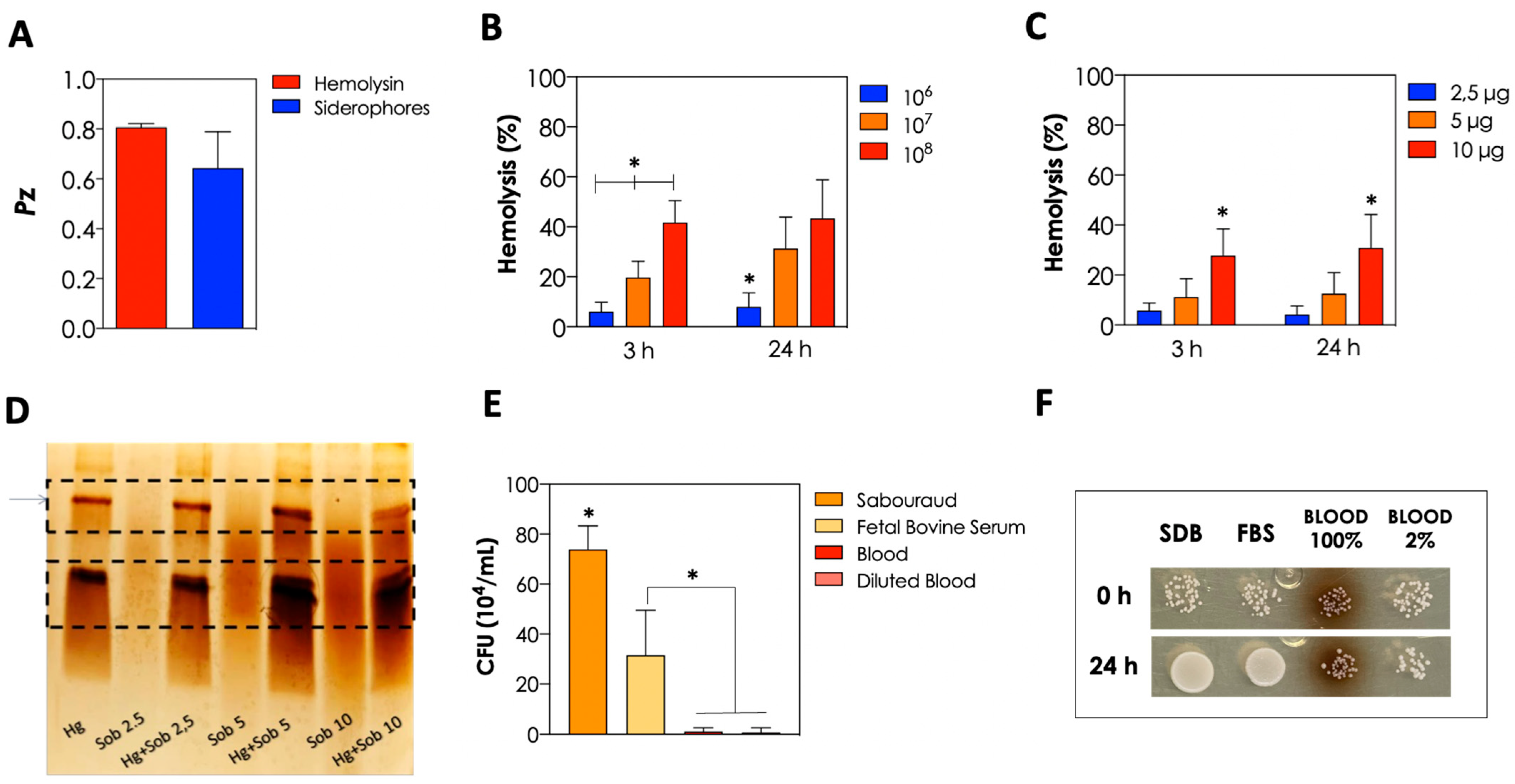
2.4. Detection of Extracellular Molecules
2.5. Culture Supernatant Harvesting
2.6. Hemolysis Assay
2.7. Hemoglobin Hydrolysis
2.8. Fungal Growth in Different Nutrient Sources
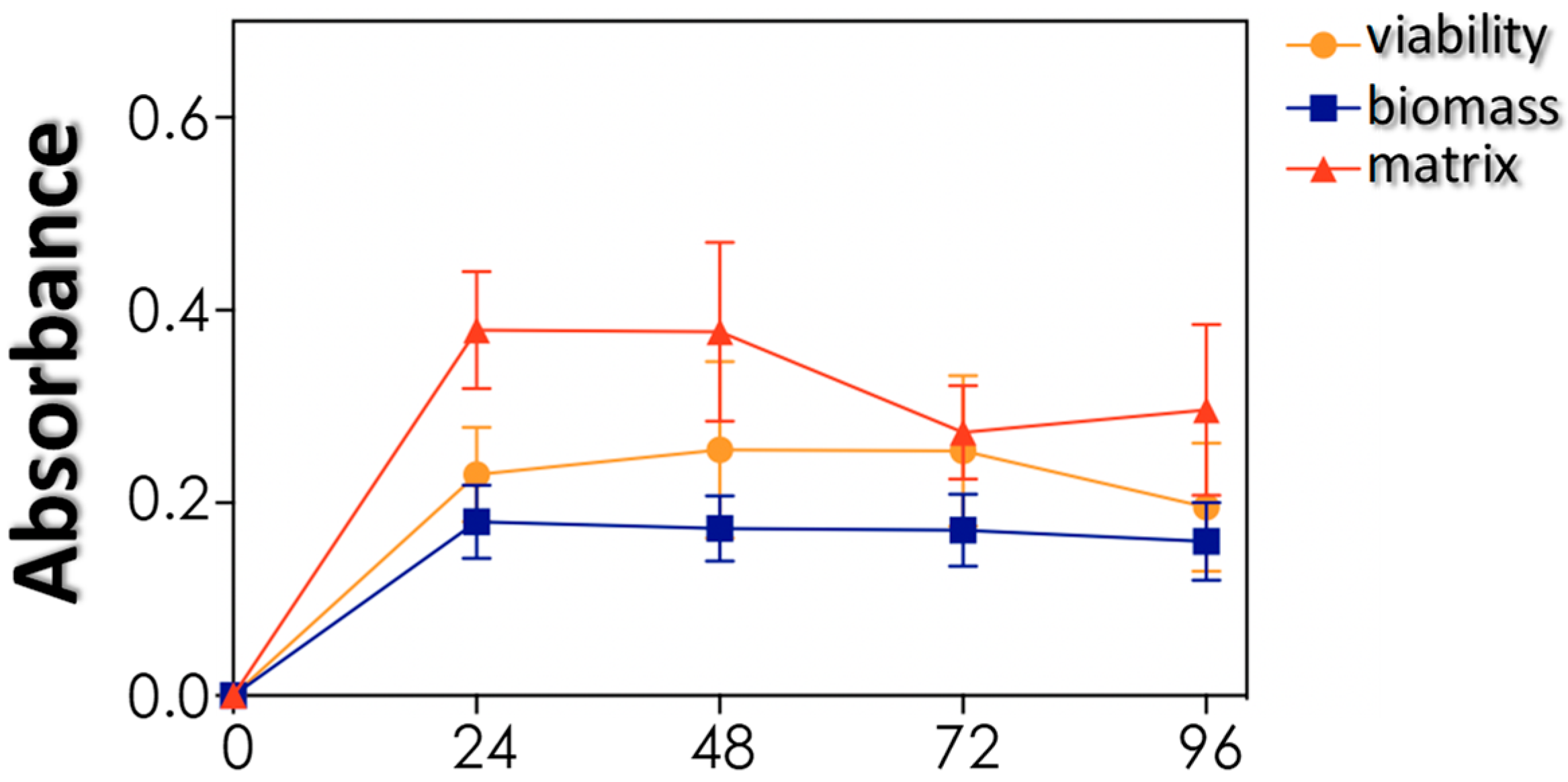
2.9. Biofilm Formation
2.10. Antifungal Susceptibility of Biofilm-Forming Cells
2.11. In Vivo Infection in Tenebrio molitor Larvae
2.12. Impact of Immunosuppression on Larvae Survival
2.13. Statistics
3. Results
3.1. Yeast Identification by Biochemical and Molecular Approaches
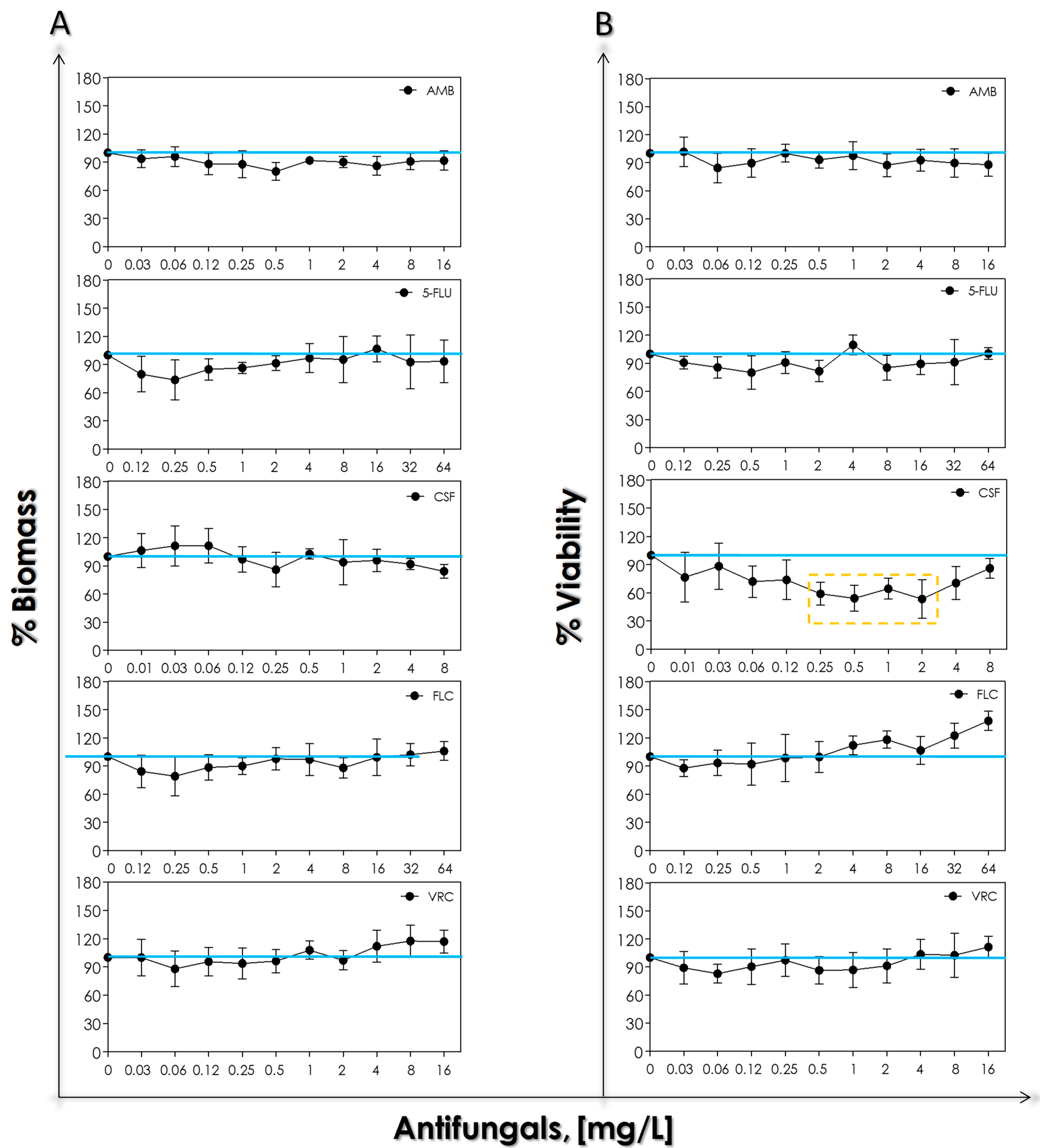
3.2. Susceptibility Profile to Antifungal Agents
3.3. Production of Biologically Active Extracellular Molecules and Growth in Different Nutrient Sources
3.4. Biofilm Formation and Impact of Antifungals on Mature Biofilms
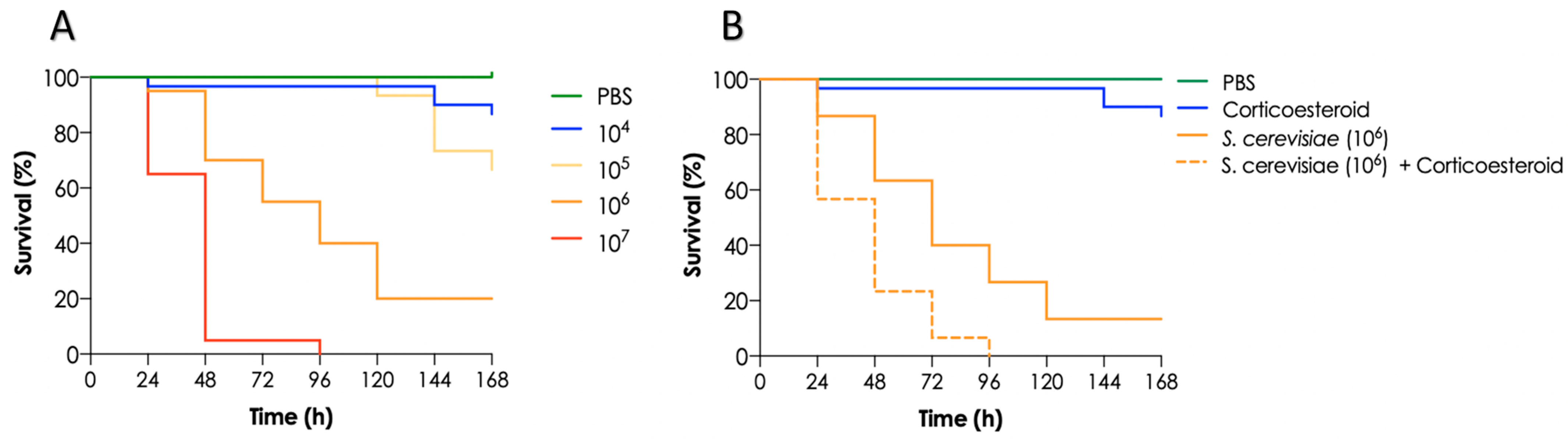
3.5. Mortality of T. molitor Larvae Infected with S. cerevisiae and Impact of Immunosuppression on Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldman, C.; Anderson, R. The role of co-infections and secondary infections in patients with COVID-19. Pneumonia 2021, 13, 5. [Google Scholar] [CrossRef]
- Musuuza, J.S.; Watson, L.; Parmasad, V.; Putman-Buehler, N.; Christensen, L.; Safdar, N. Prevalence and outcomes of co-infection and superinfection with SARS-CoV-2 and other pathogens: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0251170. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torrado, R.; Querol, A. Opportunistic strains of Saccharomyces cerevisiae: A potential risk sold in food products. Front. Microbiol. 2015, 6, 1522. [Google Scholar] [CrossRef]
- Atıcı, S.; Soysal, A.; Karadeniz Cerit, K.; Yılmaz, Ş.; Aksu, B.; Kıyan, G.; Bakır, M. Catheter-related Saccharomyces cerevisiae fungemia following Saccharomyces boulardii probiotic treatment: In a child in intensive care unit and review of the literature. Med. Mycol. Case Rep. 2017, 15, 33–35. [Google Scholar] [CrossRef]
- Ventoulis, I.; Sarmourli, T.; Amoiridou, P.; Mantzana, P.; Exindari, M.; Gioula, G.; Vyzantiadis, T.A. Bloodstream infection by Saccharomyces cerevisiae in two COVID-19 patients after receiving supplementation of Saccharomyces in the ICU. J. Fungi 2020, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.; Lima, L.; Pedra, T.; Assumpção, A.; Morgado, S.; Mascarenhas, L. Bloodstream infection by Saccharomyces cerevisiae in a COVID-19 patient receiving probiotic supplementation in the ICU in Brazil. Access Microbiol. 2021, 3, 000250. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 3rd ed.; Approved Standard, CLSI Document M27A3; CLSI: Wayne, PA, USA, 2008; Volume 28, p. 40. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Third Informational Supplement, CLSI Document M27-S3; CLSI: Wayne, PA, USA, 2008; p. 28. [Google Scholar]
- Ruchel, R.; Tegeler, R.; Trost, M. A comparison of secretory proteinases from different strains of Candida albicans. Sabouraudia 1982, 20, 233–244. [Google Scholar] [CrossRef]
- Ziccardi, M.; Souza, L.O.; Gandra, R.M.; Galdino, A.C.; Baptista, A.R.; Nunes, A.P.; Ribeiro, M.A.; Branquinha, M.H.; Santos, A.L.S. Candida parapsilosis (sensu lato) isolated from hospitals located in the Southeast of Brazil: Species distribution, antifungal susceptibility and virulence attributes. Int. J. Med. Microbiol. 2015, 305, 848–859. [Google Scholar] [CrossRef]
- Price, M.F.; Wilkinson, I.D.; Gentry, L.O. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia 1982, 20, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Aktas, E.; Yigit, N.; Ayyildiz, A. Esterase activity in various Candida species. J. Int. Med. Res. 2002, 30, 322–324. [Google Scholar] [CrossRef]
- Tsang, P.W. Differential phytate utilization in Candida species. Mycopathologia 2011, 172, 473–479. [Google Scholar] [CrossRef]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef]
- Lakshmanan, V.; Shantharaj, D.; Li, G.; Seyfferth, A.L.; Sherrier, D.J.; Bais, H.P. A natural rice rhizospheric bacterium abates arsenic accumulation in rice (Oryza sativa L.). Planta 2015, 242, 1037–1050. [Google Scholar] [CrossRef]
- Ramos, L.S.; Figueiredo-Carvalho, M.H.G.; Silva, L.N.; Siqueira, N.L.M.; Lima, J.C.; Oliveira, S.S.; Almeida-Paes, R.; Zancopé-Oliveira, R.M.; Azevedo, F.S.; Ferreira, A.L.P.; et al. The threat called Candida haemulonii species complex in Rio de Janeiro State, Brazil: Focus on antifungal resistance and virulence attributes. J. Fungi 2022, 8, 574. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Evans, B.C.; Nelson, C.E.; Yu, S.S.; Beavers, K.R.; Kim, A.J.; Li, H.; Nelson, H.M.; Giorgio, T.D.; Duvall, C.L. Ex vivo red blood cell hemolysis assay for the evaluation of pH-responsive endosomolytic agents for cytosolic delivery of biomacromolecular drugs. J. Vis. Exp. 2013, 73, e50166. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Ramos, L.S.; Oliveira, S.S.C.; Braga-Silva, L.A.; Branquinha, M.H.; Santos, A.L.S. Secreted aspartyl peptidases by the emerging, opportunistic and multidrug-resistant fungal pathogens comprising the Candida haemulonii complex. Fungal Biol. 2020, 124, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Souto, X.M.; Ramos, L.S.; Branquinha, M.H.; Santos, A.L.S. Identification of cell-associated and secreted serine-type peptidases in multidrug-resistant emergent pathogens belonging to the Candida haemulonii complex. Folia Microbiol. 2019, 64, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, N.Y.; Kang, S.Y.; Kim, K.J. Artemisia princeps inhibits biofilm formation and virulence-factor expression of antibiotic-resistant bacteria. BioMed Res. Int. 2015, 2015, 239519. [Google Scholar] [CrossRef] [PubMed]
- Ramos, L.S.; Oliveira, S.S.C.; Souto, X.M.; Branquinha, M.H.; Santos, A.L.S. Planktonic growth and biofilm formation profiles in Candida haemulonii species complex. Med. Mycol. 2017, 55, 785–789. [Google Scholar] [CrossRef] [PubMed]
- de Souza, P.C.; Morey, A.T.; Castanheira, G.M.; Bocate, K.P.; Panagio, L.A.; Ito, F.A.; Furlaneto, M.C.; Yamada-Ogatta, S.F.; Costa, I.N.; Mora-Montes, H.M.; et al. Tenebrio molitor (Coleoptera: Tenebrionidae) as an alternative host to study fungal infections. J. Microbiol. Methods 2015, 118, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.P.; Entwistle, F.; Coote, P.J. Effective immunosuppression with dexamethasone phosphate in the Galleria mellonella larva infection model resulting in enhanced virulence of Escherichia coli and Klebsiella pneumoniae. Med. Microbiol. Immunol. 2016, 205, 333–343. [Google Scholar] [CrossRef]
- Pérez-Cantero, A.; Thomson, P.; Paredes, K.; Guarro, J.; Capilla, J. Antifungal susceptibility of Saccharomyces cerevisiae and therapy in a murine model of disseminated infection. Rev. Iberoam. Micol. 2019, 36, 37–40. [Google Scholar] [CrossRef]
- Castanheira, M.; Messer, S.A.; Jones, R.N.; Farrell, D.J.; Pfaller, M.A. Activity of echinocandins and triazoles against a contemporary (2012) worldwide collection of yeast and moulds collected from invasive infections. Int. J. Antimicrob. Agents 2014, 44, 320–326. [Google Scholar] [CrossRef]
- Echeverría-Irigoyen, M.J.; Eraso, E.; Cano, J.; Gomáriz, M.; Guarro, J.; Quindós, G. Saccharomyces cerevisiae vaginitis: Microbiology and in vitro antifungal susceptibility. Mycopathologia 2011, 172, 201–205. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Bijie, H.; Dzierzanowska, D.; Klimko, N.N.; Letscher-Bru, V.; Lisalova, M.; Muehlethaler, K.; et al. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: 10.5-year analysis of susceptibilities of noncandidal yeast species to fluconazole and voriconazole determined by CLSI standardized disk diffusion testing. J. Clin. Microbiol. 2009, 47, 117–123. [Google Scholar] [CrossRef]
- de Llanos, R.; Fernández-Espinar, M.T.; Querol, A. A comparison of clinical and food Saccharomyces cerevisiae isolates on the basis of potential virulence factors. Antonie Van Leeuwenhoek 2006, 90, 221–231. [Google Scholar] [CrossRef]
- Imre, A.; Kovács, R.; Pázmándi, K.; Nemes, D.; Jakab, Á.; Fekete, T.; Rácz, H.V.; Dóczi, I.; Bácskay, I.; Gácser, A.; et al. Virulence factors and in-host selection on phenotypes in infectious probiotic yeast isolates (Saccharomyces ‘boulardii’). J. Fungi 2021, 7, 746. [Google Scholar] [CrossRef]
- Ries, E.F.; Alves Macedo, G. Improvement of phytase activity by a new Saccharomyces cerevisiae strain using statistical optimization. Enzym. Res. 2011, 2011, 796394. [Google Scholar] [CrossRef]
- Nishimoto, T.; Watanabe, T.; Furuta, M.; Kataoka, M.; Kishida, M. Roles of catalase and trehalose in the protection from hydrogen peroxide toxicity in Saccharomyces cerevisiae. Biocontrol Sci. 2016, 21, 179–182. [Google Scholar] [CrossRef]
- Fernández-Pacheco, P.; Ramos Monge, I.M.; Fernández-González, M.; Poveda Colado, J.M.; Arévalo-Villena, M. Safety evaluation of yeasts with probiotic potential. Front. Nutr. 2021, 8, 659328. [Google Scholar] [CrossRef] [PubMed]
- Bojsen, R.; Regenberg, B.; Folkesson, A. Saccharomyces cerevisiae biofilm tolerance towards systemic antifungals depends on growth phase. BMC Microbiol. 2014, 14, 305. [Google Scholar] [CrossRef] [PubMed]
- Canteri de Souza, P.; Custódio Caloni, C.; Wilson, D.; Sergio Almeida, R. An invertebrate host to study fungal infections, mycotoxins and antifungal drugs: Tenebrio molitor. J. Fungi 2018, 4, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antifungals | MIC *, [mg/L] |
|---|---|
| Amphotericin B | 0.25 |
| 5-Flucytosine | <0.125 |
| Caspofungin | 0.5 |
| Fluconazole | 1 |
| Voriconazole | <0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, L.S.; Mokus, L.; Frota, H.F.; Santos, M.V.; Oliveira, S.S.C.; Oliveira, M.M.E.; Costa, G.L.; Alves, A.L.; Bernardes-Engemann, A.R.; Orofino-Costa, R.; et al. SARS-CoV-2 Post-Infection and Sepsis by Saccharomyces cerevisiae: A Fatal Case Report—Focus on Fungal Susceptibility and Potential Virulence Attributes. Trop. Med. Infect. Dis. 2023, 8, 99. https://doi.org/10.3390/tropicalmed8020099
Ramos LS, Mokus L, Frota HF, Santos MV, Oliveira SSC, Oliveira MME, Costa GL, Alves AL, Bernardes-Engemann AR, Orofino-Costa R, et al. SARS-CoV-2 Post-Infection and Sepsis by Saccharomyces cerevisiae: A Fatal Case Report—Focus on Fungal Susceptibility and Potential Virulence Attributes. Tropical Medicine and Infectious Disease. 2023; 8(2):99. https://doi.org/10.3390/tropicalmed8020099
Chicago/Turabian StyleRamos, Lívia S., Luca Mokus, Heloisa F. Frota, Marcos V. Santos, Simone S. C. Oliveira, Manoel M. E. Oliveira, Gisela L. Costa, Ana Luísa Alves, Andréa R. Bernardes-Engemann, Rosane Orofino-Costa, and et al. 2023. "SARS-CoV-2 Post-Infection and Sepsis by Saccharomyces cerevisiae: A Fatal Case Report—Focus on Fungal Susceptibility and Potential Virulence Attributes" Tropical Medicine and Infectious Disease 8, no. 2: 99. https://doi.org/10.3390/tropicalmed8020099
APA StyleRamos, L. S., Mokus, L., Frota, H. F., Santos, M. V., Oliveira, S. S. C., Oliveira, M. M. E., Costa, G. L., Alves, A. L., Bernardes-Engemann, A. R., Orofino-Costa, R., Aor, A. C., Branquinha, M. H., & Santos, A. L. S. (2023). SARS-CoV-2 Post-Infection and Sepsis by Saccharomyces cerevisiae: A Fatal Case Report—Focus on Fungal Susceptibility and Potential Virulence Attributes. Tropical Medicine and Infectious Disease, 8(2), 99. https://doi.org/10.3390/tropicalmed8020099